Erettopterus
Erettopterus
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Rudaniano–Lochkoviano, 443,7–412,3 Ma | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Espécie-tipo | |||||||||||||||||
| †Erettopterus bilobus Salter, 1856 | |||||||||||||||||
| Espécies | |||||||||||||||||
19 espécies
| |||||||||||||||||
| Sinónimos | |||||||||||||||||
| |||||||||||||||||

Erettopterus[1] é um gênero de euriptérido predador de grande porte, um grupo extinto de artrópodes aquáticos. Fósseis de Erettopterus foram encontrados em depósitos que variam do Siluriano Inferior (Rudaniano) ao Devoniano Inferior (Lochkoviano), sendo atribuídos a várias espécies. Fósseis foram recuperados em dois continentes: Europa e América do Norte. O nome do gênero é composto pelas palavras do grego antigo ἐρέττω (eréttō), que significa "remador", e πτερόν (pterón), que significa "asa", traduzindo-se como "asa de remador".
Erettopterus é classificado como parte da família de euriptéridos Pterygotidae, distinguido de outros euriptéridos por seus télsons achatados (o segmento mais posterior do corpo) e suas quelíceras modificadas (apêndices frontais), terminando em quelas bem desenvolvidas (garras). Embora alguns euriptéridos da família Pterygotidae, como Jaekelopterus ou Acutiramus, atingissem proporções gigantescas, estima-se que a maior espécie de Erettopterus, E. Osiliensis, alcançasse 90 cm de comprimento. Erettopterus possuía um télson bilobado (dividido em dois lobos), que é sua característica principal. As formas das quelíceras variam amplamente entre as espécies, mas geralmente são longas, com dentes pequenos e curvados sem serrilhas.
Estudos sobre as quelíceras e olhos compostos de Erettopterus revelaram que era um predador com alta acuidade visual, mas não tão especializado ou ativo quanto Jaekelopterus e Pterygotus. Era mais semelhante a Slimonia acuminata, provavelmente usando suas quelíceras alargadas para agarrar em vez de uma alimentação mais especializada.
Descrição
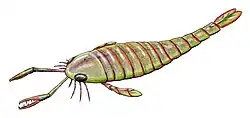
Erettopterus era um euriptérido grande, com E. osiliensis, a maior espécie, medindo aproximadamente 90 cm de comprimento.[1] Embora grande em relação à maioria dos artrópodes modernos, Erettopterus era pequeno em comparação com muitos membros de sua família ( Pterygotidae), como Jaekelopterus rhenaniae, com 2,5 m (o maior artrópode conhecido) e Acutiramus bohemicus, com 2,1 m.[1]
E. grandis atingiu tamanhos ainda maiores, 2,5 m (8 pés), mas esse tamanho é indicado por um télson incompleto isolado (o segmento mais posterior do corpo), portanto, não é totalmente confiável. A menor espécie foi E. globiceps, com apenas 9 cm (3,5 pol.),[1] embora os fósseis de E. globiceps possam ser restos de espécimes filhotes, o que significaria que a espécie poderia ter atingido tamanhos maiores.[2]
O télson era expandido e bilobado (dividido em dois lobos), sua característica principal, que o diferencia dos outros gêneros da família Pterygotidae. Historicamente, pensava-se que Erettopterus tinha cinco articulações em suas quelíceras, mas estudos com espécimes completos de Erettopterus e Acutiramus revelaram que o número real é de quatro articulações.[3] A forma das quelíceras em Erettopterus era muito variável,[4] mas geralmente eram muito longas, com dentes pequenos e curvados sem serrilhas. Seu metastoma (uma grande placa que faz parte do abdômen) era muito estreito e cordado (em forma de coração) anteriormente, com uma entalhe profundo em forma de V.[5] As pernas natatórias eram curtas. O corpo tinha forma oval alongada, gradualmente afunilando-se no abdômen.[6] Os olhos compostos eram amplamente crescentes e convexos,[6] e, como nos outros membros de sua família, estavam localizados na margem da carapaça.[7]
História da pesquisa
Século XIX
_(7394023710).jpg)
