Adelophthalmus
| Adelophthalmus | |
|---|---|
| |
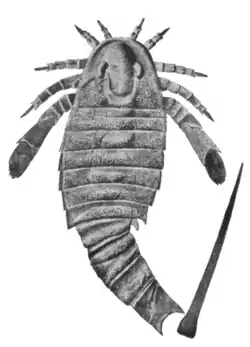
| Fóssil de A. mansfieldi ilustrado por James Hall | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Arthropoda |
| Subfilo: | Chelicerata |
| Ordem: | †Eurypterida |
| Família: | †Adelophthalmidae |
| Gênero: | †Adelophthalmus Jordan in Jordan & von Mayer, 1854[1] |
| Espécie-tipo | |
| †Adelophthalmus granosus Jordan in Jordan & von Meyer, 1854[1]
| |
| Espécies | |
|
33 espécies
| |
| Sinónimos | |
|
Sinônimos do gênero
Sinônimos de A. mansfieldi
Sinônimos de A. moyseyi
Sinônimos de A. perornatus
Sinônimos de A. sievertsi
Sinônimos de A. sellardsi
| |
Adelophthalmus é um gênero de euriptérido, um grupo extinto de artrópodes aquáticos. Fósseis de Adelophthalmus foram descobertos em depósitos que variam em idade do Devoniano Inferior ao Permiano Inferior, o que o torna o mais longevo de todos os gêneros de euriptéridos conhecidos, com um alcance temporal total de mais de 120 milhões de anos. Adelopththalmus foi o gênero final da subordem Eurypterina de euriptéridos e consistiu no único gênero conhecido de euriptéridos nadadores do Devoniano Médio até sua extinção durante o Permiano, após o qual os poucos euriptéridos sobreviventes eram todos formas caminhantes da subordem Stylonurina.
Fósseis de Adelophthalmus foram descritos em quatro continentes; América do Norte, Europa, Ásia e Austrália, o que indica que Adelophthalmus pode ter tido uma distribuição quase cosmopolita (mundial), um dos poucos gêneros de euriptéridos a alcançar tal feito, além de potencialmente Pterygotus. A expansão territorial de Adelophthalmus começou cedo, com representantes encontrados tanto na Sibéria quanto na Austrália durante o Devoniano, mas alcançou sua distribuição quase cosmopolita após a amálgama do supercontinente Pangeia durante o Carbonífero e o Permiano.
O nome genérico Adelophthalmus significa "sem olho óbvio", referindo-se ao fato de que o fóssil holótipo aparentemente representava um euriptérido sem olhos, com uma carapaça (placa da cabeça) completamente desprovida de qualquer indicação de olhos. Embora isso tenha causado muita confusão subsequente, incluindo a nomeação de vários sinônimos juniores, a aparente ausência de olhos no espécime-tipo é tratada pelos pesquisadores modernos como um artefato de preservação, e não uma característica que qualquer espécie de Adelophthalmus teria possuído em vida.
Adelophthalmus era um gênero de euriptéridos comparativamente pequenos, com espécies variando em tamanho de 4 cm (Adelophthalmus douvillei) a 32 cm (Adelophthalmus khakassicus). Em 2020, Adelophthalmus é o mais diverso taxonomicamente de todos os gêneros de euriptéridos, contendo 33 espécies consideradas válidas. Essa grande quantidade de espécies, muitas nomeadas há muito tempo, levou alguns pesquisadores a designar Adelophthalmus como um "táxon da lixeira", com relações internas e filogenia mal conhecidas. O gênero, como é visto atualmente, pode formar um grupo monofilético (um grupo que compartilha um ancestral comum), mas talvez fosse mais apropriadamente dividido em diferentes gêneros ao longo de clados distintos formados dentro dos limites atuais do gênero.
Descrição

Euriptéridos adelophthalmídeos como Adelophthalmus eram pequenos e aerodinâmicos euriptéridos nectônicos (nadadores ativos) com proeminentes esculturas cuticulares (ornamentação consistindo de pequenas e diminutas escamas em suas costas).[2] Essas escamas são talvez a característica mais distintiva do grupo, embora escamas semelhantes tenham sido relatadas em outros grupos de euriptéridos também, mais notavelmente nos pterigotídeos.[3][4]
Embora o maior adelophthalmídeo, Adelophthalmus era, em comparação com membros predadores de ápice maiores do grupo (como Jaekelopterus), um gênero de euriptéridos relativamente pequenos. A maior espécie de Adelophthalmus conhecida, Adelophthalmus khakassicus, atingiu um comprimento máximo de aproximadamente 32 cm.[5] Muitas espécies eram menores, sendo a menor a Adelophthalmus douvillei do Permiano, com apenas 4 cm de comprimento. O gênero como um todo não parece ter flutuado muito em tamanho ao longo de sua longa história evolutiva, com espécies "grandes" ocorrendo no Devoniano (Adelophthalmus sievertsi com 18 cm e Adelophthalmus waterstoni com 15 cm), no Carbonífero (os já mencionados Adelophthalmus mazonensis, Adelophthalmus wilsoni com 20 cm, e tanto Adelophthalmus granosus quanto Adelophthalmus zadrai com 15 cm) e durante o Permiano (Adelophthalmus luceroensis com 18 cm). A maioria das espécies menores é conhecida do Carbonífero, quando Adelophthalmus era mais abundante, incluindo os de "tamanho médio" Adelophthalmus irinae (13 cm) e Adelophthalmus moyseyi (12 cm) e os menores Adelophthalmus mansfieldi, Adelophthalmus pennsylvanicus (ambos com 8 cm), Adelophthalmus approximatus (7 cm) e Adelophthalmus dumonti (6 cm).[6]

Como a maioria dos euriptéridos (com algumas exceções, como Slimonia e Rhinocarcinosoma), a carapaça (o segmento que cobre o prossoma, a "cabeça") de Adelophthalmus tinha formato parabólico, com uma borda marginal estreita. A carapaça era mantida no lugar com a ajuda de um pequeno mecanismo de "travamento" triangular articulado, posicionado anteriormente. Os olhos eram reniformes (em forma de feijão) e os pequenos ocelos estavam localizados entre, ou ligeiramente atrás (dependendo da espécie), dos olhos maiores.[7] O metastoma [en] (uma grande placa que faz parte do abdômen) de Adelophthalmus tinha formato oval, com o primeiro opistossomal (o opistossoma refere-se a todos os segmentos após a carapaça, essencialmente o abdômen) tendo um comprimento reduzido e sendo afilado lateralmente. O corpo de Adelophthalmus terminava com um telson (o segmento mais posterior, aqui na forma de um espigão) longo e afiado, de formato estiliforme.[8] A característica que distingue principalmente Adelophthalmus de outros euriptéridos adelophthalmídeos é seu corpo alongado e os esporões presentes em seus segmentos abdominais.[9]
Tabela de espécies
O status dos 35 nomes (dos quais dois são sinônimos) listados abaixo segue um levantamento de 2018 dos paleontólogos alemães Jason A. Dunlop e Denise Jekel e do paleontólogo britânico David Penney, e os alcances de tamanho e temporais seguem um estudo de 2009 dos paleontólogos americanos James Lamsdell e Simon J. Braddy, salvo indicação em contrário.[10][6]
| Espécies | Autor | Ano | Status | Comprimento | Intervalo temporal | Notas & descrição |
|---|---|---|---|---|---|---|
| Adelophthalmus approximatus | Hall & Clarke | 1888 [en] | Válido | 7 cm | Fameniano (Devonian) | Originalmente descrito como uma espécie de Eurypterus.[7] A. approximatus é semelhante a A. mansfieldi, sendo os espinhos ao longo do abdômen muito semelhantes. A. approximatus pode ser diferenciado de A. mansfieldi pela falta de diferenciação entre os três primeiros pares e o último par de endognátitos ser menos distinto..[11] |
| Adelophthalmus asturica | Melendez | 1971 [en] | Válido | ? cm | Moscoviano (Carbonferous) | Originalmente descrita como uma espécie do gênero Lepidoderma, que foi sinônimo..[7] As nadadeiras posteriores da A. asturica eram particularmente grandes..[12] |
| Adelophthalmus bradorensis | Bell | 1922 [en] | Válido
Possível sinônimo de A. imhofi[13] |
? cm | Casimoviano (Carboniferous) | Originalmente descrita como uma espécie de Anthraconectes. A. bradorensis é muito semelhante à A. kidstoni, diferindo apenas por ser proporcionalmente mais curta e não possuir recortes nas bordas do exoesqueleto, como a A. kidstoni..[14] |
| Adelophthalmus caledonicus | Peach | 1882 [en] | Sinônimo de A. perornatus | – | – | Originalmente descrita como uma espécie do gênero sinônimo Glyptoscorpius.[15] Designado como sinônimo de A. perornatus, embora tenha sido considerado significativamente diferente por possuir “órgãos em forma de pente” pelo paleontólogo britânico Charles D. Waterston em 1958..[16] |
| Adelophthalmus cambieri | Pruvost | 1930 [en] | Válido
Possível sinônimo de A. imhofi[13] |
? cm | Basquiriano (Carboniferous) | Originalmente descrita como uma espécie do gênero sinônimo Anthraconectes.[7] |
| Adelophthalmus carbonarius | Chernyshev | 1933 [en] | Incerto | ? cm | Basquiriano – Casimoviano (Carboniferous) | Originalmente descrito como uma espécie de Eurypterus.[17] A. carbonarius diferia de A. luceroensis nas proporções do corpo. Sua carapaça tinha uma relação comprimento/largura semelhante ao valor médio dos espécimes de A. luceroensis da mesma classe de tamanho. A mesma relação entre o comprimento do metasoma e o comprimento do mesossoma era um pouco maior do que em A. luceroensis, enquanto a relação entre o comprimento do prosoma e o comprimento do mesossoma era menor do que neste último..[18] |
| Adelophthalmus chinensis | Grabau | 1920 [en] | Válido | ? cm | Latest Carboniferous – Asseliano (Permian) | Originalmente descrita como uma espécie do gênero sinônimo Anthraconectes.[7] |
| Adelophthalmus corneti | Pruvost | 1939 [en] | Válido
Possível sinônimo de A. imhofi[13] |
? cm | Basquiriano (Carboniferous) | Originalmente descrita como uma espécie do gênero sinônimo Anthraconectes.[7] |
| Adelophthalmus douvillei | de Lima | 1890 [en] | Válido | 4 cm | Asseliano – Sacmariano (Permian) | Originalmente descrito como uma espécie de Eurypterus.[7] |
| Adelophthalmus dubius[19] | Shpinev | 2012 [en] | Válido
Possível sinônimo de A. kamyshtensis[5] |
