Adelophthalmidae
| Adelophthalmidae | |
|---|---|
| |
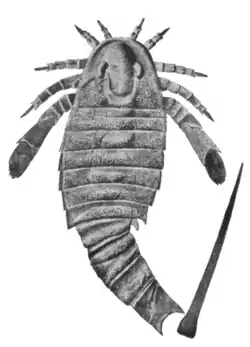
| Fóssil de Adelophthalmus mansfieldi | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Arthropoda |
| Subfilo: | Chelicerata |
| Ordem: | †Eurypterida |
| Superfamília: | †Adelophthalmoidea Tollerton, 1989 |
| Família: | †Adelophthalmidae Tollerton, 1989 |
| Espécie-tipo | |
| †Adelophthalmus granosus Jordan in Jordan & von Meyer, 1854
| |
| Generos | |
| |
| Sinónimos | |
| |
Adelophthalmidae (o nome derivando do gênero-tipo [en] Adelophthalmus, que significa "sem olhos óbvios") é uma família de euriptéridos, um grupo extinto de artrópodes aquáticos. Adelophthalmidae é a única família classificada como parte da superfamília Adelophthalmoidea, que por sua vez é classificada dentro da infraordem Diploperculata na subordem Eurypterina.
Euriptéridos adelophthalmídeos eram pequenos e nadadores que apareceram no período Siluriano. Com os membros mais antigos conhecidos do grupo, Nanahughmilleria prominens e Parahughmilleria maria, sendo conhecidos de depósitos da idade do Siluriano Inferior (possivelmente a época Llandovery) e os últimos membros, pertencentes ao gênero duradouro e difundido Adelophthalmus, extinguindo-se no Permiano Inferior, Adelophthalmidae é a família única de euriptéridos de maior duração. A sobrevivência do grupo, e de euriptéridos nadadores (a subordem Eurypterina) além do Devoniano Superior, deve-se inteiramente à sobrevivência e subsequente sucesso de Adelophthalmus ao longo do Devoniano e Carbonífero. Adelophthalmus (e possivelmente Unionopterus) representa o único gênero conhecido de euriptéridos nadadores além da extinção do resto do grupo no Devoniano Superior, estendendo o alcance temporal do grupo em mais de cem milhões de anos.
Embora os últimos euriptéridos nadadores e os membros finais da subordem tradicionalmente mais bem-sucedida e numerosa Eurypterina, adelophthalmídeos não foram os últimos euriptéridos. Stylonurinos ou os "euriptéridos caminhantes" foram os últimos, sobrevivendo na família Hibbertopteridae [en] até o evento de extinção Permiano-Triássico ou pouco antes, alguns milhões de anos após a extinção de adelophthalmídeos no Permiano Inferior.
Descrição

O tamanho dos euriptéridos adelophthalmídeos variava de 4 centímetros a 32 cm,[2] sendo Nanahughmilleria clarkei a menor espécie e Adelophthalmus khakassicus, a maior. Os adelophthalmídeos eram relativamente pequenos em comparação com seus parentes, como o gigantesco pterigotídeo Jaekelopterus rhenaniae, que facilmente excedia 2 metros, e foi o maior artrópode já descoberto.[3]
Aadelophthalmídeos eram pequenos euriptéridos nadadores com uma carapaça (a placa dorsal da cabeça, Unionopterus possivelmente representando uma exceção) parabólica (aproximadamente em forma de U) e com olhos intramarginais (ocorrendo dentro da margem). A perna natatória (sexto membro) era do tipo Adelophthalmus, ou seja, com um sétimo podômero [en] (segmentos dos membros) longo e um oitavo podômero grosseiramente serrilhado. O corpo tinha uma diferenciação de segunda ordem na seção intermediária presente (ou seja, com "extensões" laterais no sétimo segmento do corpo) e com um telson (a divisão mais posterior do corpo) lanceolado (em forma de lança).[4][5]
A morfologia do grupo varia dependendo de quão derivado (evolutivamente avançado) o gênero era. De fato, do basal ("primitivo") Nanahughmilleria ao mais derivado Adelophthalmus, ocorreu uma diminuição gradual da espinhosidade (posse de espinhos) dos apêndices [en] (membros) e um aumento no tamanho das espátulas genitais (uma peça longa e plana na área genital). O ainda mais primitivo Pittsfordipterus provavelmente nem sequer possuía esta última.[4]
História da pesquisa