Um total de 19 espécies válidas foram atribuídas a Erettopterus. A maioria das espécies foi encontrada nos Estados Unidos ou na Grã-Bretanha, embora fósseis também tenham sido encontrados no Canadá, na Escandinávia e na Estônia.[8] A espécie-tipo e a única espécie descrita na Escócia, E. bilobus, foi encontrada pela primeira vez em Lesmahagow [en] e descrita por John William Salter [en] em 1856 como uma espécie de Himantopterus [en] (um nome que é um homônimo júnior de um gênero de mariposa). Salter substituiu o nome do gênero por Erettopterus em 1859,[9] embora mais tarde tenha sido referida como uma espécie de Pterygotus.[6] É uma espécie muito conhecida e abundante que, desde sua descrição original, não recebeu muita atenção, necessitando de uma redescrição para acomodar o entendimento moderno da sistemática e evolução dos euriptéridos.[10] O nome específico bilobus refere-se ao télson bilobado da espécie.[11]
O geólogo inglês Salter reconheceu em 1859 que era possível dividir o gênero Pterygotus com base na morfologia dos télsons das espécies atribuídas a ele. Ele dividiu Pterygotus em subgêneros, incluindo Pterygotus (Erettopterus) para espécies com télson bilobado.[12] O nome genérico é composto pelas palavras do grego antigo ἐρέττω[13] (eréttō, remador)[11] e πτερόν (pterón, asa),[11] traduzido como "asa de remador". Na mesma época, a primeira espécie inglesa de Erettopterus, E. gigas, foi descrita por Salter e Thomas Henry Huxley. É conhecida por múltiplos espécimes mal preservados descobertos em depósitos da idade Pridoli (Siluriano Superior). O epíteto específico gigas enfatiza o grande tamanho indicado pelos fósseis[13] (embora o tamanho estimado atual seja 25 cm).[1] Quando outra espécie da Inglaterra, do Ludlow Superior (Siluriano Superior), foi descrita em 1961 por Erik N. Kjellesvig-Waering como E. megalodon, foram descobertas várias semelhanças entre as espécies, incluindo a posse de um dente central no ramo fixo (a ramificação externa do apêndice). Em ambas as espécies, o dente central é serrilhado ao longo da borda interna e seguido por dentes de tamanhos irregulares. No entanto, os de E. megalodon são mais semelhantes a espinhos e mais longos que os de E. gigas, o que justifica o nome específico. Todas essas características indicam uma relação próxima entre ambas as espécies.[5]
Em seguida, duas espécies na América do Norte foram descritas. Uma delas foi a canadense E. canadensis (referindo-se ao país onde foi descoberta)[11] descrita por John William Dawson [en] da idade Wenlock Superior (Siluriano Superior), conhecida por um ectognato (maxilípede, um apêndice usado na alimentação) bem preservado encontrado em uma placa de calcário de Niagara. Este ectognato de 8,8 mm de comprimento tem um processo maxilar estreito com aproximadamente 15 dentículos (serrilhas dentárias), incluindo um, o dentículo posterior, que é largo e ligeiramente entalhado na frente.[14] A outra foi E. grandis (grandis pelo grande tamanho da espécie)[11] da idade Pridoli dos Estados Unidos, descrita por Julius Pohlman. Esta espécie foi originalmente descrita como a carapaça (o segmento do exoesqueleto que cobre a cabeça) de um Ceratiocaris [en] gigantesco (um gênero extinto de filocarídeo do Siluriano), mas foi demonstrado que o espécime representa a seção bilobada do télson (o segmento mais posterior do corpo) de um Erettopterus.[2] Este télson incompleto indica que o animal atingiu um tamanho de 2,5 m, mas isso não é totalmente confiável.[1]
Em 1883, mais uma espécie foi descrita por Friedrich Schmidt da idade Ludlow na Estônia, E. osiliensis. Foi descrita a partir de uma grande série de fósseis encontrados em Saaremaa, Estônia. Nesta espécie, a carapaça era semioval com grandes olhos ovais na margem, enquanto os ocelos (olhos simples sensíveis à luz) estavam ligeiramente atrás da metade do comprimento da carapaça. O metastoma era largo, em forma de coração, estreito na parte posterior e truncado no final.[15] Os ramos terminam em uma ponta triangular afiada e bem desenvolvida.[5] E. laticauda foi inicialmente descrito como uma variedade de E. osiliensis pelo mesmo autor, mas devido a diferenças no télson e no metastoma, foi elevado à categoria de espécie. O télson de E. laticauda é muito arredondado, largo e sem serrilhas na parte posterior, ao contrário de E. osiliensis.[3] Além disso, o metastoma era mais oval que em E. osiliensis.[15]