18 cm[19] | Middle Devonian[19] | Uma espécie ligeiramente maior do que a média, A. dubius é incrivelmente pouco conhecida, sendo que o único espécime conhecido não possui olhos, apêndices e até mesmo as espinhas abdominais, que normalmente estão sempre presentes em Adelophthalmus. A espécie é mais semelhante a A. mazonensis (diferenciando-se no tamanho e tendo um prosoma relativamente mais longo), A. moyseyi (diferenciando-se no tamanho e tendo um prosoma mais estreito), A. nebraskensis (diferenciando-se no tamanho e tendo um prosoma mais largo), A. wilsoni (diferenciando-se por um prosoma menor e mais estreito) e A. zadrai (diferenciando-se no tamanho)..[19] A. dubius, A. khakassicus e A. kamyshtensis podem ser sinônimos..[5] |
| Adelophthalmus dumonti | Stainier | 1915 [en] | Válido | 6 cm | Moscoviano (Carboniferous) | Originalmente descrito como uma espécie de Eurypterus.[20] Muito semelhante ao A. mansfieldi, semelhante na forma geral e proporções e no padrão da ornamentação. Eles diferem em A. dumonti, que tem uma carapaça mais larga, olhos maiores, um tórax mais delgado e os espinhos característicos ao longo do abdômen apontando para trás (em A. mansfieldi, eles apontam para trás e para fora)..[21] |
| Adelophthalmus granosus | Jordan | 1854 [en] | Válido, espécie-tipo | 15 cm | Moscoviano (Carboniferous) | A. granosus tinha proporções relativamente amplas. Com o único espécime conhecido sem olhos e apêndices, seu status como diagnóstico é um tanto questionável. É possível que os grandes espinhos abdominais de A. granosus estejam nos esternitos [en] (uma característica compartilhada apenas por A. nebraskensis) e não nos tergitos [en], como em outras espécies, mas essa característica também pode ser simplesmente devido a deformação..[22] |
| Adelophthalmus imhofi | Reuss | 1855 [en] | Válido | ? cm | Moscoviano (Carboniferous) | Originalmente descrito como a espécie-tipo do gênero Lepidoderma, que foi sinônimo..[23] |
| Adelophthalmus irinae | Shpinev | |2006 [en] | Válido | 13 cm | Turnaciano (Carboniferous) | |
| Adelophthalmus khakassicus | Shpinev & Filimonov | 2018 [en] | Válido
Possível sinônimo de A. kamyshtensis[5] |
32 cm[5] | Givetiano (Devonian) | A. khakassicus é semelhante a A. mazonensis, A. moysei e A. sellardsi. Difere de todas estas espécies por ter um mesossoma mais estreito e um metasoma mais largo. Possuía epímeras subdesenvolvidas no sétimo, décimo primeiro e décimo segundo segmentos (sendo estes dois últimos segmentos achatados e semelhantes a folhas). A. khakassicus, A. kamyshtensis e A. dubius podem ser sinônimos..[5] |
| Adelophthalmus kamyshtensis[24] | Shpinev | 2012 [en] | Válido | 15 cm[24] | Middle Devonian[24] | Espécie de tamanho médio e pouco conhecida, a A. kamyshtensis pode ser distinguida da maioria das outras espécies pelo seu primeiro segmento ser mais estreito em relação aos outros segmentos do mesossoma. Os espinhos abdominais característicos estavam presentes no último segmento do mesossoma e em todos os segmentos metasomáticos. Em comparação com outras espécies, a A. kamyshtensis é mais semelhante à A. luceroensis, A. sellardsi, A. imhofi, A. granosus, A. mazonensis, A. wilsoni e A. sievertsi..[24] A. kamyshtensis, A. khakassicus e A. dubius podem ser sinônimos..[5] |
| Adelophthalmus kidstoni | Peach | 1888 [en] | Válido
Possível sinônimo de A. imhofi[13] |
? cm | Moscoviano (Carboniferous) | Originalmente descrita como uma espécie do gênero sinônimo Glyptoscorpius.[25] A. kidstoni é semelhante à A. bradorensis, mas possui recortes nas bordas do seu exoesqueleto, uma característica que a diferencia de todas as outras espécies conhecidas de Adelophthalmus..[14] A. kidstoni era consideravelmente diferente da A. wilsoni, uma espécie contemporânea da mesma localização..[26] |
| Adelophthalmus lohesti | Dewalque | 1889 [en] | Incerto
Possíveis afinidades com estilônurídeos [en] |
? cm | Fameniano (Devonian) | Originalmente descrito como uma espécie de Eurypterus. A. lohesti é questionavelmente referido como Adelophthalmus e tem várias características que não são consistentes com o gênero. Estas incluem uma carapaça muito larga, olhos muito grandes e o que parece ser uma crista mediana na sua carapaça..[27] |
| Adelophthalmus luceroensis | Kues & Kietzke | 1981 [en] | Válido | 18 cm | Asseliano (Permian) | Espécie de tamanho médio intimamente relacionada com outras espécies de Adelophthalmus encontradas na América do Norte, a A. luceroensis distingue-se pelo seu prosoma invulgarmente largo (em outras espécies, como A. imhofi e A. mansfieldi, o prosoma é praticamente tão longo quanto largo, mas em A. luceroensis é consideravelmente mais largo do que longo)..[28] |
| Adelophthalmus mansfieldi | Hall | 1877 [en] | Válido | 8 cm | Latest Carboniferous – Early Permian | Originalmente descrito como uma espécie de Dolichopterus [en].[29] A. mansfieldi é talvez a mais semelhante à A. dumonti, mas tem uma carapaça mais estreita, olhos menores e um tórax mais largo. Os espinhos ao longo do abdômen de A. mansfieldi apontam para trás e para fora..[21] |
| Adelophthalmus mazonensis | Meek & Worthen | 1868 [en] | Válido | 22 cm | Moscoviano (Carboniferous) | Originalmente descrita como a espécie-tipo do gênero sinônimo Anthraconestes..[30] |
| Adelophthalmus moyseyi | Woodward | 1907 [en] | Válido | 12 cm | Moscoviano (Carboniferous) | Originalmente descrito como uma espécie de Eurypterus e depois transferido para o gênero sinônimo Anthraconestes..[31] Esta espécie é semelhante à A. mansfieldi, mas os espinhos ao longo do abdômen são visivelmente menos proeminentes em A. moyseyi..[32] |
| Adelophthalmus nebraskensis | Barbour | 1914 [en] | Válido | 6 cm[33] | Sacmariano (Permian) | Originalmente descrita como uma espécie do gênero sinônimo Anthraconectes.[7] Os espinhos ao longo do abdômen da A. nebraskensis estão localizados nos esternitos, o que diferencia essa espécie de todas as outras espécies conhecidas de Adelophthalmus (com a possível exceção de A. granosus)..[22] A. nebraskensis também é conhecida por seu telson excepcionalmente longo e seu corpo geralmente esguio e delgado..[33] |
| Adelophthalmus oklahomensis | Decker | 1938 [en] | Sinônimo de A. sellardsi | – | – | A. oklahomensis foi sinonimizada com a idêntica A. sellardsi (de idade e formação estratigráfica semelhantes no Kansas) em 1959..[27] |
| Adelophthalmus pennsylvanicus | Hall | 1877 [en] | Válido | 8 cm | Moscoviano (Carboniferous) | Originalmente descrito como uma espécie de Eurypterus.[7] |
| Adelophthalmus perornatus | Peach | 1882 [en] | Incerto
Possíveis afinidades com hibbertopterídeos [en] |
? cm | Viseiano (Carboniferous)[7] | Originalmente descrito como a espécie-tipo do gênero sinônimo Glyptoscorpius..[25] Os espécimes fósseis fragmentados (consistindo em apenas cinco tergitos[16]) referidos a A. perornatus são invulgarmente grandes para um Adelophthalmus e apresentam uma ornamentação mais semelhante à observada na família Hibbertopteridae [en] do que em Adelophthalmus..[34] |
| Adelophthalmus piussii | Lamsdell, Simonetto & Selden | 2013 [en] | Válido | 4 cm[35] | Late Carboniferous | A. piussii é única dentro do gênero Adelophthalmus por possuir um sulco mediano (estrutura elevada no centro da carapaça) no prosoma e pelos cantos da carapaça não serem expandidos. Seu primeiro tergito tem uma morfologia quase idêntica à de A. wilsoni..[35] |
| Adelophthalmus pruvosti | Kjellesvig-Waering | 1948 [en] | Válido
Possível sinônimo de A. imhofi[13] |
? cm | Moscoviano (Carboniferous) | Originalmente descrita como uma espécie do gênero Lepidoderma, que foi sinônimo..[7] |
| Adelophthalmus pyrrhae | Lamsdell et al. | 2020 [en] | Válido | 7 cm[36] | Turnaciano (Carboniferous)[37] | A. pyrrhae tinha, em seus segundo a quinto apêndices, um par de espinhos ventrodistais em cada podômero. Além disso, tinha epímeras em seu pós-abdômen e o primeiro segmento não apresentava nenhuma redução lateral. Isso sugere que A. pyrrhae era semelhante a A. mansfieldi e A. mazonensis..[36] |
| Adelophthalmus raniceps | Goldenberg | 1873 [en] | Incerto | ? cm | Moscoviano (Carboniferous) | Originalmente descrito como uma espécie de Polyzosternites.[7] |
| Adelophthalmus sellardsi | Dunbar | 1924 [en] | Válido
Possível sinônimo de A. imhofi[13] |
? cm | Artinsquiano (Permian) | Originalmente descrita como uma espécie do gênero sinônimo Anthraconectes. A. sellardsi parece se assemelhar a A. mansfieldi, diferindo em uma carapaça ligeiramente mais longa e arredondada em A. mansfieldi. Outras espécies relacionadas, como A. chinensis, têm uma carapaça mais curta e arredondada do que A. sellardsi, bem como epímeras proeminentes no pré-abdômen, ao contrário desta última..[38] |
| Adelophthalmus sievertsi | Størmer | 1969 [en] | Válido | 18 cm | Emsiano (Devonian) | Originalmente descrita como uma espécie de Rhenopterus [en].[7] A. sievertsi, apesar de sua idade precoce, é mais semelhante às espécies do final do Carbonífero e do Permiano, como A. sellardsi e A. luceroensis. A espécie pode ser diferenciada das outras do gênero por sua carapaça relativamente larga, um podômero 7 curto nas pernas natatórias e fileiras de tubérculos ao longo das bordas posteriores da carapaça e dos segmentos opistosomais..[39] |
| Adelophthalmus waterstoni | Tetlie et al. | 2004 [en] | Válido | 15 cm | Frasniano (Devonian) | Originalmente descrita como uma espécie de Rhenopterus.[7] Uma espécie pouco conhecida, baseada em um único fóssil composto por uma série de segmentos fragmentados..[40] Semelhante a A. sievertsi, mas diferindo na disposição dos tubérculos nos segmentos..[41] |
| Adelophthalmus wilsoni | Woodward | 1888 [en] | Válido
Possível sinônimo de A. imhofi[13] |
20 cm | Moscoviano (Carboniferous) | Originalmente descrito como uma espécie de Eurypterus.[7] O único espécime conhecido é composto por seis segmentos corporais. Esses segmentos possuem marcas e espinhos ao longo do abdômen semelhantes aos de A. mansfieldi. Os espinhos de A. wilsoni são menos pontiagudos do que os de A. mansfieldi..[42] |
| Adelophthalmus zadrai | Přibyl | 1952 [en] | Válido
