Slimonia
| Slimonia | |
|---|---|

| |
| Fóssil de S. acuminata | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Arthropoda |
| Subfilo: | Chelicerata |
| Ordem: | †Eurypterida |
| Família: | †Slimonidae |
| Gênero: | †Slimonia Page, 1856 |
| Espécie-tipo | |
| †Slimonia acuminata Salter, 1856
| |
| Espécies | |
| |
Slimonia[1] é um gênero de euriptérido, um grupo extinto de artrópodes aquáticos. Fósseis de Slimonia foram encontrados em depósitos do Siluriano na América do Sul e na Europa. Classificado como parte da família Slimonidae, ao lado do gênero relacionado Salteropterus, o gênero inclui três espécies válidas: S. acuminata de Lesmahagow [en], Escócia, S. boliviana de Cochabamba, Bolívia, e S. dubia das Colinas de Pentland [en], Escócia, além de uma espécie duvidosa (nomen dubium), S. stylops, de Herefordshire, Inglaterra. O nome genérico deriva e homenageia Robert Slimon, um coletor de fósseis e cirurgião de Lesmahagow.
Das quatro espécies descritas de Slimonia, três tinham comprimento igual ou inferior a 20 cm. Apenas S. acuminata era maior, com os maiores espécimes atingindo 100 cm de comprimento. Embora isso seja grande para um artrópode predador, Slimonia foi superada em tamanho por membros posteriores e mais derivados (mais "avançados") da família Pterygotidae de euriptéridos, que se tornaram os maiores artrópodes conhecidos.
Descrição
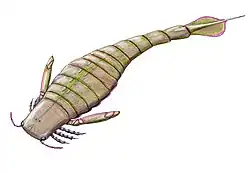
Slimonia apresenta muitas semelhanças com euriptéridos mais derivados (mais "avançados") de sua superfamília, a Pterygotioidea. Em particular, o télson (o segmento mais posterior do corpo) expandido e achatado de Slimonia é semelhante ao dos euriptéridos da família Pterygotidae, uma característica compartilhada apenas com alguns euriptéridos derivados da família Hibbertopteridae [en], onde a característica evoluiu convergentemente.[1] O télson de Pterygotidae era geralmente um pouco maior que o de Slimonia e mais delgado. No entanto, a espícula do télson de Slimonia era muito mais longa que qualquer uma observada nos Pterygotidae (constituindo pouco mais da metade do comprimento total do télson), serrilhada e terminando em uma ponta fina.[2] A maior espécie de Slimonia, S. acuminata, atingia um comprimento máximo de 100 cm, enquanto a menor, S. dubia, crescia até 12 cm.[3] Embora 100 cm seja grande para um artrópode predador, Slimonia foi superada em comprimento por membros posteriores e mais derivados da família Pterygotidae, que se tornaram os maiores artrópodes conhecidos.[4]
Slimonia pode ser distinguida de outros membros de sua família, a Slimonidae, por várias características. O prossoma (cabeça) tem forma quadrada e possui pequenos olhos compostos nos cantos frontais. O corpo era grande e cordiforme (em forma de coração), com um pós-abdômen estreito e um télson com a metade anterior fortemente expandida. As quelíceras (apêndices frontais) eram pequenas em comparação com as dos espécimes da família Pterygotidae, e as pernas tinham dentículos, mas sem espinhos. Os apêndices genitais eram longos e estreitos em machos e fêmeas.[5]
História da pesquisa
_(7394026154).jpg)