Séculos XX–XXI
No século XX, várias espécies foram descritas na Europa e nos Estados Unidos, ampliando a distribuição de Erettopterus. Em 1912, E. globiceps (globiceps sendo latim para "cabeça esférica")[11] da idade Llandovery (Siluriano Inferior) nos Estados Unidos foi descrito por John Mason Clarke [en] e Rudolf Ruedemann [en]. É até agora a menor espécie de Erettopterus, com apenas 9 cm de comprimento. Os fósseis de E. globiceps são raros e foram encontrados na fauna de Otisville na areia de Shawangunk. Os olhos compostos são muito grandes, ocupando metade do comprimento da carapaça. Isso sugere que E. globiceps poderia atingir tamanhos maiores e que os espécimes encontrados representam filhotes em estágio de crescimento, o que outros espécimes também indicam.[2] As espécies norueguesas E. vogti (homenageando Thorolf Vogt [en], líder das expedições de 1925 a 1928 em Spitsbergen)[16] do Lochkoviano e E. holmi da idade Wenlock Superior foram descritas em 1934 por Leif Størmer [en].[8]

Em 1961, Kjellesvig-Waering elevou Erettopterus ao nível de gênero próprio, reconhecendo dois subgêneros de Pterygotus; P. (Pterygotus) e P. (Acutiramus), bem como dois subgêneros de Erettopterus; E. (Erettopterus) e E. (Truncatiramus).[3] Além disso, ele descreveu quatro novas espécies inglesas, E. marstoni, E. spatulatus, E. megalodon e E. brodiei. E. marstoni (homenageando Alfred Marston, responsável por várias coleções de peixes e euriptéridos) é baseado em espécimes incompletos que, juntos, representam uma quelícera quase completa. O holótipo (BMNH 43790, no Museu de História Natural de Londres) consiste em um ramo livre, e o parátipo (BMNH 43805, no mesmo museu que o holótipo) inclui a maior parte do ramo fixo, que é muito esguio e afunilado na extremidade distal curvada. Um dente grande está presente na seção média do ramo. A extremidade do ramo está quebrada, mas provavelmente terminava em um dente duplo. Essa característica também está presente em E. brodiei, conhecido apenas por um espécime (FMNH 89411, localizado no Museu Field de História Natural) que consiste em uma quela não esmagada. Seu nome específico homenageia Peter Bellinger Brodie [en], cujas coleções de euriptéridos ajudaram a comunidade científica. E. spatulatus é notável por seu télson em forma de pá ou leque, mais largo que longo, que dá o nome específico. Esses télsons são raros e foram vistos apenas em E. grandis.[5] No mesmo ano, ele descreveu uma espécie americana, E. serratus (latim para "serrilhado"),[11] da idade Devoniano Inferior em Ohio. Esta espécie é baseada em um ramo livre completo e bem preservado (FMNH 5104, no Museu Field de História Natural). Esta espécie difere das outras por sua quela espessa e os dentes curtos e robustos.[17] Três anos depois, Kjellesvig-Waering descreveu E. saetiger do Siluriano na Pensilvânia. O holótipo (FMNH 157, abrigado no Museu Field de História Natural) consiste em um metastoma excepcionalmente bem preservado. O metastoma foi definido como aproximadamente elipsoidal, truncado posteriormente e não excessivamente cordado na margem anterior.[3]

Em 1966, outra espécie foi incluída no gênero por Kjellesvig-Waering e Willard P. Leutze com base em uma quelícera, que é o holótipo, dois cefalotórax (cabeça) e um metastoma. Esta espécie foi descrita como E. exophthalmus, da idade Ludlow-Pridoli na Virgínia Ocidental. O cefalotórax é muito longo, com olhos compostos anterolaterais proeminentes, protuberantes e elípticos. A quelícera é composta por um ramo livre bem preservado que retém um soquete de dente duplo. Ambos os dentes terminais são largos e curtos, embora um seja mais longo que o outro. Uma fina estriação é visível em ambos os dentes, mas não nos outros. Esses outros dentes são geralmente pequenos, curvados e de tamanhos irregulares. O metastoma é cordado anteriormente e estreita-se para um posterior arredondado.[18]