Possível sinônimo de A. imhofi[13] |
15 cm | Basquiriano (Carboniferous) | Os espinhos abdominais do A. zadrai são mais angulares em comparação com outras espécies, como A. granosus (onde são mais arredondados). Outras diferenças entre A. zadrai e a espécie-tipo são o fato da ornamentação de A. zadrai ser mais grosseira, e A. zadrai ser significativamente mais esguio na forma..[43] |
História de pesquisa
Primeiras descobertas
O primeiro espécime de Adelophthalmus a ser descoberto foi escavado em 1851 pelo paleontólogo alemão Hermann Jordan em um poço de ferrovia em Jägersfreude, perto de Saarbrücken, na Alemanha. Este espécime foi descrito três anos depois, em 1854, na obra Ueber die Crustaceen der Steinkohlenformation von Saarbrücken ("Sobre os crustáceos da formação de carvão de Saarbrücken"), escrita por Jordan e Hermann von Meyer e apresentando descrições de vários outros táxons de artrópodes. O fóssil foi imediatamente reconhecido por Jordan como o de um euriptérido, com tanto a forma geral quanto as partes individuais (particularmente a cabeça e os apêndices) sendo muito semelhantes às de Eurypterus, que havia sido descrito nos Estados Unidos em 1825, 29 anos antes. Entre as diferenças notadas entre os espécimes estavam o tamanho menor e a idade posterior do fóssil de Saarbrücken e o que Jordan e von Meyer perceberam ser uma completa ausência de olhos.[1]
Como a carapaça preservada não tinha indicação de que alguma vez houvesse olhos presentes, Jordan e von Meyer presumiram que o animal teria sido completamente sem olhos em vida, com a descrição original do fóssil citando vários casos em que formas sem olhos ocorrem em grupos de artrópodes que, de outra forma, possuem olhos (como em crustáceos e trilobitas).[1] Essa aparente ausência de olhos motivou a escolha do nome, Adelophthalmus, que significa "sem olho óbvio".[31] O nome da espécie, granosus, deriva do latim grānōsus ("granulado" ou "cheio de grãos"), referindo-se ao estado de preservação do fóssil que deu a alguns dos fósseis uma textura granulada.[1][44] O espécime-tipo, até hoje o único espécime referido Adelophthalmus granosus, está guardado nas coleções de paleontologia de artrópodes do Museu de História Natural de Berlim sob o número de espécime MB.A. 890.[45]

Embora os pesquisadores modernos tendam a tratar a suposta ausência de olhos como um artefato de preservação e não uma característica que A. granosus teria tido em vida, essa questão não foi resolvida imediatamente, o que tornou a nomeação de espécies descobertas posteriormente confusa e problemática.[46] Lepidoderma imhofi, nomeado em 1855 a partir de depósitos da idade do Carbonífero na Alemanha, mostra olhos definidos. O descritor, o paleontólogo austríaco August Emanuel von Reuss [en], notou que Lepidoderma provavelmente era sinônimo de Adelophthalmus, mas ignorou as regras de prioridade taxonômica e usou seu nome mais recente por ser baseado em material que ele considerava mais bem preservado.[23] O nome Lepidoderma deriva do latim lepidus ("elegante" ou "fino") e do grego antigo δέρμα (ðerma, "pele").[47][48]
Em 1868, os paleontólogos americanos Fielding Bradford Meek [en] e Amos Henry Worthen [en] descreveram Anthraconectes mazonensis, sendo Anthraconectes designado um subgênero de Eurypterus, com base em fósseis recuperados em depósitos da idade do Carbonífero em Mazon Creek [en], Illinois (a primeira espécie a ser descrita da América do Norte).[30] Após examinar o espécime-tipo de Adelophthalmus em 1934, o paleontólogo alemão Paul Guthörl observou que Anthraconectes e Adelophthalmus eram tão semelhantes que teriam sido sinônimos se Adelophthalmus possuísse olhos.[31] O nome Polyzosternites foi cunhado pelo paleontólogo alemão Friedrich Goldenberg (que também nomeou a espécie Polyzosternites raniceps, hoje reconhecida como A. raniceps) em 1873 para substituir o nome Adelophthalmus em relação a espécimes descritos após o espécime-tipo, na crença de que o tipo de Adelophthalmus representava os restos fósseis de uma barata.[49] Glyptoscorpius foi erigido para incluir alguns fósseis do Carbonífero da Escócia, incluindo as espécies Glyptoscorpius perornatus (designada como tipo, o espécime-tipo consistindo de apenas cinco tergitos[16]), Glyptoscorpius caledonicus e Glyptoscorpius kidstoni, pelo geólogo britânico Ben Peach em 1882.[25] Glyptoscorpius seria por muito tempo erroneamente considerado como representando os restos fósseis de um escorpião e não de um euriptérido.[50]
A segunda espécie a ser descrita da América do Norte foi Adelophthalmus pennsylvanicus (como Eurypterus pennsylvanicus), por Meek e Worthen a partir das camadas de carvão do condado de Venango, Pensilvânia, em 1877. No mesmo ano, o paleontólogo americano James Hall descreveu a espécie Adelophthalmus mansfieldi (sob o nome Eurypterus (Dolichopterus) mansfieldi) com base em fósseis recuperados em Cannelton [en], também na Pensilvânia.[29] Em 1888, Hall descreveu a espécie Adelophthalmus approximatus (como Eurypterus approximatus) juntamente com o paleontólogo americano John Mason Clarke [en] com base em fósseis também recuperados da Pensilvânia.[10]
O geólogo inglês Henry Woodward descreveu a espécie Eurypterus wilsoni (=Adelophthalmus wilsoni) em 1888 com base em um fóssil recuperado por um Edward Wilson do Museu de Bristol [en], nomeando a espécie em sua homenagem. O único espécime conhecido é composto por seis segmentos corporais e Woodward notou que nomear a espécie pode ter sido um pouco prematuro. Ele observou que o espécime possuía marcações e espinhos ao longo do abdômen de maneira semelhante a Adelophthalmus mansfieldi (então classificado como Eurypterus mansfieldi).[42]
O paleontólogo português Pereira de Lima descreveu a espécie Eurypterus douvillei (hoje vista como Adelophthalmus douvillei) em 1890 com base em fósseis de Bussaco, em Portugal.[10]
Século XX
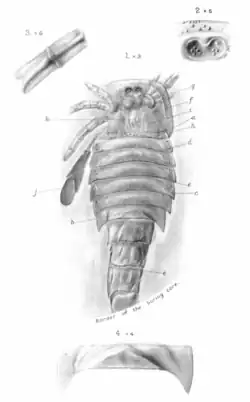
Em 1907, Henry Woodward descreveu Eurypterus moyseyi (hoje reconhecido como Adelophthalmus moyseyi) com base em fósseis recuperados de Radstock, Somerset [en], na Inglaterra. Woodward comparou o único espécime de E. moyseyi com espécimes fósseis de Adelophthalmus mansfieldi da América, achando os espinhos ao longo do abdômen muito semelhantes, embora tenha notado que eram menos proeminentes em Eurypterus moyseyi. Woodward descreveu espécimes fósseis muito grandes, com a carapaça medindo sozinha 21 cm e os sete segmentos corporais associados medindo juntos mais 25 cm.[32] Apesar disso, as estimativas de tamanho mais recentes disponíveis para Adelophthalmus moyseyi colocam a espécie com 12 cm de comprimento.[6]
Adelophthalmus nebraskensis foi descrito como Eurypterus (Anthraconectes) nebraskensis em 1914 pelo geólogo americano Erwin H. Barbour [en] com base em fósseis recuperados em Nebraska, EUA. A espécie foi descrita juntamente com outros fósseis dos sedimentos associados, o que ajudou a reforçar a ideia de Adelophthalmus (ou Anthraconectes) como um animal de água doce.[33]
A espécie Adelophthalmus dumonti, de idade Carbonífera, foi descrita pelo paleontólogo belga Xavier Stainier em 1915 como Eurypterus dumonti. O espécime-tipo, um fóssil relativamente completo medindo apenas 3,3 centímetros de comprimento, foi descoberto através de perfuração [en] em um novo campo de carvão em Campine [en], norte da Bélgica. Embora o fóssil tivesse sido ligeiramente danificado, incluindo a contraparte inteira sendo fragmentada, devido ao uso descuidado de martelos e brocas de diamante durante a escavação, o fóssil pôde, no entanto, ser estudado em detalhes e comparado com espécies de euriptéridos conhecidas.[20] Como Stainier considerava que todo euriptérido conhecido do Carbonífero fazia parte do gênero Eurypterus (entre eles várias espécies hoje reconhecidas como Adelophthalmus, como a espécie-tipo Adelophthalmus granosus, Adelophthalmus imhofi e Adelophthalmus pennsylvanicus), ele classificou o novo euriptérido belga nesse gênero também.[51] Ele notou que a nova espécie era muito semelhante a espécies como Eurypterus pennsylvanicus e especialmente Eurypterus mansfieldi (ambas vistas como espécies de Adelophthalmus hoje).[21] O nome da espécie dumonti homenageia o proeminente geólogo belga André Dumont.[52]
O geólogo americano Amadeus William Grabau [en] descreveu a espécie Anthraconectes chinensis em 1920, com base em fósseis descobertos em Zhaozezhuang, China.[10]
O geólogo canadense Walter A. Bell descreveu a espécie Adelophthalmus bradorensis em 1922 (como uma espécie de Anthraconectes) com base em um único fóssil recuperado em New Campbelton no Condado de Victoria [en], Canadá, referindo-a ao gênero devido a semelhanças com a escocesa Adelophthalmus kidstoni e a americana Adelophthalmus mansfieldi.[14]
O ano de 1924 viu a descrição da espécie Anthraconectes sellardsi pelo paleontólogo americano Carl Owen Dunbar [en] com base em dois fósseis incompletos e alguns outros pequenos fragmentos de Elmo, no Kansas. O primeiro espécime preserva a carapaça e os quatro primeiros tergitos do pré-abdômen, enquanto o segundo preserva cinco tergitos pré-abdominais e três pós-abdominais; este espécime representa o dobro do tamanho do primeiro.[53]
A espécie Adelophthalmus oklahomensis foi descrita pelo paleontólogo americano Carl E. Decker em 1938 com base em fósseis de idade Permiana em Oklahoma. Como o espécime de A. oklahomensis era virtualmente idêntico a espécimes de Adelophthalmus sellardsi de idade semelhante e de um horizonte estratigráfico semelhante no Kansas, Adelophthalmus oklahomensis foi designado um sinônimo júnior de Adelophthalmus sellardsi pelo geólogo americano Carl Colton Branson, com o apoio de Decker, em 1959.[27]