A espécie-tipo de Slimonia, S. acuminata, foi descrita inicialmente como uma espécie de Pterygotus, "Pterygotus acuminata" (acuminata sendo latim para "afiado" ou "afunilado"), por John William Salter [en] em 1856, com base em fósseis recuperados de depósitos das épocas Llandovery-Wenlock (Siluriano Inferior a Médio) em Lesmahagow, Escócia. No mesmo ano, David Page [en] criou um novo gênero para a espécie, pois várias características distintas a diferenciavam de outras espécies conhecidas de Pterygotus, incluindo a forma da carapaça e a ausência das grandes garras típicas de Pterygotus.[6] O nome genérico deriva e homenageia Robert Slimon, coletor de fósseis e cirurgião de Lesmahagow, que foi o primeiro a descobrir fósseis de euriptéridos em Lesmahagow, chamando a atenção de Roderick Murchison em 1851.[7] S. acuminata permanece como a maior espécie conhecida, com os maiores espécimes medindo até 100 cm de comprimento.[3]
Em 1899, uma espécie adicional, S. dubia, foi incluída no gênero. Essa espécie foi recuperada de depósitos ligeiramente mais antigos (Llandovery) nas Colinas de Pentland, Escócia, e podia ser distinguida de S. acuminata pelo télson mais alongado (menos largo nas partes posteriores), espícula do télson mais fina e uma forma corporal ligeiramente diferente, afunilando uniformemente em vez de estreitar repentinamente próximo ao sétimo segmento, como em S. acuminata.[8][9] O espécime-tipo de S. dubia é uma carapaça mal preservada, com fragmentos de vários graus de completude dos primeiros onze segmentos associados. Apesar de sua natureza fragmentária, a forma quadrangular da carapaça e os olhos posicionados em seus cantos permitiram ao zoólogo e paleontólogo Malcolm Laurie [en] classificá-la em Slimonia ao descrevê-la em 1899.[8] O tamanho da carapaça sugere que a espécie atingia 12 cm de comprimento.[3]
Outra espécie, S. stylops, foi inicialmente considerada uma espécie de Pterygotus quando descrita por John William Salter em 1859. A natureza altamente fragmentária dos fósseis conhecidos torna a identificação precisa difícil e problemática. Apenas um espécime, a parte anterior de uma carapaça com os olhos compostos na margem, é conhecido e, embora se assemelhe a Slimonia, também poderia ser atribuído a Hughmilleria ou mesmo representar a carapaça de Salteropterus abbreviatus (um euriptérido da família Slimonidae próximo, conhecido apenas pelo télson e metastoma [en], uma grande placa do abdômen).[10] Os fósseis foram recuperados de depósitos da época Pridoli (Siluriano Superior) em Herefordshire, Inglaterra, e sugerem que a espécie crescia até 12 cm de comprimento.[3] Devido à sua natureza problemática, S. stylops é considerada um nomen dubium por pesquisadores modernos.[11]
Em 1973, outra espécie de Slimonia foi nomeada por Kjellesvig-Waering com base em um único fóssil recuperado por Eduardo Rodriguez da Formação Kirusillas [en], de idade Ludlow-Pridoli (Siluriano Superior), em Cochabamba, Bolívia. Nomeada S. boliviana, o holótipo (BLV15, depositado no Museu Nacional de História Natural da França) consiste em um télson bem preservado, típico do gênero, inflado lateralmente e com uma ponta terminal em forma de adaga. Era anteriormente coberto por pequenas escamas semilunares a "espinhosas", agrupadas em uma única fileira de grandes escamas marginais que formam uma borda serrilhada linear. Uma quilha dorsal sutil está presente ao longo do télson. Havia uma área triangular na base do télson, que poderia ser um ponto de união com os músculos. S. boliviana diferia de S. acuminata por ter a quilha muito menos desenvolvida, mais estreita e não alcançando a espícula terminal. Esta era mais larga, menos pontiaguda e com serrilhas menos desenvolvidas. O próprio télson era mais largo e mais curto que na espécie-tipo. Essa espécie foi o terceiro euriptérido do Siluriano descrito no Hemisfério Sul, os outros dois provenientes da Austrália.[12] O fóssil sugere um comprimento corporal total de 20 cm.[3]