Em 1971, E. serricaudatus e E. carinatus da idade Wenlock Inferior na Suécia foram descritos por Kjellesvig-Waering. Foram os únicos membros da família Pterygotidae na área. O holótipo de E. serricaudatus consiste no ramo fixo com terminação aguda de uma grande quelícera. Esta espécie é notável pelo grupo de dentes opostos diagonalmente no ramo. Esta espécie difere de E. osiliensis (espécie na qual E. serricaudatus foi erroneamente incluído) por seus ramos curvados para fora, ao contrário dos ramos retos de E. serricaudatus. Além disso, a forma e o agrupamento dos dentes são completamente diferentes entre eles. O holótipo de E. carinatus é o fragmento da base de um télson que mede 20 mm de comprimento e 13 mm de largura, indicando que o télson completo media 40 mm de comprimento estimado e 20 mm de largura máxima. Esta espécie difere das outras por ter lobos laterais estreitos, estriações grossas na área cordada e seu télson em forma de quilha.[11][19]
Em 1974, Størmer elevou Acutiramus e Truncatiramus ao nível de gêneros separados. As diferenças entre Erettopterus e Truncatiramus estavam nas quelíceras, que eram mais longas em Erettopterus do que em Truncatiramus. Em Erettopterus, os dentes eram curvados e pequenos, enquanto em Truncatiramus eram de tamanho irregular e podiam ser curvados ou retos.[5] No entanto, foi questionado se as quelíceras servem como fator para distinguir entre gêneros, já que sua morfologia depende do estilo de vida e varia ao longo da ontogenia (formação e desenvolvimento individual de um organismo), embora possam ser aceitáveis para diferenciar entre espécies.[4] Portanto, Truncatiramus foi posteriormente reconhecido como um sinônimo de Erettopterus.[12][8]
Além disso, Pterygotus waylandsmithi foi transferido para Erettopterus em 2007 com base nas semelhanças das quelíceras com E. osiliensis. Pterygotus monroensis foi considerado sinônimo de E. osiliensis, estendendo a distribuição da espécie aos Estados Unidos.[12]
Classificação

Erettopterus é classificado como parte da família Pterygotidae,[8] um grupo de euriptéridos altamente derivados (com inovações evolutivas) dos períodos Siluriano ao Devoniano, que diferem de outros grupos por várias características, principalmente nas quelíceras e no télson. As quelíceras dos membros de Pterygotidae eram alargadas e robustas, claramente usadas para caçar.[20] Suas pernas ambulantes eram pequenas e esguias, sem espinhos,[21] e provavelmente não eram capazes de caminhar em terra.[22]
O que distingue Erettopterus dos outros membros de sua família é a morfologia do seu télson bilobado, diferente do télson de Ciurcopterus, com uma quilha mediana dorsal,[7] e do de Pterygotus, Jaekelopterus e Acutiramus, que formam uma espinha curta.[23]
O cladograma abaixo, publicado por Braddy et al. (2007), é baseado nas nove espécies de Pterygotidae mais conhecidas e dois táxons de grupo externo (usados como grupo de referência), Slimonia acuminata e Hughmilleria socialis. O cladograma também contém os tamanhos máximos alcançados pelas espécies em questão, sugeridos como possivelmente uma característica evolutiva do grupo conforme a regra de Cope ("gigantismo filético").[22][24]
| Pterygotioidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecologia

As penas de quelíceras dos membros da família Pterygotidae foram claramente adaptadas para captura ativa de presas, mais semelhantes às garras de alguns crustáceos modernos, com dentes bem desenvolvidos nas garras, do que às quelíceras de outros grupos de euriptéridos.[20] Outra característica que distingue o grupo de outros grupos de euriptéridos eram seus télsons achatados e expandidos, provavelmente usados como lemes ao nadar.[23]
A morfologia das quelíceras e a acuidade visual dos euriptéridos desta família os separam em grupos ecológicos distintos. O método principal para determinar a acuidade visual em artrópodes é contar o número de lentes em seus olhos compostos e o ângulo interommatidial (abreviado em inglês como IOA, referindo-se ao ângulo entre os eixos ópticos das lentes adjacentes). O IOA é particularmente importante, pois pode distinguir diferentes papéis ecológicos em artrópodes, sendo baixo em predadores modernos ativos de artrópodes.[25]
