Kuru kulla
| Kuru kulla | |
|---|---|
| |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Classe: | Reptilia |
| Clado: | Dinosauria |
| Clado: | Saurischia |
| Clado: | Theropoda |
| Família: | †Dromaeosauridae |
| Clado: | †Eudromaeosauria |
| Subfamília: | †Velociraptorinae |
| Gênero: | †Kuru Napoli et al., 2021 |
| Espécies: | †K. kulla
|
| Nome binomial | |
| †Kuru kulla Napoli et al., 2021
| |
| Sinónimos | |
| |
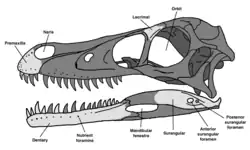
Kuru kulla (nomeado em homenagem á Kurukullā, uma deidade do lamaísmo) foi um gênero de dinossauro terópode maniraptorano do período Maastrichtiano do Cretáceo Superior. K. kulla é a única espécie do seu gênero, e é conhecida por um esqueleto que inclui um crânio parcial.
Descoberta e nomeação
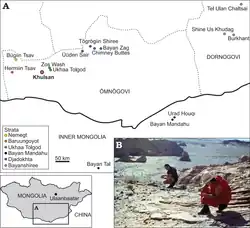
O holótipo de Kuru kulla foi encontrado por uma expedição America-Mongoliana em Khulsan no Deserto de Gobi, na Mongólia, em 5 de Julho de 1991, algumas horas antes do holótipo de Shri devi ser descoberto. O espécime foi redirecionado ao Centro de Geologia da Academia Mongoliana de Ciências, em Ulaanbaatar. O fóssil foi nomeado IGM 100/987 e consiste de um esqueleto que inclui uma pré-maxila direita, o osso lacrimal direito, uma mandíbula direita parcial, um osso surangular, 14 vértebras presacrais, 3 vértebras caudais, fragmentos de ambos os braços, um ílio fragmentado, os fins distais de ambas as púbis, pedaços de ambos os fêmures, a tíbia direita e fragmentos de ambos os pés. A qualidade de fossilização era inconsistente; fazendo o fóssil Kuru kulla ser de menor qualidade se comparado a Shri devi.[1]
Em 1992, o holótipo foi considerado um dromaeossaurídeo distinto numa conferência da Sociedade de Paleontologia dos Vertebrados.[2] Em um papel feito por Perle et al. descrevendo Achillobator, foi colocada uma unidade bibliográfica chamada (em tradução literal) "Morfologia de um dinossauro Dromaeossáurio - Airakoraptor do Cretáceo Superior Mongoliano".[3] No entanto, não há registros dessa unidade existindo, e é provável que se refira á um trabalho não-publicado, o que faz de "Airakoraptor" um nomen nudum. Napoli et al., em sua descrição formal de Kuru; presume que o nome "Airakoraptor" signifique "Ladrão de Kumis", uma bebida alcóolica da Ásia Central feita com leite de jumenta, cuja qual também é chamada de airag ou airak (airaque, aportuguesadamente). É explícitamente detalhado na descrição de Kuru kulla que seu nome não se refere á EET de mesmo nome.[4]
O osso surangular do holótipo foi referido na descrição de Tsaagan mangas.[5] No entanto, mais tarde no mesmo ano, o espécime foi confundido com um denominado "Espécime de Zos Wash" e foi catalogado como IGM 100/3503 e colocado no gênero Velociraptor.[1][6] O gênero foi finalmente nomeado e descrito por James Napoli, Alexander Ruebenstahl, Bhart-Anjan Bullar, Alan Turner e Mark Norell.[1]
Descrição

Kuru kulla era um eudromaeossauro de porte pequeno, de tamanho similar a Velociraptor. O esqueleto é incompleto, mas é similar aos seus parentes próximos, indicando que, em vida, se pareceria com animais como Velociraptor e Tsaagan mangas. É distinguível de outros velociraptorinos por autapomorfias como:
- Um sulco profundo na frente das narinas
- Uma protusão pequena parecida com um chifre no osso lacrimal
- Dois forâmens na traseira do osso surangular
- Ossos pneumáticos nas vértebras dorsais que estão projetadas para a frente se comparados com outros gêneros
Kuru tinha um crânio mais robusto, membros mais longos e uma "garra de foice" reduzida. Apesar de não haver estudos sobre sua anatomia funcional, essas diferenças indicam leves divergências ecológicas entre Kuru e seus relativos.[1]
Crânio
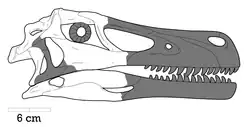
O crânio de Kuru é representado pela porção anterior de pré-maxila direita (na frente da narina) e a parte posterior do osso lacrimal direito. A pré-maxila tem apenas dois alvéolos dentários preservados, mas é certo que tinha quatro dentes pré-maxilares, como todos os terópodes dentados. Várias características da superfície óssea estão preservadas. A mais proeminente é um sulco em forma de "c", anterior á narina. Essa estrutura também está presente em Tsaagan mangas e Linheraptor, mas é mais pronunciada nesse gênero.