Os primeiros fósseis de adelophthalmídeos a serem descobertos foram os do gênero-tipo [en], Adelophthalmus. O paleontólogo alemão Hermann Jordan coletou o primeiro espécime da espécie Adelophthalmus granosus em Jägersfreude, em Saarland, Alemanha. O espécime seria descrito três anos depois por Jordan e Hermann von Meyer, que imediatamente reconheceram a natureza euriptérida dos fósseis pela grande semelhança da forma geral da carapaça e dos apêndices com a de Eurypterus. Uma das principais diferenças que Jordan e von Meyer notaram foi a aparente falta de olhos, o que dá nome a Adelophthalmus (que significa "sem olhos óbvios"[6]) e a toda a superfamília.[7] Essa característica é agora assumida como sendo devida a um artefato de preservação e que não estava presente em Adelophthalmus, nem em qualquer outro adelophthalmídeo.[8] Desde então, um total de 33 espécies[2][9][10][11] foram descritas, algumas das quais foram historicamente classificadas em outros gêneros (Anthraconectes, Glyptoscorpius, Lepidoderma e Polyzosternites, todos agora sinônimos de Adelophthalmus),[8] tornando Adelophthalmus o gênero de euriptérido mais diverso até hoje.[9] No entanto, muitas dessas espécies são fragmentárias e poderiam representar sinônimos de outras espécies dentro de Adelophthalmus ou mesmo espécies de outros gêneros.[12] É possível que a grande quantidade de espécies em Adelophthalmus eventualmente provoque sua separação em dois ou três gêneros separados.[13]
Em 1948, o paleontólogo e geólogo ucraniano Boris Isidorovich Chernyshev descreveu um novo gênero conhecido por um único espécime do Cazaquistão (na época parte da União Soviética) e o classificou provisoriamente como parte da família Pterygotidae.[14] Foi nomeado Unionopterus, e sua classificação é controversa devido às ilustrações pobres dadas por Chernyshev e ao fato de que o único fóssil conhecido presume-se perdido. O gênero foi tratado como um euriptérido indeterminado (Incertae sedis), um adelophthalmídeo ou um membro do gênero Adelophthalmus, bem como completamente ignorado. A espinhosidade de seus apêndices pode sugerir a segunda opção, mas isso não é totalmente certo e a classificação filogenética de Unionopterus pode nunca ser resolvida.[4]
Em 1961, o paleontólogo americano Erik Norman Kjellesvig-Waering considerou várias espécies do gênero Hughmilleria suficientemente diferentes da espécie-tipo para serem separadas em um novo subgênero, que ele nomeou Nanahughmilleria. Essas espécies compartilhavam uma característica chave, pequenos olhos intramarginais de forma reniforme (em forma de feijão), em contraste com os grandes olhos ovóides localizados na margem da carapaça presentes no genótipo e suas formas aliadas. Kjellesvig-Waering designou H (Nanahughmilleria) norvegica como a espécie-tipo do novo subgênero. Ao mesmo tempo, ele erigiu um novo gênero, Parahughmilleria, com Parahughmilleria salteri como a espécie-tipo. Kjellesvig-Waering baseou seu novo clado principalmente na presença de lobos suplementares (lobos distais em um segmento em forma de placa que contém a abertura genital, chamado opérculo genital), bem como nos olhos intramarginais.[15]

Em 1964, Kjellesvig-Waering descreveu o opérculo genital da espécie H. (N.) phelpsae e sugeriu que esta parte do corpo poderia ter grande importância filogenética no futuro.[16] Dois anos depois, juntamente com o paleontólogo americano Kenneth Edward Caster, ele elevou H. (N.) phelpsae ao nível genérico sob o nome Pittsfordipterus. Eles também descreveram um novo gênero e espécie, Bassipterus virginicus. Ele difere essencialmente de outros adelophthalmídeos pela especialização de seu apêndice genital, com características reminiscentes de Eurypterus.[17]