O espécime-tipo de Adelophthalmus zadrai, MB.A. 889, foi coletado na República Tcheca em 1930 ou 1931 e mencionado pela primeira vez em um manuscrito do trabalhador do Carbonífero francês Pierre Pruvost, que o apelidou de "Eurypterus (Anthraconectes) Zadrai", mas ele não descreveu formalmente o espécime ou táxon. Pruvost tinha experiência anterior com o gênero, tendo descrito a espécie Anthraconectes cambieri em 1930 com base em fósseis de Charleroi, Bélgica. A. zadrai foi descrito formalmente pela primeira vez em 1952 como Adelophthalmus zadrai, num momento em que o espécime-tipo parecia perdido. O espécime foi redescoberto em Berlim sob um nome de espécie diferente com base no coletor original do fóssil (Dr. Palisa) e sem qualquer designação de que representava um espécime-tipo. Pruvost também foi homenageado através da nomeação de A. pruvosti (descrito como Lepidoderma pruvosti pelo paleontólogo norueguês Erik N. Kjellesvig-Waering em 1948 com base em fósseis descobertos em Lens, França).[8][10]
Em 1933, o paleontólogo ucraniano Boris Isidorovich Chernyshev descreveu a espécie Adelophthalmus carbonarius com base em um único espécime do Donets na Ucrânia. Uma nova expedição em 2012 realizada pelo paleontólogo russo Evgeniy S. Shpinev e outros nas localidades respectivamente russa e ucraniana de Kakichev e Lomuvatka trouxe uma série de fósseis bem preservados, presumivelmente juvenis, de A. carbonarius. A identificação exata desses fósseis não é possível, mas eles são identificados como A. carbonarius, pois não há características que mostrem o contrário.[54] Outra espécie belga, Adelophthalmus corneti, foi descrita por Pruvost em 1939 com base em fósseis de Quaregnon.[10]
Todos os gêneros sinônimos; Anthraconectes, Glyptoscorpius, Lepidoderma e Polyzosternites, foram subsumidos em Adelophthalmus em estudos durante meados do século XX, notavelmente o do paleontólogo belga Fredrik Herman van Oyen (1956).[7] Embora a maioria dos autores atribua todas as espécies descritas a Adelophthalmus, alguns, como van Oyen em 1956, consideraram que Anthraconectes poderia representar um gênero distinto, citando que escorpiões com anatomias dorsais semelhantes podem ser bastante diferentes ventralmente e que o mesmo poderia ser verdade para o Adelophthalmus do Carbonífero, onde a morfologia ventral ainda não é conhecida. Um gênero Anthraconectes dessa natureza seria problemático devido à sua classificação depender do estado de preservação de qualquer espécime dado.[46]
Adelophthalmus asturica foi descrito como Lepidoderma asturica pelo paleontólogo espanhol Bermudo Meléndez em 1971 com base em fósseis de d'Ablana, na Espanha.[10]
A espécie Adelophthalmus luceroensis foi descrita pelos paleontólogos americanos Barry S. Kues e Kenneth K. Kietzke em 1981 com base em 150 espécimes fósseis recuperados da formação Madera [en] do Novo México. A grande quantidade de espécimes recuperados, representando indivíduos em vários estágios de desenvolvimento e ontogenia, permitiu que estudos detalhados fossem realizados sobre a ontogenia e a variação intraespecífica dentro de Adelophthalmus.[55]
O paleontólogo americano Roy E. Plotnick referiu uma espécie de Eurypterus, Eurypterus lohesti (descrita pela primeira vez em 1889) a Adelophthalmus em 1983 (como Adelophthalmus lohesti), mas essa classificação é questionável, pois a morfologia do espécime de A. lohesti não é consistente com a conhecida de Adelophthalmus. As diferenças incluem A. lohesti ter olhos maiores, uma carapaça mais larga e o que poderia ser uma crista mediana na carapaça.[27]
Século XXI

Em 2004, o paleontólogo alemão Markus Poschmann referiu a espécie Adelophthalmus sievertsi, descrita pela primeira vez como parte do gênero Rhenopterus pelo paleontólogo norueguês Leif Størmer em 1969 com base em restos fósseis da formação Klerf [en] do Devoniano na Alemanha, ao gênero. Poschmann também referiu a espécie Rhenopterus waterstoni (descrita no início de 2004 com base no único espécime BMNH In 60174 do Devoniano Superior da Austrália) a Adelophthalmus. Esta espécie não havia sido anteriormente atribuída ao gênero, apesar de claras semelhanças com outras espécies de Adelophthalmus, em parte devido à falta anterior de evidências sólidas da presença de Adelophthalmus já no Devoniano.[41]
Adelophthalmus irinae foi descrito em 2006 com base em espécimes fósseis (incluindo o holótipo, um prossoma, "cabeça", com o número de espécime PIN nº 5109/4) coletados pela expedição geológica de Krasnoyarsk perto de Sakhapta, uma vila no distrito de Nazarovsky [en] da região de Krasnoyarsk da Rússia. Os fósseis, da formação Solomennyi Stan do Tournaisiano, puderam ser confiantemente atribuídos a Adelophthalmus com base em sua ornamentação semelhante a escamas, a posição de seus olhos e a forma da carapaça logo após sua escavação. A espécie é a primeira espécie de Adelophthalmus a ser descrita da Rússia e o primeiro euriptérido do Carbonífero conhecido do país. É também um dos poucos euriptéridos do Carbonífero encontrados no território da antiga União Soviética, sendo os únicos outros Adelophthalmus carbonarius da Ucrânia e Unionopterus do Cazaquistão.[56]
Shpinev descreveu duas novas espécies de Adelophthalmus em 2012; Adelophthalmus kamyshtensis e Adelophthalmus dubius (o nome derivando do latim dubius = "duvidoso"), ambas baseadas em fósseis originalmente coletados pelo geólogo russo Yuriy Fedorovich Pogonya-Stefanovich em 1960 em depósitos a 3 km a sudeste da vila de Kamyshta (que emprestou seu nome a Adelophthalmus kamyshtensis) da República da Cacássia, Rússia, e agora alojados no Instituto Paleontológico Borissiak [en]. Apesar de quão mal preservados são esses fósseis, várias características (notavelmente a carapaça parabólica e a presença de espinhos ao longo do abdômen) colocam ambas as espécies dentro de Adelophthalmus.[57]
Em 2013, Adelophthalmus piussii tornou-se o primeiro euriptérido a ser descrito da Itália. O espécime (número de espécime MFSNgp 31681, alojado no Museo Friulano di Storia Naturale ("Museu Friulano de História Natural") em Udine) foi coletado no banco de cascalho de um pequeno riacho perto do riacho maior Bombaso, ao norte da vila de Pontebba e consiste em uma carapaça e sete segmentos opistossomais em um grande bloco de arenito. O nome da espécie, piussii, homenageia o coletor do espécime-tipo, Stefano Piussi.[58]
Em 2018, Shpinev e o pesquisador russo A. N. Filimonov descreveram uma nova espécie chamada Adelophthalmus khakassicus com base em muitos espécimes bem preservados. Encontrado na formação Ilemorovskaya da Cacássia (daí o nome) em 2014 por Filimonov, representa a maior espécie do gênero. O holótipo, PM TGU 168/108, é composto por partes do metassoma e um telson completo, com vários outros parátipos conhecidos. Como Adelophthalmus khakassicus é conhecido de níveis estratigráficos semelhantes aos de Adelophthalmus kamyshtensis e Adelophthalmus dubius, foi sugerido que essas três espécies poderiam representar sinônimos.[59]
Em 2020, Lamsdell, Victoria E. McCoy, Opal A. Perron-Felle e Melanie J. Hopkins descreveram uma nova espécie de Adelophthalmus do estágio Tournaisiano da (muito provavelmente) formação Lydiennes [en], na França. Seu único espécime conhecido, GLAHM A23113, é um corpo quase completo, faltando apenas o telson e preservado em nódulos [en] fosfáticos [en]. Por esta razão, foi chamado de Adelophthalmus pyrrhae, em homenagem a Pirra da Tessália, uma figura da mitologia grega que, juntamente com seu marido Deucalião, atirou pedras que se transformaram em bebês para repovoar o mundo. A boa preservação de Adelophthalmus pyrrhae permitiu aos pesquisadores examinar partes de seu sistema respiratório, e após seu estudo foi confirmado que, mesmo que tivessem um estilo de vida principalmente aquático, os euriptéridos podiam se aventurar em terra por longos períodos.[60]
História evolutiva
Devoniano
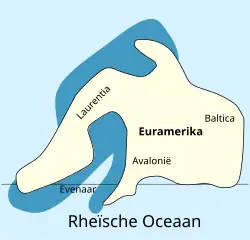
Adelophthalmídeos provavelmente apareceram pela primeira vez nas águas do continente Báltica no Siluriano Superior, fazendo parte de uma rápida diversificação de euriptéridos nadadores (subordem Eurypterina) ao longo do Siluriano. A Báltica colidiria mais tarde com os continentes Avalônia e Laurência e formaria a massa de terra Euramérica, onde a maior parte da evolução basal dos adelophthalmídeos ocorreu no Devoniano Inferior.[61] A espécie mais antiga conhecida de Adelophthalmus é Adelophthalmus sievertsi de depósitos do Devoniano Inferior (Emsiano) da formação Klerf em Wilwerath (na Renânia-Palatinado), Alemanha, então parte da Avalônia dentro da Euramérica. A. sievertsi vivia em ambientes próximos à costa, tipicamente um habitat variado e instável, o que indica que Adelophthalmus era euritópico (capaz de sobreviver em uma ampla gama de ambientes).[39]
Três outras espécies do Devoniano Médio, Adelophthalmus khakassicus, Adelophthalmus kamyshtensis e Adelophthalmus dubius, são as espécies mais antigas conhecidas de Adelophthalmus de fora da Europa, com fósseis das três tendo sido recuperados da Cacássia na Rússia.[57][62] No Devoniano Superior, Adelophthalmus já havia se espalhado, com a espécie Adelophthalmus waterstoni tendo sido recuperada de depósitos de idade Frasniana (~382,2 a 372,2 milhões de anos) na formação Gogo [en] da Austrália Ocidental, o único euriptérido, com exceção de Acutiramus e Pterygotus, conhecido do continente.[63][64]
A única outra espécie de Adelophthalmus conhecida do Devoniano é a Adelophthalmus lohesti do Fameniano (Devoniano mais tardio), conhecida de depósitos fósseis em Pont de Bonne em Liège, Bélgica. Juntamente com uma espécie Fameniana de Hibbertopterus, Hibbertopterus dewalquei, Adelophthalmus lohesti representa o euriptérido mais antigo até então descoberto na Bélgica. A. lohesti é, no entanto, representado por um único espécime fragmentário cuja identificação como Adelophthalmus ou mesmo eurypterino em geral é questionável, podendo representar um euriptérido stylonurídeo [en].[65] Espécimes devonianos de Adelophthalmus foram supostamente recuperados também da Sibéria, o que significaria que a distribuição do gênero incluía águas ao redor de todos os continentes então existentes.[61]