Classificação
_(7394030590).jpg)

Slimonia é classificado como parte da família de euriptéridos Slimonidae, dentro da superfamília Pterygotioidea.[11] Historicamente, Slimonia foi considerado um membro dos Pterygotidae, até ser reclassificado, junto com Hughmilleria e outros gêneros, para a família Hughmilleriidae em 1951 por Erik N. Kjellesvig-Waering.[13] Nestor Ivanovich Novozhilov [en] classificou Slimonia como parte de uma família própria em 1968.[11]
Slimonia é um dos gêneros mais próximos da família Pterygotidae, e os Slimonidae são frequentemente interpretados como um grupo-irmão dos Pterygotidae. A outra família, Hughmilleriidae, também foi considerada o grupo-irmão mais próximo de Pterygotidae. A descoberta de Ciurcopterus, o gênero mais primitivo conhecido da família Pterygotidae, e estudos revelando que Ciurcopterus combina características de Slimonia (os apêndices são particularmente semelhantes) e de membros mais derivados da família, indicaram que os Slimonidae são mais próximos dos Pterygotidae do que os Hughmilleriidae.[14]
O cladograma abaixo é simplificado a partir de um estudo de O. Erik Tetlie (2007),[15] e mostra a posição de Slimonia em relação ao restante da subordem Eurypterina de euripterídeos, com a subordem Stylonurina como grupo externo.
| Eurypterida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia
Em 2017, W. Scott Persons IV e John Acorn relataram a descoberta de um espécime de S. acuminata, MB.A 863, na Formação Patrick Burn [en], Escócia, datado do Teliquiano, cerca de 430 milhões de anos atrás. O espécime era uma série completa e articulada de segmentos de télson, pós-abdominais e pré-abdominais, mostrando uma forte curvatura lateral no pós-abdômen. Persons e Acorn admitiram que ele poderia ter sofrido alguma desarticulação pós-morte ou representar uma muda parcial (exúvia), mas concluíram que, como não havia desarticulação aparente no metassoma, a articulação observada nos segmentos pós-abdominais (também vista em outros fósseis de euriptéridos, como Eurypterus e Alkenopterus) provavelmente era possível em vida.[2]
Estudos biomecânicos sobre os télsons e segmentos pós-abdominais de euriptéridos próximos a Slimonia, particularmente os da família Pterygotidae, revelaram que o corpo era muito rígido, e o télson achatado provavelmente funcionava como um leme, permitindo agilidade e curvas rápidas ao perseguir presas, contradizendo hipóteses anteriores de que o télson teria uma função propulsora.[1] Embora o pós-abdômen de Slimonia fosse provavelmente rígido e inflexível dorsalmente (para cima e para baixo), Persons e Acorn sugeriram que seu espécime indicava alta flexibilidade lateral (de lado a lado). Assim, teorizaram que a cauda poderia ser usada como arma. A espícula do télson, serrilhada nas laterais e mais longa que o télson achatado, termina em uma ponta afiada, e eles propuseram que poderia perfurar presas.[2][16]