Em 1989, Victor P. Tollerton Jr. descreveu a família Adelophthalmidae juntamente com muitas outras, incluindo Adelophthalmus, Parahughmilleria, Bassipterus e Unionopterus. Este clado foi baseado na presença de espinhos no segundo a quinto par de apêndices, uma perna natatória do tipo Adelophthalmus e a presença de epímeros (extensões laterais do segmento) no sétimo segmento. Tollerton comentou que algumas espécies de Adelophthalmus que não tinham espinhos nos apêndices poderiam ser melhor colocadas em um novo gênero na família Slimonidae (ele mencionou a agora inválida Slimonioidea).[18] Embora um novo gênero para espécies sem espinhos pudesse ser filogeneticamente apoiado, movê-lo para Slimonidae com base na perda de uma característica que parece ter sido perdida separadamente nos dois grupos não está de acordo com a prática comum.[4]
Odd Erik Tetlie, em uma tese não publicada de 2004, erigiu a superfamília Adelophthalmoidea e a família Nanahughmilleridae. Adelophthalmoidea foi diagnosticada como euriptéridos com carapaças parabólicas, pequenos olhos reniformes, apêndices de espinhosidade variável e um telson lanceolado, entre outros.[19] Esta superfamília seria formalmente descrita dois anos depois por Tetlie e Peter Van Roy. Por outro lado, Nanahughmilleridae foi descrita para conter adelophthalmoides sem ou com espátulas genitais reduzidas e o segundo a quinto par de apêndices prossomais (do prossoma, "cabeça") do tipo Hughmilleria. Esta família continha Nanahughmileria, Pittsfordipterus e talvez Parahughmilleria.[19] No entanto, o clado quase nunca[4] foi usado em estudos e listas subsequentes de euriptéridos, e em vez disso, eles classificam nanahughmillerídeos como parte de Adelophthalmidae.[9]

Em 2008, Nanahughmilleria patteni foi reconhecida como uma espécie diferente e muito mais basal e, portanto, um novo gênero, Eysyslopterus, foi nomeado por Tetlie e Markus Poschmann. A carapaça de Eysyslopterus e de outros membros basais de Pterygotioidea (Herefordopterus), que são parentes próximos, e de waeringopteroides ( Orcanopterus) mostrou-se quase idêntica, diferindo entre eles apenas pela posição dos olhos. Devido à posição intramarginal dos olhos, Eysyslopterus foi classificado dentro de Adelophthalmidae, mas também foi sugerido que seja o grupo-irmão (parente mais próximo) de um clado formado por Adelophthalmoidea e Pterygotioidea. No entanto, isso não pode ser demonstrado até que mais material fóssil seja encontrado.[12]
Em 2023, Archopterus anjiensis [en], um novo gênero e espécie, foi descrito com base em um único espécime encontrado na formação Wenchang, de idade Ordoviciana, de Zhejiang, China, representando o mais antigo registro conhecido de Adelophthalmidae, estendendo o registro estratigráfico da família em cerca de 10 milhões de anos, do início do Siluriano para o final do Ordoviciano.[20]
História evolutiva

Euriptéridos como grupo atingiram o pico de diversidade durante o Siluriano;[21][22] das aproximadamente 250 espécies válidas contabilizadas até hoje, cerca de 139 (≈56 %) eram apenas do Siluriano.[9] Muitos grupos de euriptéridos são registrados pela primeira vez no Siluriano, como Pterygotioidea, Mycteropoidea, Stylonuroidea [en] e Adelophthalmoidea. Os membros mais primitivos de Adelophthalmoidea evoluíram na Laurússia (um antigo supercontinente, também conhecido como Euramérica). De fato, a espécie mais basal do clado até agora (Eysyslopterus patteni) foi recuperada de depósitos Ludlovianos (cerca de 427–423 Ma) do paleocontinente Báltica (Escandinávia e Leste Europeu, precisamente a Estônia). No entanto, não é possível determinar onde o clado se originou, provavelmente na Báltica ou na Laurência (a maior parte do leste continental da América do Norte). Embora a maioria dos representantes da Adelophthalmoidea tenha sido encontrada na Laurência, Avalônia (Alemanha, Grã-Bretanha, partes do leste da América do Norte) e Báltica (ou seja, Laurússia), o gênero quase cosmopolita (mundial) Adelophthalmus também estava presente no Terreno Reno-Hercínico [en] (Europa ocidental e central), Sibéria e na parte de Gondwana da atual Austrália.[21]