Os euriptéridos foram um dos grupos mais afetados pelo evento de extinção do Devoniano Superior. Após um grande declínio na diversidade durante o Devoniano Inferior, os euriptéridos eram raros em ambientes marinhos no Devoniano Superior. Das 16 famílias de euriptéridos que estavam vivas no início do Devoniano, apenas três persistiram no Carbonífero. Todos esses eram grupos não marinhos.[66] Enquanto a subordem Stylonurina permaneceu relativamente ilesa, adaptando novas estratégias (como a alimentação por varredura) para evitar a competição, e veio a se diversificar novamente no Carbonífero, a Eurypterina foi quase completamente extinta, com Adelophthalmus tornando-se o único sobrevivente de toda a subordem.[67]
Carbonífero

Após a extinção de todos os outros euriptéridos nadadores no Devoniano, Adelophthalmus tornou-se o mais comum de todos os euriptéridos do Paleozoico tardio, existindo em número muito maior do que os membros sobreviventes de Stylonurina, tanto em termos de número de indivíduos quanto de número de espécies.[68] Adelophthalmus experimentou uma rápida diversificação ao longo do Carbonífero, com 23 de suas 33 espécies tendo sido descritas apenas do Carbonífero, e atingiu seu pico de diversidade no Pensilvânico (Carbonífero Superior).[10][69][37] Essa diversificação não levou à evolução de nenhum novo gênero – Adelophthalmus permaneceu o único gênero de euriptéridos eurypterinos até a extinção do grupo.[67]
Já difundido e representado em todas as principais massas de terra no Devoniano Superior, a amálgama da Pangeia em um supercontinente global durante o Carbonífero e o Permiano permitiria ao hábil nadador Adelophthalmus ganhar uma distribuição quase mundial, com fósseis de Adelophthalmus da idade do Carbonífero tendo sido recuperados dos Estados Unidos, Espanha, Bélgica, Ucrânia, China, Alemanha, República Tcheca, Rússia, Inglaterra, País de Gales, Escócia, França e Itália.[10][61]
O Carbonífero Inferior viu o aparecimento de algumas novas espécies, notavelmente Adelophthalmus approximatus, o registro mais antigo de Adelophthalmus na América do Norte (embora esta espécie possa ter ocorrido já no estágio Fameniano, o último estágio do Devoniano). O gênero também se espalhou para a Escócia moderna (Adelophthalmus perornatus recuperado de depósitos fósseis do Carbonífero Inferior em Glencartholm [en]) e Ásia (Adelophthalmus irinae de idade Tournaisiana conhecida de depósitos fósseis perto de Krasnoyarsk, Rússia). O aparecimento de A. irinae é particularmente notável, pois representa o único euriptérido do Carbonífero conhecido até então na Rússia.[56] Adelophthalmus pyrrhae, descoberto na França, também é conhecido desta época.[37] O Carbonífero Superior veria o aparecimento de várias outras espécies em vários lugares do mundo. Durante o estágio Bashkiriano (de 323,2 a 315,2 milhões de anos atrás), duas espécies apareceram na Bélgica, Adelophthalmus cambieri de Charleroi e Adelophthalmus corneti de Quaregnon, e uma terceira espécie, Adelophthalmus zadrai, foi relatada em depósitos de idade Bashkiriana em Moravo-Silesia, na República Tcheca.[6]
A abundância do bivalve Anthracomya sugere fortes evidências de deposição de água doce no habitat de Adelophthalmus bradorensis, uma espécie Radstockiana (Westphaliana [en] Superior) do Canadá.[14]
O estágio Moscoviano (de 315,2 a 307 milhões de anos atrás) viu o aparecimento de várias novas espécies, incluindo as duas espécies alemãs Adelophthalmus raniceps e Adelophthalmus granosus, ambas de Saarbrücken. Outras espécies de idade Moscoviana incluem uma variedade de Adelophthalmus da Europa e da América do Norte; Adelophthalmus asturica de d’Ablana, Espanha, Adelophthalmus kidstoni de Radstock, Inglaterra, Adelophthalmus imhofi de Vlkhys, República Tcheca, Adelophthalmus mazonensis de Illinois, EUA, Adelophthalmus moyseyi de Blaengarw, País de Gales, Adelophthalmus pennsylvanicus da Pensilvânia, EUA, Adelophthalmus pruvosti de Lens, França, Adelophthalmus wilsoni de Radstock, Inglaterra e Adelophthalmus dumonti de Mechelen-sur-Meuse, Bélgica. O final do Carbonífero e o início do Permiano veriam o aparecimento de Adelophthalmus mansfieldi na Pensilvânia, EUA e Adelophthalmus chinensis de Zhaozezhuang, China. Além disso, a espécie Adelophthalmus piussii foi recuperada de depósitos do Carbonífero Superior nos Alpes Cárnicos da Itália, o primeiro e até então único euriptérido conhecido do país.[6][10][58]
Permiano


O registro fóssil do Permiano de Adelophthalmus inclui cinco espécies, todas confinadas ao Permiano Inferior. O primeiro estágio do período, o Asseliano (de 298,9 a 295 milhões de anos atrás), viu o aparecimento de Adelophthalmus luceroensis no Novo México, EUA, Adelophthalmus douvillei em Bussaco, Portugal e a sobrevivência contínua da espécie chinesa do Carbonífero Adelophthalmus chinensis. Adelophthalmus douvillei durou até o estágio subsequente, o Sacmariano (de 295 a 290,1 milhões de anos atrás), que também viu o aparecimento de Adelophthalmus nebraskensis em Nebraska, EUA. Sabe-se que A. nebraskensis vivia em um ambiente de água doce, sendo seu fóssil encontrado em associação com fósseis de plantas terrestres.[33] A espécie mais jovem descrita é Adelophthalmus sellardsi, conhecida do estágio Artinsquiano (290,1 a 283,5 milhões de anos atrás) de Elmo [en], no Kansas, EUA.[6][10]
De todas as espécies conhecidas de Adelophthalmus, os últimos remanescentes do gênero (Adelophthalmus luceroensis e Adelophthalmus sellardsi) eram mais semelhantes à primeira espécie conhecida, Adelophthalmus sievertsi, apesar de estarem separados por um período de mais de cem milhões de anos. As semelhanças provavelmente se devem a um nicho ecológico generalizado, e não especializado. Este conservadorismo morfológico em Adelophthalmus sugere que o gênero tornou-se braditélico, evoluindo a um ritmo mais lento do que a taxa padrão entre os euriptéridos. Tipicamente, organismos braditélicos têm uma ampla distribuição geográfica, algo que foi visto em Adelophthalmus ao longo do final do Devoniano e do Carbonífero.[39]
Como muitas outras espécies de Adelophthalmus, Adelophthalmus luceroensis parece ter vivido em ambientes de água salobra a doce em uma planície deltaica adjacente a uma planície costeira. As condições climáticas favoráveis à disseminação e manutenção de tais ambientes foram ótimas durante o Carbonífero Superior e o Permiano Inferior, com Adelopthalmus sendo difundido e numeroso nessas épocas. Na maioria dos locais onde Adelophthalmus estava presente, parece ter sido ecologicamente semelhante.[70]
Embora habitats desse tipo fossem muitos, difundidos e ecologicamente estáveis por um tempo no início do Permiano, eles se revelariam delicados. Uma mudança climática durante o Permiano alterou os padrões de deposição e vegetação em todo o hemisfério norte, o que afetou drasticamente ambientes anteriormente difundidos, como os característicos pantânos de carvão do Carbonífero, bem como habitats de água salobra e doce. À medida que seu habitat desaparecia, Adelophthalmus diminuía em número. Enquanto alguns euriptéridos stylonurinos (Hastimima [en] e Campylocephalus) que ocupavam nichos fora desses habitats continuaram a sobreviver por um tempo, Adelophthalmus, restrito a um tipo de ambiente que desaparecia rapidamente, extinguiu-se.[70]
Classificação
_(20618910349).jpg)

Adelophthalmus é classificado como parte de (e empresta seu nome a) a família Adelophthalmidae, a única família dentro da superfamília Adelophthalmoidea, ao lado dos gêneros Parahughmilleria, Nanahughmilleria, Bassipterus, Pittsfordipterus e Eysyslopterus.[10] O cladograma abaixo apresenta as posições filogenéticas inferidas da maioria dos gêneros incluídos nas três superfamílias mais derivadas da subordem Eurypterina de euriptéridos (Adelophthalmoidea, Pterygotioidea e os waeringopteroides), conforme inferido por O. Erik Tetlie e Markus Poschmann em 2008, com base nos resultados de uma análise de 2008 especificamente pertencente à Adelophthalmoidea e uma análise anterior de 2004.[34]
Uma relação próxima entre os três grupos é confirmada em parte devido a membros basais de todos os três grupos, Orcanopterus, Eysyslopterus e Herefordopterus, compartilharem formas de carapaça semelhantes. Adelophthalmus sendo o membro mais derivado de sua família é confirmado por seus apêndices natatórios serem os mais finos de todos os gêneros incluídos e por seus olhos estarem mais próximos do centro da carapaça. Nos adelophthalmoides, os olhos parecem se aproximar do centro da carapaça a cada gênero mais derivado, e embora a posição dos olhos possa refletir estilos de vida e ambientes habitados, presume-se que eles também (particularmente neste caso, com uma progressão clara) incluam informações filogeneticamente importantes.[34]
| Diploperculata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Filogenia interna e monofilia


A filogenia interna e as relações dentro de Adelophthalmus são pouco conhecidas, devido à sua longa história e à grande quantidade de espécies atribuídas ao gênero, muitas baseadas em restos fragmentários.[35]
O paleontólogo americano Victor P. Tollerton sugeriu em 1989 que algumas espécies de Adelophthalmus poderiam ser melhor colocadas em um novo gênero na família Slimonidae de euriptéridos, citando sua falta de espinhos, no entanto, observou que o material então disponível de Adelophthalmus tornava difícil avaliar se as pernas eram verdadeiramente não espiníferas. Um novo gênero para espécies não espiníferas poderia ser filogeneticamente apoiado, mas transferir o novo gênero para os Slimonidae com base na perda de uma característica que parece ter sido perdida independentemente nos dois grupos não está de acordo com a prática comum.[71]