A visão de Erettopterus era semelhante à do gênero mais basal Slimonia e mais aguda do que a do mais derivado Acutiramus, embora não fosse tão aguda quanto a visão dos predadores de topo Jaekelopterus e Pterygotus ou de artigos predadores modernos. Além disso, as grandes quelas de Erettopterus sugerem que era um alimentador generalista, não um predador altamente especializado, e que usava suas quelíceras (apêndices frontais) para agarrar. Os dentes nas quelas de Erettopterus eram na maioria curtos e adequados para segurar a presa, podendo cortá-la eficazmente.[26][25]
O número de lentes nos olhos compostos de Erettopterus é comparável ao número de Pterygotus e de Jaekelopterus, o que sugere que, embora indubitavelmente predatório, Erettopterus não era tão ativo nem tão especializado quanto esses gêneros.[25]
Erettopterus foi encontrado na América do Norte e na Europa. Os depósitos de idade Llandovery das formações Formação Kip Burn [en] e Formação Patrick Burn [en] em Lesmahagow, onde os primeiros fósseis de E. bilobus foram encontrados, preservam fósseis de uma grande quantidade de outros euriptéridos, incluindo Nanahughmilleria lanceolata, Hardieopterus lanarkensis [en], Eusarcana obesus, Parastylonurus sigmoidalis [en], Carcinosoma scorpionis e Slimonia acuminata. Outros organismos encontrados na zona incluem o gastrópode Euomphalopterus, a lampreia Jamoytius [en] ou Loganellia (Thelodonti).[27][28] Os depósitos da Formação Rootsikula em Saaremaa, nos quais foram encontrados fósseis de E. laticauda e E. osiliensis, abrigam várias faunas de euriptéridos, como Mixopterus simonsi, Strobilopterus laticeps e Eysyslopterus patteni. Também foram encontrados restos fósseis de osteostráceos indeterminados e membros de Thelodonti.[29] Outros fósseis também foram encontrados em diferentes lugares, como Grã-Bretanha, Canadá, Escandinávia e Estados Unidos.[8]
Ver também
Referências
- ↑ a b c d e f Lamsdell, James C.; Braddy, Simon J. (2009). «Cope's rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates». Biology Letters. 6 (2): 265–269. ISSN 1744-9561. PMC 2865068
 . PMID 19828493. doi:10.1098/rsbl.2009.0700. Supplemental material
. PMID 19828493. doi:10.1098/rsbl.2009.0700. Supplemental material
- ↑ a b c «The Eurypterida of New York/Volume 1/Pterygotidae - Wikisource, the free online library». en.wikisource.org (em inglês). Consultado em 10 de junho de 2025
- ↑ a b c d Kjellesvig-Waering, Erik N. (1964). «A Synopsis of the Family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida)». Journal of Paleontology. 38 (2): 331–361. JSTOR 1301554
- ↑ a b «An isolated pterygotid ramus (Chelicerata: Eurypterida) from the Devonian Beartooth Butte Formation, Wyoming». Journal of Paleontology. 84 (6): 1206–1208. 2010. doi:10.1666/10-040.1
- ↑ a b c d e Kjellesvig-Waering, Erik N. (1961). «The Silurian Eurypterida of the Welsh Borderland». Journal of Paleontology. 35 (4): 789–835. JSTOR 1301214
- ↑ a b c «The Quarterly Journal of the Geological Society of London». Geological Society of London. 1856
- ↑ a b Tetlie, O. Erik; Briggs, Derek E. G. (2009). «The origin of pterygotid eurypterids». Palaeontology. 52 (5): 1141–1148. Bibcode:2009Palgy..52.1141T. ISSN 1475-4983. doi:10.1111/j.1475-4983.2009.00907.x
- ↑ a b c d e Dunlop, J. A., Penney, D. & Jekel, D. 2015. A summary list of fossil spiders and their relatives. World Spider Catalog. Natural History Museum Bern, consultado em http://wsc.nmbe.ch, versão 18.5 http://www.wsc.nmbe.ch/resources/fossils/Fossils18.5.pdf (PDF).
- ↑ D. Waterston, Charles (1964). «II.—Observations on Pterygotid Eurypterids». Transactions of the Royal Society of Edinburgh. 66 (2): 9–33. doi:10.1017/S0080456800023309
- ↑ Lomax, Dean; Lamsdell, James; Ciurca, Samuel (2011). A collection of eurypterids from the Silurian of Lesmahagow collected pre 1900. 9. [S.l.: s.n.]
- ↑ a b c d e f g h Meaning of bilobus, canadensis, carinatus, eréttō (em grego), globiceps, grandis and pterus. www.wiktionary.org.