No Siluriano, a maioria dos gêneros de adelophthalmídeos apareceria, mas todos se extinguiram logo depois ou no Devoniano Médio.[21] Os representantes mais antigos do grupo foram Parahughmilleria maria e Nanahughmilleria prominens, ambos do Llandovery (cerca de 444–433 Milhões de anos), sugerindo que adelophthalmídeos apareceram pela primeira vez por volta desta época. Isso é apoiado pelo aparecimento de pterigotioides basais, o grupo-irmão dde adelophthalmoides, no Llandovery. No entanto, o registro fóssil de Adelophthalmoidea é muito pobre no Siluriano Inferior e ambas as espécies têm idades contestadas. O representante inequivocamente mais antigo foi P. hefteri, com fósseis encontrados na formação Kip Burn [en], Lesmahagow [en], Escócia, do início da época Wenlockiana (cerca de 433–427 Milhões de anos). Esses fósseis diferem ligeiramente nas proporções do corpo dos fósseis de sua localidade-tipo (na Alemanha) e foram comparados com estes últimos, mas a atribuição dos fósseis escoceses a P. hefteri não é totalmente certa. As primeiras espécies viviam em águas salobras-estuarinas ou em habitats totalmente marinhos.[12]
No Emsiano (cerca de 393–408 Ma, no Devoniano Inferior), a primeira espécie de Adelophthalmus apareceu, Adelophthalmus sievertsi, apresentando características basais como a perna natatória larga (como em Nanahughmilleria e Parahughmilleria).[23] Os euriptéridos foram um dos grupos mais afetados pelo evento de extinção do Devoniano Superior. Após um grande declínio na diversidade durante o Devoniano Inferior, os euriptéridos eram raros em ambientes marinhos no Devoniano Superior. Das 16 famílias de euriptéridos que estavam vivas no início do Devoniano, apenas três persistiram no Carbonífero, todas elas grupos não marinhos.[24] A subordem Eurypterina foi quase completamente extinta, sobrevivendo apenas a Adelophthalmoidea (representada por Adelophthalmus).[25]

Adelophthalmus se diversificaria rapidamente, já estando presente na Sibéria e em Gondwana (Austrália) desde o Devoniano. No Carbonífero, a distribuição de Adelophthalmus tornou-se aproximadamente circum-equatorial (ao redor do Equador).[21] Das 33 espécies referidas a Adelophthalmus, 23 (69%) eram apenas do Carbonífero,[9][11] atingindo seu pico de diversidade no Carbonífero Superior[13] e tornando-se o mais comum de todos os euriptéridos do Paleozoico Superior.[4] Essa rápida diversificação pode ser devido à sua morfologia, convertendo os adelophthalmídeos em um dos nadadores mais hábeis entre os euriptéridos. A amálgama (união) da Pangeia em um supercontinente global também foi um fator importante. Os pterigotioides também foram nadadores bem-sucedidos, mas se extinguiram no Devoniano Médio, muito antes da formação da Pangeia.[21] Embora Unionopterus também tenha aparecido no Carbonífero, este gênero é muito pouco conhecido e geralmente é ignorado pelos pesquisadores de euriptéridos.[4]