O cladograma abaixo exibe os resultados de uma análise filogenética conduzida por O. Erik Tetlie e Markus Poschmann em 2008, apresentando sete espécies de Adelophthalmus e excluindo outras espécies com base no fato de que eram muito incompletamente conhecidas. Todos os caracteres foram tratados como não ordenados e receberam peso igual. Orcanopterus, parte de um clado que também contém Grossopterus [en] e Waeringopterus [en], foi incluído na análise como um grupo externo para polarizar os caracteres.[34]
Os resultados da análise mostraram que todos os gêneros apresentados (incluindo Adelophthalmus), com exceção de Nanahughmilleria, onde a espécie basal Nanahughmilleria patteni foi atribuída ao novo gênero Eysyslopterus, eram (ou tinham o potencial de ser) monofiléticos. A monofilia de Adelophthalmus foi apoiada por várias sinapomorfias, incluindo a presença de um triângulo anterior na carapaça (cuja função é incerta), uma área circular central da carapaça sendo elevada, os olhos estando mais distantes da margem da carapaça do que dos ocelos, um metastoma oval, um telson longo e a presença de epímeros no pré-abdômen.[34]
Adelophthalmus sievertsi foi recuperado como mais basal do que outras espécies, o que se encaixa com o fato de ser também a espécie mais antiga conhecida no registro fóssil, principalmente devido ao apêndice natatório largo ser semelhante aos apêndices largos de Parahughmilleria e Nanahughmilleria. Todas as outras espécies de Adelophthalmus onde este apêndice é conhecido possuem um que é mais fino.[34]
| Diploperculata |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
A análise deixou de fora muitas espécies fragmentárias de Adelophthalmus, pois seus estados de caracteres não puderam ser confiantemente levados em conta, e Adelophthalmus em termos de todas as espécies que é reconhecido como contendo não pode, portanto, ser totalmente declarado com confiança como monofilético. Espécies mais fragmentárias precisam ser redescritas e mais caracteres filogenéticos precisam ser confiantemente estabelecidos antes que o status do gênero possa ser certo.[34]
Adelophthalmus, como é atualmente entendido, pode formar um grupo monofilético e, portanto, filogeneticamente válido, mas que provavelmente sofre de uma sub-divisão no nível de gênero e super-divisão no nível de espécie. É possível que a grande quantidade de espécies forme dois ou mais clados distintos que poderiam ser divididos em diferentes gêneros.[72] Embora a maioria das espécies incluídas no gênero pareça formar um grupo monofilético, algumas espécies foram sugeridas como representantes de espécies de outros gêneros reconhecidos, com Adelophthalmus dumonti supostamente sendo semelhante ao obscuro Unionopterus em sua suposta carapaça trapezoide (uma característica agora conhecida por ser incorreta e baseada em uma ilustração incorreta) e o grande Adelophthalmus perornatus mostrando ornamentação semelhante à vista nos Hibbertopteridae.[34]
Muitas das espécies mais fragmentárias poderiam muito bem ser sinônimos de espécies mais conhecidas. Em particular, Adelophthalmus imhofi foi sugerido por Fredrik Herman van Oyen em 1956 como possivelmente representando um sinônimo sênior de muitas espécies, incluindo Adelophthalmus zadrai, Adelophthalmus corneti, Adelophthalmus cambieri, Adelophthalmus pruvosti, Adelophthalmus bradorensis, Adelophthalmus kidstoni, Adelophthalmus wilsoni e Adelophthalmus sellardsi. As sinonimizações de Van Oyen basearam-se apenas em proporções da carapaça, ignorando outras características filogenéticas importantes, bem como possíveis efeitos tafonômicos (defeitos produzidos durante a fossilização) nos fósseis.[27] Pesquisas subsequentes provaram a validade de algumas espécies, agora definidas com base em características claras e distintas, incluindo Adelophthalmus mazonensis, Adelophthalmus mansfieldi e Adelophthalmus moyseyi.[26]
A taxonomia e o status precisos das espécies dentro de Adelophthalmus é uma área de pesquisa contínua. Talvez a questão mais importante que permanece sem resposta seja a relação exata entre a espécie-tipo Adelophthalmus granosus e a segunda espécie mais antiga descrita, Adelophthalmus imhofi, o que poderia ter grandes implicações para a filogenia interna do gênero.[27]
Status como um táxon da lixeira
Adelophthalmus contém uma grande quantidade de espécies (33[10][57][73][37] em 2020, a maior quantidade de qualquer euriptérido), é geograficamente difundido, nomeado há muito tempo (1854) e a forma nominal de um táxon superior (emprestando seu nome à família Adelopthalmidae e à superfamília Adelopthalmoidea), atendendo a todos os critérios para ser apelidado de "táxon da lixeira", um táxon que existe com o único propósito de classificar organismos que não se encaixam em outro lugar.[35]
Além disso, a maioria das espécies referidas a Adelophthalmus foi descrita por autores que não eram especialistas em euriptéridos (já que os pesquisadores de euriptéridos concentraram seus esforços principalmente em euriptéridos mais diversos do pré-Carbonífero) e a maioria das descrições carece de comparações com espécies previamente descritas do gênero. Como tal, as diferenças entre as espécies são muitas vezes triviais, talvez em parte resultando do fato de que o primeiro artigo de revisão sobre o táxon foi publicado apenas em 1948, altura em que 26 espécies já haviam sido descritas.[27]
Paleoecologia
_(20579579132).jpg)
Adelophthalmoidea viviam principalmente em ambientes próximos a habitats costeiros, com preferência por habitats com salinidade reduzida, como deltas de rios, estuários ou lagoas. Influências marinhas são frequentemente registradas nesses habitats e nos depósitos que contêm fósseis de adelophthalmoides, mas fósseis-índice marinhos típicos (fósseis que indicam um ambiente e ecossistema marinho) não são encontrados associados aos restos de euriptéridos. Os fósseis ocasionais de Adelophthalmus encontrados em depósitos obviamente marinhos, como o Adelophthalmus waterstoni do Devoniano Superior da Austrália, podem ter sido transportados de seu habitat original.[74] No caso de A. waterstoni, isso é visto como particularmente provável, pois é representado por um único espécime que também é o único espécime de euriptérido coletado da formação em que foi encontrado, a formação Gogo da Austrália Ocidental.[63]
Em geral, euriptéridos pós-Devonianos são raros e ocorrem em habitats de água salobra ou doce, tendo migrado dos ambientes marinhos marginais habitados durante o Siluriano.[2] Os primeiros adelophthalmoides, como o Parahughmilleria hefteri do Devoniano, são recuperados em depósitos não marinhos, como em ambientes que já foram habitats salobros ou estuarinos. A evolução de Adelophthalmus viu uma mudança de ambientes salobros para habitats dominados por água doce. Em habitats onde tanto Parahughmilleria quanto espécies primitivas de Adelophthalmus são encontrados, como em sítios fósseis do Devoniano Inferior na Alemanha, onde fósseis de Adelophthalmus sievertsi foram descobertos, Parahughmilleria é encontrado em seções que são consideravelmente mais marginalmente marinhas do que as seções habitadas por Adelophthalmus.[74]
A maior presença de Adelophthalmus em habitats de água doce ocorreu nos estágios Bashkiriano e Moscoviano do Carbonífero, dos quais fósseis de Adelophthalmus são recuperados em estratos que contêm carvão (indicando um ambiente de pântano de carvão) juntamente com fósseis de bivalves de água doce e organismos terrestres. É possível que essas "conquistas" de água doce estejam relacionadas à diversificação do próprio gênero e ao aparecimento de várias novas espécies durante o Carbonífero, em vez de refletir uma mudança na preferência de habitat do gênero como um todo. De fato, esses Adelophthalmus de pântanos de carvão parecem formar uma minoria, com a maioria das espécies confinada a bacias parálicas ou de planície em ambientes de deposição com conexões próximas a habitats marginalmente marinhos.[74]
Por exemplo, os últimos exemplos sobreviventes de Adelophthalmus na bacia do Saar-Nahe [en] da Alemanha (de idade Moscoviana) são de uma época em que a bacia fazia parte de, ou pelo menos estava conectada a, uma área de subsidência ocidental e a drenagem da bacia era para o Oceano Paleo-Tethys [en], localizado 1.500 km ao sul. Com o soerguimento geológico no sul durante o Pensilvaniano e o Permiano Inferior, a drenagem passou a ser direcionada para o Oceano Panthalassa ao norte, o que resultou na bacia sendo localizada 1.300 km mais longe do oceano. Nesses depósitos mais jovens, Adelophthalmus não é encontrado em lugar nenhum, o que indica que uma mudança para um ambiente mais distante do oceano causou a extinção dessas populações, o que indica que várias espécies precisavam de alguma forma de conexão com habitats de natureza marginalmente marinha, mesmo que não vivessem neles.[74]
Localidades fósseis posteriores contendo Adelophthalmus, do Moscoviano Superior, do Carbonífero posterior e do Permiano Inferior, mostram uma maior presença em habitats com influência marinha, particularmente habitats de ambientes estuarinos com influência de marés. Apesar de Adelophthalmus se espalhar para ambientes totalmente de água doce, suas conquistas desses ambientes aparentemente não foram tão bem-sucedidas quanto as de outros grupos semelhantes, por exemplo, alguns xifossúrios do Carbonífero da família Belinuridae [en], que ocorreram em lagos e bacias de água doce que careciam completamente de euriptéridos.[74]
Dieta e predação