No entanto, a teoria de Persons e Acorn foi contestada em 2018 por James Lamsdell, David Marshall e Derek Briggs. Embora o estudo de Persons e Acorn afirmasse que o fóssil não apresentava sinais de desarticulação, Lamsdell, Marshall e Briggs argumentaram que isso provavelmente não é verdade. Eles destacaram que os tergitos 8 e 10 claramente se sobrepunham aos outros de maneira não natural. Além disso, afirmaram que o espécime era definitivamente uma muda, não uma carcaça, sugerindo que a pose do fóssil não representava uma posição viável em vida. Também argumentaram que a quilha no télson de Slimonia criaria resistência significativa ao tentar varrer o télson lateralmente para perfurar presas. Por fim, sugeriram que as serrilhas no télson provavelmente eram pontos de fixação para cerdas que auxiliavam na percepção do fluxo de água, facilitando a navegação.[17]
A acuidade visual, a clareza da visão, pode ser determinada em artrópodes pelo número de lentes em seus olhos compostos e pelo ângulo interommatidial (IOA, a sigla em inglês para o ângulo entre os eixos ópticos das lentes adjacentes). O IOA é especialmente importante, pois pode distinguir diferentes papéis ecológicos em artrópodes, sendo baixo em predadores artrópodes ativos modernos.[18] Slimonia era muito semelhante ao gênero basal Erettopterus em termos de acuidade visual, com o número de lentes comparável ao de Pterygotus e Jaekelopterus e um IOA entre 2 e 3 (maior que o de Pterygotus e Jaekelopterus, sugerindo que a acuidade visual de Slimonia era boa, mas inferior à dos membros derivados de Pterygotidae).[18]
Paleoecologia

Fósseis de Slimonia foram recuperados em depósitos com faunas diversificadas de euriptéridos. Depósitos do Teliquiano nas Colinas de Pentland, onde restos de S. dubia foram encontrados, preservam fósseis de muitos outros euriptéridos, incluindo Drepanopterus pentlandicus [en], Laurieipterus elegans [en], Parastylonurus ornatus [en], Hardieopterus macrophthalmus [en], Carcinosoma scoticus, Stoermeropterus conicus [en] e Pentlandopterus minor [en]. Também estão preservados fósseis de espécimes da ordem Orthocerida [en], como Geisonoceras maclareni [en].[8] Níveis semelhantes de diversidade de euriptéridos são observados em depósitos fósseis onde outras espécies de Slimonia foram encontradas. S. acuminata foi associada a Nanahughmilleria lanceolata, H. lanarkensis [en], Eusarcana obesus, Parastylonurus sigmoidalis [en], C. scorpionis e Erettopterus bilobus.[19][20] S. stylops foi encontrada associada a Nanahughmilleria pygmaea, Eusarcana salteri, H. megalops, Erettopterus brodiei, Erettopterus gigas, Hughmilleria banksi, Eurypterus cephalaspis e Pterygotus ludensis.[21]
O ambiente de vida dos Pterygotidae variava de gênero para gênero, com alguns (como Pterygotus) encontrados em estuários, enquanto outros (como Jaekelopterus) habitavam ambientes de água doce; Slimonia foi encontrada em ambientes que parecem ser intertidais a marinhos. A Formação Patrick Burn, por exemplo, é considerada não marinha ou marginalmente marinha[22] ou apenas marinha.[23] Slimonia provavelmente predava peixes menores, pois não possuía as garras aumentadas dos Pterygotidae e era menor que os maiores membros desse grupo. As presas provavelmente incluíam peixes sem mandíbula, como Heterostraci e Osteostraci primitivos, que Slimonia capturava com seus apêndices frontais. Slimonia se movia em seu ambiente com pernas finas ou usando seus apêndices natatórios. Os pulmões do gênero estavam localizados na parte inferior do corpo, em uma série de dobras.[24]