O osso lacrimal não está completamente preservado, mas, com base em outros dromeossaurídeos, provavelmente tinha formato de "T". Na parte anterior, a conexão do lacrimal com o osso nasal está preservada, e forma uma protusão que se conectaria com a parte ventral da narina em vida. Na parte dorsal posterior, há um "chifre" que fica ao lado da órbita. A parte dorsal dessa protusão possui uma textura áspera; a qual é observável em outros dromeossaurídeos. Uma excavação está presente na parte ventral do lacrimal, que foi interpretada como a passagem do canal lacrimal.[1]
A mandíbula remanescente é composta pelo osso suprangular direito e um maxilar inferior direito quase completo, mas os ossos não foram preservados continuamente e são feitos de vários cacos. Há também 9 dentes inferiores preservados e 3 que podem ser tanto do maxilar superior quanto do inferior. O maxilar inferior é reto, ao contrário de outros velociraptorinos e sendo similar á Dromaeosaurus e Deinonychus. Há 14 (talvez 15) posições preservadas; o que é muito mais alto do que as 11-12 de um dromeossaurídeo médio, mas Saurornitholestes tem um número de dentes parecido. Os dentes são zifodontes em formato (mais finos na medida médiolateral do que na anteroposterior) e tem raízes e serrações profundas nas bordas da frente e atrás, pelo menos nos dentes que estão preservados o suficiente para conter tantos detalhes. No entanto, o esmalte dentário serrilhado dos lados não está presente; ao contrário de outros dromeosaurídeos. Notávelmente, o terceiro dente é muito maior do que os seus "vizinhos", formando uma "presa" característica na mandíbula inferior. No entanto, o alvéolo do quarto dente é tão grande quanto o do terceiro, então é bastante provável que o quarto dente ainda estava crescendo e que, em vida, era tão grande quanto o terceiro.[1]
O osso surangular é o mais completo e têm vários traços diagnósticos do gênero Kuru. O traço mais notável é a presença de duas foraminas ao invés de uma. A prevalência desse traço em velociraptorinos é ambígua por causa da preservação incompleta e pobre dessa área do crânio em Tsaagan mangas e Linheraptor. Na descrição de Kuru, Napoli et al. comentaram sobre o tiranossaurídeo putativo Bagaraatan, que também tem essa característica, mas também lembraram que o espécime pode ser uma quimera e assim talvez não ofereça um valor real. Existe uma protusão inclinada dorsalmente no surangular, similar a Deinonychus, mas difere de todos os outros dromeossaurídeos por ser inclinada lateralmente.[1]
Esqueleto pós-cranial
Várias vértebras estão preservadas no holótipo, mas a maioria não está articulada e nem em linha, e várias preservam apenas o centro vertebral. Quatro vértebras cervicais, dez vértebras dorsais e três vértebras caudais; dezessete no total, estão preservadas. As posições das vértebras na coluna vertebral são desconhecidas; mas comparações com Tsaagan mangas e Shri devi deixaram Napoli et al. colocá-las em posições tentativas. Algumas vértebras dorsais ainda foram preparadas incorretamente, e puderam ser visualizadas apenas com o uso de tomografia computadorizada. Os centros dorsais aparentam encurtar da secção anterior á posterior.[1]
O braço de Kuru é composto pelos fins distais dos úmeros, um fragmento proximal do úmero direito, o fim proximal da ulna direita, ambos os fins do rádio direito, três metacarpos esquerdos atriculados com o carpo semilunar esquerdo, várias falanges isoladas e duas garras da mão. O úmero têm partes rugosas, que provavelmente são os locais aonde os músculos se anexavam. As ulnas não têm tubérculos (que foram encontrados em Velociraptor), mas é provável que seja um resultado de preservação de baixa qualidade. Há uma crista que se estende do processo coronoide da ulna, característica vista no táxon controverso Balaur. Kuru têm uma caraterística emblemática dos maniraptores, que é uma expansão plana no fim distal do rádio. Kuru não têm nenhuma característica nas mãos que o distingua de qualquer outro dromeosaurídeo.