Como Adelophthalmus em muitos aspectos representava o último de sua espécie, sendo o último euriptérido a possuir apêndices natatórios, ele não existia em faunas diversificadas de euriptéridos como as observadas com gêneros durante o Siluriano ou o Devoniano inferior. Em vez disso, os ambientes de água salobra a doce tipicamente habitados por Adelophthalmus, como a formação Madera do Permiano Inferior no Novo México (onde fósseis de Adelophthalmus luceroensis foram recuperados) preservam outros organismos, como insetos, branquiópodes, ostracodes, milípedes e vermes spirorbídeos [en]. As nadadeiras finas e longas de Adelophthalmus indicam que era um bom nadador, embora seja provável que passasse a maior parte do tempo rastejando na lama. Como as quelíceras (apêndices frontais) de Adelophthalmus eram pequenas, é mais provável que se alimentasse de pequenos organismos, possivelmente em parte dos ostracodes e branquiópodes conhecidos de fósseis associados. Há uma notável falta de insetos nos leitos fósseis com densos fósseis de plantas, onde deveriam ser mais comuns, e uma surpreendente abundância em leitos fósseis com poucos euriptéridos, possivelmente indicando que Adelophthalmus se alimentava de insetos que caíam na água, impedindo que estes fossem preservados como fósseis.[70]
As localidades em que Adelophthalmus foi preservado na formação Madera fazem todas parte do membro Red Tanks, que não preserva nenhum organismo conhecido que teria sido capaz de predar Adelophthalmus. É provável, no entanto, que vários peixes predadores, anfíbios e répteis primitivos conhecidos por estarem presentes na época teriam predado os pequenos euriptéridos. Tanto peixes quanto anfíbios são conhecidos de ambientes semelhantes da mesma idade nas vizinhas montanhas Manzanita.[70]
Respiração
Através de imagens de microtomografia de raios-X [en], os pesquisadores puderam observar em detalhes a estrutura dos órgãos respiratórios do único espécime conhecido de Adelophthalmus pyrrhae. Um nódulo de fosfato no lado ventral do animal está dividido de maneira que a câmara branquial (trato branquial) é visível. Isso revela quatro pares de brânquias foliáceas (brânquias externas dispostas como as páginas de um livro), embora provavelmente fossem cinco, como nos xifossúrios. Estas estão orientadas horizontalmente e todas, exceto as do sexto segmento, são fragmentárias. Lá, elas têm formato oval, presas perto da linha média do corpo e consistem em seis lamelas. Pensa-se que o número de lamelas nos outros segmentos anteriores tenha sido maior, como indicado por alguns fragmentos e um espécime de Onychopterella augusti que tinha 45 lamelas em cada um de seus quatro pares de brânquias foliáceas do segundo ao quinto segmento. As brânquias foliáceas de Onychopterella, no entanto, eram orientadas verticalmente. Isso e um fóssil do xifossúrio Tachypleus syriacus sugerem que as brânquias foliáceas de Adelophthalmus pyrrhae sofreram uma deformação tafonômica e que eram originalmente orientadas verticalmente também.[36]
A superfície dorsal de cada lamela é coberta por trabéculas em forma de pilar regularmente espaçadas, localizadas entre cada lamela, deixando um espaço preenchido com hemolinfa (um fluido encontrado em artrópodes, análogo ao sangue dos vertebrados) em cada uma. As trabéculas são comumente encontradas em aracnídeos com pulmões e representam uma adaptação terrestre para respirar ar. Elas impedem que as lamelas grudem umas nas outras e eliminem o espaço entre elas, o que sufocaria o organismo. Portanto, a presença de trabéculas em A. pyrrhae indica que os euriptéridos eram capazes de respirar em ambientes terrestres com seus órgãos respiratórios, ao contrário dos xifossúrios ou outros euartrópodes basais. Suas trabéculas também são muito semelhantes às dos aracnídeos, especialmente as de um espécime de uma espécie indeterminada de Palaeocharinus [en] do Devoniano.[75]
A presença de trabéculas também confirma que as estruturas vasculares ventrais dos euriptéridos atuavam de fato como estruturas respiratórias ativas durante a respiração aérea, como sugerido anteriormente. Isso e a evidência de incursões terrestres feitas por stylonurinos implicam que os euriptéridos poderiam ficar fora da água por períodos prolongados. Isso não muda o fato de que eram criaturas predominantemente aquáticas, assim como suas nadadeiras (que Adelophthalmus pyrrhae também possuía) indicam. Além disso, é possível que estar fora da água teria sido ineficaz para eles durante sua alimentação, limitando o tempo que permaneciam lá. No entanto, eles podem ter se movido de poça em poça para se reproduzir em locais mais seguros, apoiado pela usual separação entre euriptéridos adultos e juvenis no registro fóssil e pela posse de espermatóforos, que poderiam ter permitido aos euriptéridos armazenar esperma por meses para lhes dar tempo de procurar um ambiente seguro.[76]
Segregação baseada na idade
.jpg)
Na formação Madera, Adelophthalmus e organismos associados viviam em corpos de água salobra a doce no que se presume ter sido uma planície deltaica. A falta de grandes leitos de carvão sugere que as localidades fósseis que renderam Adelophthalmus eram uma região moderadamente elevada com vegetação menos densa e melhor drenagem do que os pântanos que ocupavam grande parte do resto dos Estados Unidos. A descoberta de uma grande assembleia de Adelophthalmus luceroensis, incluindo vários adultos e juvenis, permitiu aos pesquisadores determinar diferentes preferências de habitat para diferentes grupos etários. Indivíduos maiores (adultos) são encontrados associados a grandes fragmentos de plantas, incluindo ramos de Walchia [en] e folhas de Cordaites, mas indivíduos menores (juvenis) são encontrados em leitos fósseis contendo menos material orgânico e principalmente fragmentos de plantas menores. Os grandes fragmentos de plantas do habitat adulto foram depositados em condições calmas, provavelmente através da queda de folhas em lagoas fechadas ou lagoas paradas.[70]
Os juvenis parecem ter se desenvolvido e vivido em condições um tanto diferentes das dos adultos. Em leitos onde os juvenis são mais comuns, fósseis de insetos também são mais comuns, indicando uma falta de adultos que seriam capazes de devorá-los, e a presença de fósseis de plantas menores sugere uma cobertura vegetal menos prolífica, sendo o ambiente juvenil possivelmente áreas mais baixas na planície deltaica entre as lagoas. Periodicamente, tempestades levariam água marinha para as lagoas, onde a salinidade seria, portanto, variável, enquanto os juvenis poderiam viver em ambientes mais frescos e menos variáveis, mais longe da linha da costa. É possível que os adultos acasalassem nos riachos que alimentavam as lagoas e depois retornassem para viver nas lagoas devido a um suprimento de comida mais rico estar presente.[70]
A segregação baseada na idade desse tipo entre juvenis e adultos da mesma população é relativamente normal em artrópodes, por exemplo, juvenis do aparentado e moderno Limulus vivem em diferentes ambientes e regiões que os adultos. A vantagem dessa forma de segregação não é apenas permitir que indivíduos mais jovens vivam em condições mais estáveis do ponto de vista da salinidade, mas também manter os juvenis seguros de situações em que quantidades substanciais de água marinha dizimaram as populações nas lagoas, alterando demais as condições de vida. Em tal situação, as populações mais jovens poderiam, após algum tempo, recolonizar os antigos habitats.[70]
Ver também
- Campylocephalus—o último euriptérido caminhante (stylonurino) sobrevivente conhecido.
- Pterygotus—outro euriptérido com uma distribuição quase mundial.
- Lista de gêneros de Eurypterida
Referências
Citações
- ↑ a b c d e Jordan & von Meyer 1854, pp. 1–15.
- ↑ a b Tetlie & van Roy 2006, p. 79.
- ↑ Tetlie & Dunlop 2005, p. 3.
- ↑ Størmer 1955, p. 23.
- ↑ a b c d e f g Shpinev & Filimonov 2018, p. 1559.
- ↑ a b c d e f Lamsdell & Braddy 2009, Supplementary information.
- ↑ a b c d e f g h i j k l m n o p Tetlie & Dunlop 2005, p. 5.
- ↑ a b Tetlie & Dunlop 2005, p. 4.
- ↑ Størmer 1955, p. 30.
- ↑ a b c d e f g h i j k l m Dunlop, Penney & Jekel 2018, p. 24–25.
- ↑ Clarke & Ruedemann 1912, p. 222.
- ↑ Romano & Meléndez 1985, p. 322.
- ↑ a b c d e f g h Tetlie & Poschmann 2008, p. 239.
- ↑ a b c d Bell 1922, pp. 164–165.
- ↑ O'Connell 1916, p. 30.
- ↑ a b c Waterston 1958, p. 267.
- ↑ Chernyshev 1948, p. 119.
- ↑ Shpinev 2014, pp. 288–291.
- ↑ a b c d Shpinev 2012, pp. 473–474.
- ↑ a b Stainier 1915, p. 639.
- ↑ a b c Stainier 1915, p. 645.
- ↑ a b Tetlie & Dunlop 2005, pp. 7–8.
- ↑ a b Reuss 1855, pp. 81–83.
- ↑ a b c d Shpinev 2012, pp. 470–473.
- ↑ a b c Peach 1882, pp. 517–522.
- ↑ a b Waterston 1968, pp. 3–4.
- ↑ a b c d e f g Tetlie & Dunlop 2005, p. 10.
- ↑ Kues & Kietzke 1981, p. 722.
- ↑ a b Woodward 1907, p. 278.
- ↑ a b Meek & Worthen 1868, p. 544.
- ↑ a b c Wills 1964, p. 475.
- ↑ a b Woodward 1907, pp. 279–281.
- ↑ a b c d Barbour 1914, pp. 201–203.
- ↑ a b c d e f g h Tetlie & Poschmann 2008, pp. 239–241.
- ↑ a b c d Lamsdell, Simonetto & Selden 2013, p. 149.
- ↑ a b c Lamsdell et al. 2020, p. 2.
- ↑ a b c d Lamsdell et al. 2020, p. 1.
- ↑ Dunbar 1924, pp. 200–201.
- ↑ a b c Poschmann 2006, pp. 80–81.
- ↑ Tetlie et al. 2004, p. 805.
- ↑ a b Poschmann 2006, pp. 67.
- ↑ a b Woodward 1888, p. 419.
- ↑ Tetlie & Dunlop 2005, pp. 10–11.
- ↑ Latin Lexicon – granosus.
- ↑ Tetlie & Dunlop 2005, p. 7.
- ↑ a b Tetlie & Dunlop 2005, p. 6.
- ↑ Latin Lexicon – lepidus.
- ↑ Glosbe – δέρμα.
- ↑ Goldenberg 1873, p. 18.
- ↑ Laurie 1895, pp. 526–527.
- ↑ Stainier 1915, p. 641.
- ↑ Stainier 1915, p. 646.
- ↑ Dunbar 1924, p. 199.
- ↑ Shpinev 2014, p. 287.
- ↑ Kues & Kietzke 1981, p. 709.
- ↑ a b Shpinev 2006, pp. 431–433.
- ↑ a b c Shpinev 2012, p. 470.