Como muitas espécies de euriptéridos, Slimonia acuminata requer uma redescrição moderna para estabelecer características e traços definitivos. Alguns traços aparentemente únicos de S. acuminata foram descritos com base em espécimes no Museu e Galeria de Arte de Doncaster [en], incluindo fileiras de pústulas (protuberâncias) ao longo da margem do corpo e apêndices. Em alguns artrópodes, pústulas servem como pontos de fixação de cerdas (estruturas semelhantes a pelos com funções sensoriais). Fileiras de pústulas semelhantes foram descobertas no euriptérido Drepanopterus abonensis [en], um alimentador por varredura que usava a margem para procurar presas no substrato de seu ambiente. Se as pústulas de S. acuminata possuíam cerdas, poderiam funcionar como órgãos táteis e sensoriais para localizar e identificar presas, junto com os pedipalpos (o segundo par de apêndices, atrás das quelíceras).[25]
Ver também
Referências
- ↑ a b c Plotnick, Roy E.; Baumiller, Tomasz K. (1 de janeiro de 1988). «The pterygotid telson as a biological rudder». Lethaia (em inglês). 21 (1): 13–27. Bibcode:1988Letha..21...13P. ISSN 1502-3931. doi:10.1111/j.1502-3931.1988.tb01746.x
- ↑ a b c Persons, W. Scott; Acorn, John (2017). «A Sea Scorpion's Strike: New Evidence of Extreme Lateral Flexibility in the Opisthosoma of Eurypterids». The American Naturalist (em inglês). 190 (1): 152–156. PMID 28617636. doi:10.1086/691967
- ↑ a b c d e Lamsdell, James C.; Braddy, Simon J. (2009). «Cope's rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates». Biology Letters. 6 (2): 265–269. ISSN 1744-9561. PMC 2865068
 . PMID 19828493. doi:10.1098/rsbl.2009.0700. Supplemental material
. PMID 19828493. doi:10.1098/rsbl.2009.0700. Supplemental material
- ↑ Braddy, Simon J.; Poschmann, Markus; Tetlie, O. Erik (2007). «Giant claw reveals the largest ever arthropod». Biology Letters. 4 (1): 106–109. PMC 2412931. PMID 18029297. doi:10.1098/rsbl.2007.0491
- ↑ Størmer, L 1955. Merostomata. Treatise on Invertebrate Paleontology, Part P Arthropoda 2, Chelicerata, P: 30.
- ↑ Nicholson, Henry Alleyne (1 de janeiro de 1868). «III. On the Occurrence of Fossils in the Old Red Sandstone of Westmoreland». Transactions of the Edinburgh Geological Society (em inglês). 1 (1): 15–18. ISSN 0371-6260. doi:10.1144/transed.1.1.15
- ↑ Clarkson, Euan N.K.; Harper, David A.T. (2016). «Silurian of the Midland Valley of Scotland and Ireland». Geology Today. 32 (5). 195 páginas. Bibcode:2016GeolT..32..195C. doi:10.1111/gto.12152
- ↑ a b c Laurie, Malcolm (1900). "XIX.—On a Silurian Scorpion and some additional Eurypterid Remains from the Pentland Hills". Earth and Environmental Science Transactions of The Royal Society of Edinburgh. 39 (3): 575–590. doi:10.1017/S0080456800035109 ISSN 2053-5945
- ↑ Lamont, Archie (1 de janeiro de 1955). «Scottish Silurian Chelicerata». Transactions of the Edinburgh Geological Society (em inglês). 16 (2): 200–216. ISSN 0371-6260. doi:10.1144/transed.16.2.200
- ↑ Kjellesvig-Waering, Erik N. (1961). «The Silurian Eurypterida of the Welsh Borderland». Journal of Paleontology. 35 (4): 789–835. JSTOR 1301214
- ↑ a b c Dunlop, J. A., Penney, D. & Jekel, D. 2015. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch , version 16.0 http://www.wsc.nmbe.ch/resources/fossils/Fossils16.0.pdf (PDF).