Durante o Carbonífero Superior e o Permiano Inferior, Adelophthalmus vivia em ambientes de água salobra e doce adjacentes a planícies costeiras, um tipo de habitat comum e estável na época. Embora a formação da Pangeia tenha ajudado o gênero a se espalhar, também afetou seus ambientes. Eles começaram a desaparecer devido a uma mudança climática que causou alterações nos padrões de deposição e vegetação [en] em todo o mundo, provocando uma diminuição no número do gênero.[26] Adelophthalmus sellardsi da época Artinskiana (cerca de 290–284 Milhões de anos, Permiano Inferior) do Kansas, Estados Unidos, foi a última espécie de Adelophthalmus e, portanto, de toda a subordem Eurypterina.[3][9] O gênero expandiu o alcance temporal da subordem em cerca de 100 milhões de anos[25] e tornou sua família, a família única de euriptéridos de maior duração.[21] No entanto, os euriptéridos continuaram a existir representados pelos stylonurídeos. A espécie de hibbertoptérido [en] russa Campylocephalus permianus persistiu até o estágio Changhsingiano (cerca de 254–252 Milhões de anos, Permiano Superior), sendo o último euriptérido conhecido.[21] Nenhum euriptérido é conhecido de camadas fósseis posteriores ao Permiano, indicando que eles provavelmente morreram no evento de extinção em massa Permiano-Triássico ou pouco antes.[27]
Classificação
Filogenia externa
Adelophthamídeos são classificados dentro da infraordem Diploperculata, na subordem Eurypterina de euriptéridos.[9] A infraordem Diploperculata contém as quatro superfamílias mais derivadas de euriptéridos eurypterinos; Carcinosomatoidea, Adelophthalmoidea, Pterygotioidea e os waeringopteroides, unidos pela característica compartilhada de que o opérculo genital (a estrutura que contém o apêndice genital) é composto por dois segmentos fundidos.[28]
Adelophthalmoidea foi um dos clados de euriptéridos mais diversos taxonomicamente, com cerca de 40 espécies descritas. O grupo-irmão de Adelophthalmoidea, Pterygotioidea, superou essa quantidade com cerca de 50 espécies descritas, tornando-se a superfamília mais diversa até hoje. Esses clados-irmãos são os mais derivados na Eurypterina e compõem mais de um terço de todas as espécies de euriptéridos, com quase 100 espécies entre ambos.[21]
O cladograma abaixo é simplificado de um estudo de 2007 de O. Erik Tetlie, mostrando a posição de Adelophthalmoidea dentro da subordem Eurypterina.[21] A colocação de Diploperculata segue Lamsdell et al. 2013.[28]
| Eurypterida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Filogenia interna

Adelophthalmídeos têm uma série de características compartilhadas que os tornam diferentes do restante de euriptéridos.[4] No entanto, alguns gêneros desenvolveram características diferentes dentro de Adelophthalmidae que dividem a família em vários clados e agrupamentos menores. Os gêneros Parahughmilleria e Adelophthalmus formam um clado derivado com base na presença de espinhos aumentados em pelo menos um podômero no apêndice V (quinto membro), a presença de epímeros no pós-abdômen (segmentos corporais 8 a 12) e as grandes espátulas que foram associadas ao opérculo genital. Nanahughmilleria é colocado como o grupo-irmão deste clado, mas mais basal devido ao aumento da espinhosidade de seu apêndice V[12] e ao pequeno tamanho das espátulas genitais.[4] Bassipterus e Pittsfordipterus são posicionados como relativamente mais basais a este clado e formam um grupo apoiado por duas sinapomorfias (características compartilhadas diferentes das de seu último ancestral comum); olhos longos e estreitos e uma terminação complexa do apêndice genital.[12]
Na base da família, Eysyslopterus foi interpretado como o mais basal adelophthalmídeo. A carapaça deste gênero, Herefordopterus e Orcanopterus, eram quase idênticas e eram principalmente diferenciadas pela posição dos olhos. No último, os olhos eram quase marginais, mas eram separados da margem pela borda marginal. Os olhos de Herefordopterus eram completamente marginais, uma característica presente em todos os gêneros pterigotioides. Embora os olhos de Eysyslopterus fossem intramarginais, eles estavam muito mais próximos da margem do que em seus parentes, sugerindo que os olhos migraram gradualmente para o centro da carapaça dos gêneros basais para o derivado Adelophthalmus. A posição de seus olhos levou alguns pesquisadores a questionar se Eysyslopterus é mesmo um adelophthalmídeo ou um grupo-irmão de Adelophthalmidae, mas mais fósseis são necessários para provar ou refutar sua posição filogenética dentro da família ou fora dela.[12]