- ↑ a b Lamsdell, Simonetto & Selden 2013, p. 148.
- ↑ Shpinev & Filimonov 2018, pp. 1553–1559.
- ↑ Lamsdell et al. 2020, pp. 1–5.
- ↑ a b c Tetlie 2007, p. 570.
- ↑ Shpinev & Filimonov 2018, p. 1553.
- ↑ a b Tetlie et al. 2004, p. 801.
- ↑ Bicknell, Smith & Poschmann 2020, p. 10.
- ↑ Tetlie & van Roy 2006, p. 80.
- ↑ Hallam & Wignall 1997, p. 70.
- ↑ a b Lamsdell & Braddy 2009, p. 265.
- ↑ Tetlie & van Roy 2006, p. 81.
- ↑ Lamsdell, Simonetto & Selden 2013, p. 147.
- ↑ a b c d e f g Kues & Kietzke 1981, pp. 725–728.
- ↑ Tetlie & van Roy 2006, p. 83.
- ↑ Lamsdell, Simonetto & Selden 2013, p. 150.
- ↑ Shpinev & Filimonov 2018, p. 1555.
- ↑ a b c d e Tetlie & Poschmann 2008, pp. 241–243.
- ↑ Lamsdell et al. 2020, pp. 2–3.
- ↑ Lamsdell et al. 2020, pp. 3–4.
Bibliografia
- Barbour, Erwin H. (1914). «Eurypterid Beds of Nebraska with Notice of a New Species, "Eurypterus Nebraskaensis"». Nebraska Geological Survey. 4 (12): 193–203
- Bell, Walter A. (1922). «A New Genus of Characeae and New Merostomata from the Coal Measures of Nova Scotia». Transactions of the Royal Society of Canada. 16: 159–167
- Bicknell, Russell D. C.; Smith, Patrick M.; Poschmann, Markus (2020). «Re-evaluating evidence of Australian eurypterids». Gondwana Research. 86: 164–181. Bibcode:2020GondR..86..164B. doi:10.1016/j.gr.2020.06.002
- Chernyshev, Boris I. (1948). «New representative of Merostomata from the Lower Carboniferous». State University of Kiev, Geological Collections. 2: 119–130
- Clarke, John Mason; Ruedemann, Rudolf (1912). The Eurypterida of New York. [S.l.]: University of California Libraries. ISBN 978-1125460221
- Dunbar, Carl O. (1924). «Kansas Permian insects, Part 1, The geologic occurrence and the environment of the insects». American Journal of Science. 7 (39): 171–209. Bibcode:1924AmJS....7..171D. doi:10.2475/ajs.s5-7.39.171
- Dunlop, Jason A.; Penney, David; Jekel, Denise (2018). «A summary list of fossil spiders and their relatives» (PDF). World Spider Catalog. [S.l.]: Natural History Museum Bern
- Goldenberg, Friedrich (1873). Fauna Saraepontana Fossilis. Die fossilien Thiere aus der Steinkohlenformation von Saarbrücken. [S.l.]: Chr. Möllinger Verlag
- Hallam, Anthony; Wignall, Paul B. (1997). Mass Extinctions and Their Aftermath. [S.l.]: Oxford University Press. ISBN 978-0198549161
- Jordan, Hermann; von Meyer, Hermann (1854). «Ueber die Crustaceen der Steinkohlenformation von Saarbrücken». Palaeontographica. 4: 1–15
- Kues, Barry S.; Kietzke, Kenneth K. (1981). «A Large Assemblage of a New Eurypterid from the Red Tanks Member, Madera Formation (Late Pennsylvanian-Early Permian) of New Mexico». Journal of Paleontology. 55 (4): 709–729. JSTOR 1304420
- Lamsdell, James C.; Braddy, Simon J. (2009). «Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates». Biology Letters. 6 (2): 265–269. PMC 2865068
 . PMID 19828493. doi:10.1098/rsbl.2009.0700
. PMID 19828493. doi:10.1098/rsbl.2009.0700 - Lamsdell, James C.; McCoy, Victoria E.; Perron-Feller, Opal A.; Hopkins, Melanie J. (2020). «Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion». Current Biology. 30 (21): 4316–4321. PMID 32916114. doi:10.1016/j.cub.2020.08.034
- Lamsdell, James C.; Simonetto, Luca; Selden, Paul A. (2013). «First Eurypterid from Italy: A new species of Adelophthalmus (Chelicerata: Eurypterida) from the Upper Carboniferous of the Carnic Alps (Friuli, NE Italy)». Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy). 119 (2): 147–151. doi:10.13130/2039-4942/6029
- Laurie, Malcolm (1895). «The Anatomy and Relations of the Eurypteridæ». Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 37 (2): 509–528. doi:10.1017/S0080456800032713
- Meek, Fielding Bradford; Worthen, Amos Henry (1868). Paleontology of Illinois. Col: Geological Survey of Illinois. 3: Geology and Palaeontology. [S.l.: s.n.]
- O'Connell, Marjorie (1916). «The Habitat of the Eurypterida». The Bulletin of the Buffalo Society of Natural Sciences. 11 (3): 1–278
- Peach, Ben N. (1882). «Further Researches among the Crustacea and Arachnida of the Carboniferous Rocks of the Scottish Border». Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 30 (2): 511–529. ISSN 2053-5945. doi:10.1017/S0080456800026569
- Poschmann, Markus (2006). «The Eurypterid Adelophthalmus Sievertsi (chelicerata: Eurypterida) from the Lower Devonian (emsian) Klerf Formation of Willwerath, Germany». Palaeontology. 49 (1): 67–82. doi:10.1111/j.1475-4983.2005.00528.x
- Romano, Marco; Meléndez, Bermudo (1985). «An arthropod (Merostome) ichnocoenosis from the Carboniferous of northwest Spain.». Ninth International Geological Congress, Urbana, Illinois. 5: 317–325
- Reuss, Adolf E. (1855). «Über eine neue Krusterspecies aus der Böhmischen Steinkohlenformation». Denkschriften der Königlich-kaiserlichen Akademie der Wissenschaften in Wien. 10: 81–83
- Shpinev, Evgeniy S. (2006). «A new species of Adelophthalmus (Eurypterida) from the lower carboniferous of the Krasnoyarsk Region». Paleontological Journal. 40 (4): 431–433. doi:10.1134/S0031030106040083
- Shpinev, Evgeniy S. (2012). «New species of the genus Adelophthalmus (Eurypterida, Chelicerata) found in the Middle Devonian of Khakassia». Paleontological Journal. 46 (5): 470–475. doi:10.1134/S0031030112050103
- Shpinev, Evgeniy S. (2014). «New data on eurypterids (Eurypterida, Chelicerata) of the Upper Carboniferous of the Donets Basin». Paleontological Journal. 48 (3): 287–293. doi:10.1134/S0031030114030162
- Shpinev, Evgeniy S.; Filimonov, A. N. (2018). «A New Record of Adelophthalmus (Eurypterida, Chelicerata) from the Devonian of the South Minusinsk Depression». Paleontological Journal. 52 (13): 1553–1560. doi:10.1134/S0031030118130129
- Stainier, Xavier (1915). «On a New Eurypterid from the Belgian Coal Measures». Quarterly Journal of the Geological Society. 71 (1–4): 639–647. doi:10.1144/GSL.JGS.1915.071.01-04.24
- Størmer, Leif (1955). «Merostomata». Treatise on Invertebrate Paleontology, Part P Arthropoda 2, Chelicerata. [S.l.]: University of Kansas Press. ASIN B0043KRIVC
- Tetlie, O. Erik; Braddy, Simon J.; Butler, Piers D.; Briggs, Derek E. G. (2004). «A New Eurypterid (Chelicerata: Eurypterida) from the Upper Devonian Gogo Formation of Western Australia, With A Review of the Rhenopteridae». Palaeontology. 47 (4): 801–809. doi:10.1111/j.0031-0239.2004.00390.x
- Tetlie, O. Erik; Dunlop, Jason A. (2005). «A redescription of the Late Carboniferous eurypterids Adelophthalmus granosus von Meyer, 1853 and A. zadrai Přibyl, 1952». Fossil Record. 8 (1): 3–12. ISSN 1860-1014. doi:10.1002/mmng.200410001
- Tetlie, O. Erik; van Roy, Peter (2006). «A reappraisal of Eurypterus dumonti Stainier, 1917 and its position within the Adelophthalmidae Tollerton, 1989» (PDF). Bulletin de l'Institut Royal des Sciences Naturelles de Belgique. 76: 79–90
- Tetlie, O. Erik (2007). «Distribution and dispersal history of Eurypterida (Chelicerata)». Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. ISSN 0031-0182. doi:10.1016/j.palaeo.2007.05.011
- Tetlie, O. Erik; Poschmann, Markus (2008). «Phylogeny and palaeoecology of the Adelophthalmoidea (Arthropoda; Chelicerata; Eurypterida)». Journal of Systematic Palaeontology. 6 (2): 237–249. doi:10.1017/S1477201907002416
- Wills, Leonard J. (1964). «The ventral anatomy of the Upper Carboniferous eurypterid Anthraconectes Meek and Worthen» (PDF). Palaeontology. 7 (3): 474–507
- Waterston, Charles D. (1958). «The Scottish Carboniferous Eurypterida». Transactions of the Royal Society of Edinburgh. 63 (2): 265–288. doi:10.1017/S0080456800009492
- Waterston, Charles D. (1968). «Further Observations on the Scottish Carboniferous Eurypterids». Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 68 (1): 1–20. doi:10.1017/s0080456800014472
- Woodward, Henry (1888). «Note on Eurypterus from the Carboniferous». Geological Magazine. 5 (9): 419–421. Bibcode:1888GeoM....5..419W. doi:10.1017/S0016756800182494
- Woodward, Henry (1907). «Two New Species of Eurypterus from the Coal-Measures of Ilkeston, Derbyshire». Geological Magazine. 4 (6): 277–282. Bibcode:1907GeoM....4..277W. doi:10.1017/S0016756800133515
Websites
- «Glosbe – δέρμα». glosbe.com (em inglês)
- «Latin Lexicon – granosus». latinlexicon.org (em inglês)
- «Latin Lexicon – lepidus». latinlexicon.org (em inglês)
| ||||||||||||||||||||||||||||||||
| Taxonomia |
| | ||||||||||||||||||||||||||||||
| Geocronologia |
| |||||||||||||||||||||||||||||||
| Geografia |
| |||||||||||||||||||||||||||||||
| Gêneros notáveis | ||||||||||||||||||||||||||||||||
| Ichnotáxon |
| |||||||||||||||||||||||||||||||
| Grupos relacionados | ||||||||||||||||||||||||||||||||
| Artigos relacionados |
| |||||||||||||||||||||||||||||||