- ↑ Kjellesvig-Waering, Erik N. (1973). «A new Silurian Slimonia (Eurypterida) from Bolivia». Journal of Paleontology. 47 (3): 549–550. JSTOR 1303202
- ↑ Kjellesvig-Waering, Erik N. (1964). «A Synopsis of the Family Pterygotidae Clarke and Ruedemann, 1912 (Eurypterida)». Journal of Paleontology. 38 (2): 331–361. JSTOR 1301554
- ↑ Tetlie, O. Erik; Briggs, Derek E. G. (1 de setembro de 2009). «The origin of pterygotid eurypterids (Chelicerata: Eurypterida)». Palaeontology (em inglês). 52 (5): 1141–1148. Bibcode:2009Palgy..52.1141T. ISSN 1475-4983. doi:10.1111/j.1475-4983.2009.00907.x
- ↑ O. Erik Tetlie (2007). «Distribution and dispersal history of Eurypterida (Chelicerata)» (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (3–4): 557–574. Bibcode:2007PPP...252..557T. doi:10.1016/j.palaeo.2007.05.011. Arquivado do original (PDF) em 18 de julho de 2011
- ↑ «A killer tail spine likely helped this ancient sea scorpion subdue its prey». Science | AAAS (em inglês). 17 de abril de 2017. Consultado em 18 de março de 2018
- ↑ James C. Lamsdell; David J. Marshall; Derek E. G. Briggs (2018). «Hit and Miss: (A Comment on Persons and Acorn, "A Sea Scorpion's Strike: New Evidence of Extreme Lateral Flexibility in the Opisthosoma of Eurypterids")». The American Naturalist. 191 (3): 352–354. doi:10.1086/695955
- ↑ a b McCoy, Victoria E.; Lamsdell, James C.; Poschmann, Markus; Anderson, Ross P.; Briggs, Derek E. G. (1 de agosto de 2015). «All the better to see you with: eyes and claws reveal the evolution of divergent ecological roles in giant pterygotid eurypterids». Biology Letters. 11 (8). 20150564 páginas. PMC 4571687. PMID 26289442. doi:10.1098/rsbl.2015.0564
- ↑ «Eurypterid-Associated Biota of the Patrick Burn Fm., Lesmahagow (Siltstones) (Silurian to of the United Kingdom) - Fossilworks». fossilworks.org. Consultado em 17 de dezembro de 2021
- ↑ «Eurypterid-Associated Biota of the Kip Burn Fm., Lesmahagow, Scotland (Silurian to of the United Kingdom) - Fossilworks». fossilworks.org. Consultado em 17 de dezembro de 2021
- ↑ «Eurypterid-Associated Biota, Platyschisma Beds, Downton Castle Sst., Herefordsh. (Silurian of the United Kingdom) - Fossilworks». fossilworks.org. Consultado em 17 de dezembro de 2021
- ↑ Tetlie, O. Erik; Braddy, Simon J. (2003). «The first Silurian chasmataspid, Loganamaraspis dunlopi gen. et sp. nov. (Chelicerata: Chasmataspidida) from Lesmahagow, Scotland, and its implications for eurypterid phylogeny». Earth and Environmental Science Transactions of the Royal Society of Edinburgh (em inglês). 94 (3): 227–234. ISSN 1473-7116. doi:10.1017/S0263593300000638
- ↑ Bicknell, Russell D. C.; Pates, Stephen (2020). «Pictorial Atlas of Fossil and Extant Horseshoe Crabs, With Focus on Xiphosurida». Frontiers in Earth Science. 8: 98. Bibcode:2020FrEaS...8...98B. ISSN 2296-6463. doi:10.3389/feart.2020.00098
- ↑ Fortey, Richard A. (1998). Life: A Natural History of the First Four Billion Years of Life on Earth. Nova York: Alfred A. Knopf. pp. 146–7. ISBN 978-0-375-40119-0
- ↑ Lomax, Dean; Lamsdell, James; Ciurca, Samuel (1 de janeiro de 2011). «A collection of eurypterids from the Silurian of Lesmahagow collected pre 1900». Geological Curator. 9 (6): 331–348. doi:10.55468/GC75
| ||||||||||||||||||||||||||||||||
| Taxonomia |
|  | ||||||||||||||||||||||||||||||
| Geocronologia |
| |||||||||||||||||||||||||||||||
| Geografia |
| |||||||||||||||||||||||||||||||
| Gêneros notáveis | ||||||||||||||||||||||||||||||||
| Ichnotáxon |
| |||||||||||||||||||||||||||||||
| Grupos relacionados | ||||||||||||||||||||||||||||||||
| Artigos relacionados |
| |||||||||||||||||||||||||||||||