O cladograma abaixo apresenta as posições filogenéticas inferidas da maioria dos gêneros incluídos nas três superfamílias mais derivadas da subordem Eurypterina de euriptéridos (Adelophthalmoidea, Pterygotioidea e os waeringopteroides), conforme inferido por O. Erik Tetlie e Markus Poschmann em 2008, com base nos resultados de uma análise de 2008 especificamente pertencente à Adelophthalmoidea e uma análise anterior de 2004.[12]
| Diploperculata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecologia

Adelophthalmídeos como um todo habitavam ambientes situados perto do domínio costeiro, com preferências em lagoas, estuários ou deltas, que têm salinidade reduzida. Uma influência marinha é comumente encontrada em horizontes [en] (seções [en] do sítio fóssil) que compreendem euriptéridos, mas fósseis-índice marinhos (fósseis que indicam um ambiente e ecossistema marinho) em muitos casos não são associados aos restos de euriptéridos. Em alguns casos, onde os adelophthalmídeos são muito raros, os fósseis podem ter sido depositados em um lugar diferente daquele de onde se originaram,[12] exemplificado por Adelophthalmus waterstoni, que é conhecido por um único espécime que, por sua vez, é o único euriptérido da zona, a formação Gogo [en] da Austrália, onde mais de 2.000 crustáceos foram encontrados.[29]
Os primeiros adelophthalmídeos, por exemplo, o escocês Parahughmilleria hefteri, foram conservados em habitats estuarinos-salobros não marinhos, com possível influência de marés, embora formas basais que habitavam depósitos completamente marinhos sejam conhecidas. Essa preferência permaneceu constante em Adelophthalmidae até o aparecimento de Adelophthalmus. Em depósitos do Devoniano Inferior da Alemanha, onde Parahughmilleria hefteri e Adelophthalmus sievertsi coexistiram, uma influência marinha mais óbvia é observada nas seções dominadas por Parahughmilleria do que naquelas dominadas por Adelophthalmus. Isso sugere que Adelophthalmus tinha uma preferência por habitats dominados por água doce, ao contrário de outras formas mais basais. Essa preferência por habitats de água doce é mais forte durante as épocas Bashkiriana e Moscoviana (no Carbonífero Superior), épocas em que fósseis de Adelophthalmus foram associados a bivalves de água doce e organismos terrestres em estratos contendo carvão (indicando um ambiente de pântano de carvão).[12]
No entanto, isso pode estar relacionado à diversificação de Adelophthalmus nessas épocas mais do que a uma mudança ecológica em todo o gênero. A maioria das espécies de Adelophthalmus estava confinada a bacias parálicas (em águas rasas perto da costa) ou de planície, em ambientes deposicionais que tinham uma conexão próxima com habitats marginalmente marinhos. Por exemplo, durante o Moscoviano, a bacia do Saar-Nahe [en] (onde fósseis de Adelophthalmus granosus foram encontrados), estava conectada ou mesmo fazia parte de uma vasta área de subsidência ocidental (uma zona afundada) cuja drenagem ia para o Oceano Paleo-Tethys [en], cerca de 1.500 quilômetros ao sul. Durante o Pensilvaniano Superior e o Permiano Inferior, devido ao soerguimento do sul, a drenagem começou a ir para o norte, em direção ao Oceano Panthalassa, aumentando a distância para o oceano (cerca de 2.400 km). Adelophthalmus não é encontrado nesses depósitos, enquanto outros indicativos de uma conexão com um habitat marinho marginal, como os xifossúrios belinurídeos [en], persistiram. Algumas das últimas ocorrências dos adelophthalmídeos (final do Carbonífero e Permiano Inferior) parecem ter tido uma espécie de reversão nas preferências ecológicas, habitando novamente ambientes com forte influência marinha, especialmente os ambientes estuarinos com influência de marés.[12]