- ↑ a b c Ciurca, Samuel J.; Tetlie, O. Erik (2007). «Pterygotids (Chelicerata; Eurypterida) from the Silurian Vernon Formation of New York». Journal of Paleontology. 81 (4): 725–736. ISSN 0022-3360. doi:10.1666/pleo0022-3360(2007)081[0725:PEFTSV]2.0.CO;2
- ↑ a b Huxley, Thomas H.; Salter, John W. (1859). «On the anatomy and affinities of the genus Pterygotus and description of new species of Pterygotus». Printed for H. M. Stationery Off. Memoirs of the Geological Survey of the United Kingdom; Monograph 1. Monograph I (1859): 1–105
- ↑ Harrington, Bernard J.; Donald, James T.; Billings, Elkanah (1881). «The Canadian Naturalist and Quarterly Journal of Science with the Proceedings of the Natural History Society of Montreal». 9: 103–104
- ↑ a b Schmidt, Friedrich (1883) "Die Crustaceenfauna der_Eurypterenschichten_von_Rootziküll_auf_Oesel"
- ↑ Størmer, Leif (1934). I Kommisjon hos Jacob Dybwad, ed. Merostomata from the Downtonian Sandstone of Ringerike, Norway. [S.l.: s.n.]
- ↑ Kjellesvig-Waering, Erik N. (1961) "Eurypterids of the Devonian Holland Quarry Shale of Ohio"
- ↑ Kjellesvig-Waering, Erik N.; Leutze, Willard P. (1966). «Eurypterids from the Silurian of West Virginia». Journal of Paleontology. 40 (5): 1109–1122. JSTOR 1301985
- ↑ Sven Laufeld; Roland Skoglund (1979). Lower Wenlock faunal and floral dynamics – Vattenfallet section, Gotland (PDF). [S.l.]: Geological Survey of Sweden. ISBN 978-9171581709
- ↑ a b Tetlie, O. Erik (2007). «Distribution and dispersal history of Eurypterida (Chelicerata)» (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. doi:10.1016/j.palaeo.2007.05.011. Arquivado do original (PDF) em 18 de julho de 2011
- ↑ Størmer, L. 1955. Merostomata. Treatise on Invertebrate Paleontology, Part P Arthropoda 2, Chelicerata, P: 30–31.
- ↑ a b Braddy, Simon J.; Poschmann, Markus; Tetlie, O. Erik (2007). «Giant claw reveals the largest ever arthropod». Biology Letters. 4 (1): 106–109. doi:10.1098/rbsl.2007.0491
- ↑ a b Plotnick, Roy E.; Baumiller, Tomasz K. (1988). «The pterygotid telson as a biological rudder». Lethaia. 21 (1): 13–27. doi:10.1111/j.1502-3931.1988.tb01746.x
- ↑ Gould, Gina C.; MacFadden, Bruce J. (2004). «Chapter 17: Gigantism, Dwarfism, and Cope's Rule: "Nothing in Evolution Makes Sense without a Phylogeny"». Bulletin of the American Museum of Natural History. 285: 219–237. doi:10.1206/0003-0090(2004)285<0219:C>2.0.CO;2
- ↑ a b c McCoy, Victoria E.; Lamsdell, James C.; Poschmann, Markus; Anderson, Ross P.; Briggs, Derek E. G. (1 de agosto de 2015). «All the better to see you with: eyes and claws reveal the evolution of divergent ecological roles in giant pterygotid eurypterids». Biology Letters. 11 (8). 20150564 páginas. PMC 4571687. PMID 26289442
- ↑ Paul A. «Autecology of Silurian eurypterids». Special Papers in Palaeontology. 32
- ↑ «Eurypterid-Associated Biota of the Patrick Burn Fm., Lesmahagow (Siltstones) (Silurian to of the United Kingdom) - Fossilworks». fossilworks.org. Consultado em 17 de dezembro de 2021
- ↑ «Eurypterid-Associated Biota of the Kip Burn Fm., Lesmahagow, Scotland (Silurian to of the United Kingdom) - Fossilworks». fossilworks.org. Consultado em 17 de dezembro de 2021
- ↑ «PBDB Collection». paleobiodb.org. Consultado em 10 de junho de 2025
Ligações externas
 Media relacionados com Erettopterus no Wikimedia Commons
Media relacionados com Erettopterus no Wikimedia Commons
| ||||||||||||||||||||||||||||||||
| Taxonomia |
|  | ||||||||||||||||||||||||||||||
| Geocronologia |
| |||||||||||||||||||||||||||||||
| Geografia |
| |||||||||||||||||||||||||||||||
| Gêneros notáveis | ||||||||||||||||||||||||||||||||
| Ichnotáxon |
| |||||||||||||||||||||||||||||||
| Grupos relacionados | ||||||||||||||||||||||||||||||||
| Artigos relacionados |
| |||||||||||||||||||||||||||||||