Adelophthalmidae é considerado um clado eurialino (capaz de se adaptar a diferentes salinidades) que habitava habitats estressantes com relação à paleossalinidade [en] e possivelmente oxigênio, cuja conquista de habitats de água doce não foi tão bem-sucedida quanto em outros animais, como os já mencionados bellinurídeos.[12]
Referências
- ↑ Lamsdell, J. C. (2025). «Codex Eurypterida: a revised taxonomy based on concordant parsimony and Bayesian phylogenetic analyses». Bulletin of the American Museum of Natural History. 473: 1–196. doi:10.1206/0003-0090.473.1.1
 . hdl:2246/7507
. hdl:2246/7507
- ↑ a b Shpinev, Evgeniy S.; Filimonov, A. N. (2018). «A New Record of Adelophthalmus (Eurypterida, Chelicerata) from the Devonian of the South Minusinsk Depression». Paleontological Journal. 52 (13): 1553–1560. Bibcode:2018PalJ...52.1553S. doi:10.1134/S0031030118130129
- ↑ a b Lamsdell, James C.; Braddy, Simon J. (2009). «Cope's rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates». Biology Letters. 6 (2): 265–269. ISSN 1744-9561. PMC 2865068. PMID 19828493. doi:10.1098/rsbl.2009.0700. Supplemental material
- ↑ a b c d e f g h i Tetlie, O.E.; van Roy, P. (2006). «A reappraisal of Eurypterus dumonti Stainier, 1917 and its position within the Adelophthalmidae Tollerton, 1989» (PDF). Bulletin de l'Institut Royal des Sciences Naturelles de Belgique. 76: 79–90
- ↑ Tollerton, V. P. (1989). «Morphology, Taxonomy, and Classification of the Order Eurypterida Burmeister, 1843». Journal of Paleontology. 63 (5): 642–657. Bibcode:1989JPal...63..642T. JSTOR 1305624. doi:10.1017/S0022336000041275
- ↑ «The ventral anatomy of the Upper Carboniferous eurypterid Anthraconectes Meek and Worthen | The Palaeontological Association». www.palass.org (em inglês). Consultado em 14 de maio de 2018
- ↑ Jordan, H. & von Meyer, H. 1854. "Ueber die Crustaceen der Steinkohlenformation von Saarbrücken". Palaeontographica 4: 1–15.
- ↑ a b Tetlie, O. Erik; Dunlop, Jason A. (1 de novembro de 2005). «A redescription of the Late Carboniferous eurypterids Adelophthalmus granosus von Meyer, 1853 and A. zadrai Přibyl, 1952». Fossil Record (em inglês). 8 (1): 3–12. ISSN 1860-1014. doi:10.1002/mmng.200410001
- ↑ a b c d e f g Dunlop, J. A.; Penney, D.; Jekel, D. (2018). «A summary list of fossil spiders and their relatives» (PDF). World Spider Catalog. [S.l.]: Natural History Museum Bern
- ↑ Shpinev, Evgeniy S. (2012). «New species of the genus Adelophthalmus (Eurypterida, Chelicerata) found in the Middle Devonian of Khakassia». Paleontological Journal. 46 (5): 470–475. Bibcode:2012PalJ...46..470S. doi:10.1134/S0031030112050103
- ↑ a b Lamsdell, James C.; McCoy, Victoria E.; Perron-Feller, Opal A.; Hopkins, Melanie J. (2020). «Air Breathing in an Exceptionally Preserved 340-Million-Year-Old Sea Scorpion». Current Biology. 30 (21): 4316–4321. PMID 32916114. doi:10.1016/j.cub.2020.08.034
- ↑ a b c d e f g h i j k Erik Tetlie, O; Poschmann, Markus (1 de junho de 2008). «Phylogeny and palaeoecology of the Adelophthalmoidea (Arthropoda; Chelicerata; Eurypterida)». Journal of Systematic Palaeontology. 6 (2): 237–249. Bibcode:2008JSPal...6..237T. doi:10.1017/S1477201907002416
- ↑ a b Lamsdell, James C.; Simonetto, Luca; Selden, Paul A. (31 de julho de 2013). «First Eurypterid from Italy: A new species of Adelophthalmus (Chelicerata: Eurypterida) from the Upper Carboniferous of the Carnic Alps (Friuli, NE Italy)». Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy). 119 (2). ISSN 2039-4942. doi:10.13130/2039-4942/6029
- ↑ Chernyshev, Boris I. (1948). «New representative of Merostomata from the Lower Carboniferous». State University of Kiev, Geological Collections. 2: 119–130
- ↑ Kjellesvig-Waering, Erik N. (1961). «The Silurian Eurypterida of the Welsh Borderland». Journal of Paleontology. 35 (4): 789–835. JSTOR 1301214
- ↑ Kjellesvig-Waering, Erik N. (1964). «Eurypterida: Notes on the Subgenus Hughmilleria (Nanahughmilleria) from the Silurian of New York». Journal of Paleontology. 38 (2): 410–412. JSTOR 1301566
- ↑ Kjellesvig-Waering, Erik N.; Leutze, Willard P. (1966). «Eurypterids from the Silurian of West Virginia». Journal of Paleontology. 40 (5): 1109–1122. JSTOR 1301985
- ↑ Tollerton, V. P. (1989). «Morphology, taxonomy, and classification of the order Eurypterida Burmeister, 1843». Journal of Paleontology (em inglês). 63 (5): 642–657. Bibcode:1989JPal...63..642T. ISSN 0022-3360. doi:10.1017/S0022336000041275
- ↑ a b Tetlie, Odd Erik (2004). Eurypterid phylogeny with remarks on the origin of arachnids (PhD). University of Bristol. pp. 1–344. Consultado em 1 de novembro de 2018. Arquivado do original em 30 de julho de 2021
- ↑ Wang, Han; Braddy, Simon J.; Botting, Joseph; Zhang, Yuandong (10 de maio de 2023). «The first documentation of an Ordovician eurypterid (Chelicerata) from China». Journal of Paleontology (em inglês). 97 (3): 606–611. Bibcode:2023JPal...97..606W. ISSN 0022-3360. doi:10.1017/jpa.2023.21
- ↑ a b c d e f g h i O. Erik Tetlie (2007). «Distribution and dispersal history of Eurypterida (Chelicerata)» (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. Bibcode:2007PPP...252..557T. doi:10.1016/j.palaeo.2007.05.011. Arquivado do original (PDF) em 18 de julho de 2011
- ↑ O'Connell M. «Chapter I». The Habitat of the Eurypterida. [S.l.: s.n.]
- ↑ Poschmann, Markus (1 de janeiro de 2006). «The Eurypterid Adelophthalmus Sievertsi (chelicerata: Eurypterida) from the Lower Devonian (emsian) Klerf Formation of Willwerath, Germany». Palaeontology (em inglês). 49 (1): 67–82. Bibcode:2006Palgy..49...67P. ISSN 1475-4983. doi:10.1111/j.1475-4983.2005.00528.x
- ↑ Mass Extinctions and Their Aftermath. [S.l.]: Oxford University Press. 1997. pp. 70. ISBN 978-0198549161
- ↑ a b Lamsdell, James C.; Braddy, Simon J. (2009). «Cope's Rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates». Biology Letters. 6 (2): 265–269. PMC 2865068. PMID 19828493. doi:10.1098/rsbl.2009.0700
- ↑ Kues, Barry S.; Kietzke, Kenneth K. (1981). «A Large Assemblage of a New Eurypterid from the Red Tanks Member, Madera Formation (Late Pennsylvanian-Early Permian) of New Mexico». Journal of Paleontology. 55 (4): 709–729. JSTOR 1304420
- ↑ Bergstrom, Carl T.; Dugatkin, Lee Alan (2012). Evolution. [S.l.]: Norton. ISBN 978-0393913415
- ↑ a b Lamsdell, James; Hoşgör, Izzet; Selden, Paul (31 de janeiro de 2013). «A new Ordovician eurypterid (Arthropoda: Chelicerata) from southeast Turkey: Evidence for a cryptic Ordovician record of Eurypterida». Gondwana Research. 23 (1): 354–366. Bibcode:2013GondR..23..354L. doi:10.1016/j.gr.2012.04.006
- ↑ Tetlie, O. Erik; Braddy, Simon J.; Butler, Piers D.; Briggs, Derek E. G. (2004). «A New Eurypterid (Chelicerata: Eurypterida) from the Upper Devonian Gogo Formation of Western Australia, With A Review of the Rhenopteridae». Palaeontology (em inglês). 47 (4): 801–809. Bibcode:2004Palgy..47..801T. ISSN 0031-0239. doi:10.1111/j.0031-0239.2004.00390.x
| ||||||||||||||||||||||||||||||||
| Taxonomia |
| | ||||||||||||||||||||||||||||||
| Geocronologia |
| |||||||||||||||||||||||||||||||
| Geografia |
| |||||||||||||||||||||||||||||||
| Gêneros notáveis | ||||||||||||||||||||||||||||||||
| Ichnotáxon |
| |||||||||||||||||||||||||||||||
| Grupos relacionados | ||||||||||||||||||||||||||||||||
| Artigos relacionados |
| |||||||||||||||||||||||||||||||