As pernas são compostas pelo processo pós-acetabular esquerdo e o nervo ciático do ílio, os fins distais de ambos os púbis, ambos os fêmures, a ponta proximal da tíbia esquerda, os três metatarsos esquerdos e dois metatarsos direitos, o tálus direito, dezesseis falanges pedais de ambos os pés e quatro garras. As pernas são similares ás de Velociraptor e Shri devi, com os três táxons tendo uma crista aumentada na lateral do fêmur. O quarto trocanter está ausente em Kuru, mas há uma estrutura no fêmur que aparenta ser homóloga com a estrutura.[1]
Classificação
Em sua descrição de Kuru, Napoli et al. conduziram uma análise filogenética em vários gêneros de paravianos. Os dados usados foram tirados primariamente de um estudo conduzido por Steve Brusatte em 2014.[7] with the addition of several taxa scored by Pei and colleagues in 2020.[8] Napoli et al. ajustaram alguns atributos de Tsaagan mangas, Adasaurus e Velociraptor baseados em observações independentes.[1]
Na análise, Kuru foi colocado como um táxon-irmão de Adasaurus, da Formação Nemegt. As sinapomorfias dos dois gêneros incluem um forâmen no osso surangular posterior que escava 30% do osso, a falta do quarto trocanter e vértebras torácicas que são finas e longas. Kuru também têm uma "garra de foice" reduzida, similarmente á Adasaurus.[1] Um cladograma dos resultados pode ser visto abaixo:
| Eudromaeosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||

Uma filogenia mais recente incluindo Kuru feita for Łukasz Czenpiński em 2023 e veio junto com sua descrição de um crânio de Shri devi. Em sua análise, Czerpiński replicou aquelas feitas por Napoli et al.[1], conjuntamente com as de Power et al.[9], usando os dados que eles forneceram.[10] Notávelmente, Czerpiňski colocou Adasaurus fora de Velociraptorinae, mas também não colocou o gênero dentro de Dromaeosaurinae ou Saurornitholestinae.
Na análise filogenética feita em 2025 de Shri rapax, da Formação Djadokhta, Mourtille et al. reuniram Adasaurus e Kuru como táxons-irmãos, os colocando em uma politomia não-resolvida junto com Luanchuanraptor e Yurgovuchia. Esses resultados foram possíveis com a matriz focada em terópodes feita por Andrea Cau[11], que ajudou na descrição de S. rapax.[12]
Paleoecologia
Dieta
A dieta e ecologia de Kuru ainda não foi estudada completamente. No entanto, várias publicações mencionam a aparente existência de pelo menos 3 dromeossauros distintos na Formação Baruungoyot. Kuru, Shri devi e um animal atribuído á Velociraptor, mas que pode representar um táxon novo, estão incluídos.[1][13]
É especulado que esses animais tenham tido preferências de presas similares[1], mas é também possível que cada um tenha tido um nicho único, considerando a diferença dos crânios de S. devi e Kuru em comprimento.[10] Entretanto, pesquisadores apontam que os crânios são muito incompletos para qualquer hipótese concreta ser tomada.[13]
Paleoambiente
Kuru foi encontrado na área Khulsan da Formação Baruungoyot, que fica perto de Gurvan Tes, Ömnögovi, Mongólia.[1] Esse local é próximo de um afloramento da Formação Nemegt, cuja qual é acreditada a ser mais jovem do que a Formação Baruungoyot.[14][15]
Os sedimentos da formação foram depositados em várias condições, com a parte inferior sendo composto da dunas e lagos, que existiam em áreas entre as anteriores, enquanto a parte superior é composta de sedimentos que foram depositados em uma área similar á taquires que era inundada em intervalos regulares.[16] A formação é caracterizada por vários leitos vermelhos, majoritariamente areias de cores claras (amareladas, cinza-marrons e raramenta avermelhadas) que são cementadas localmente. Rochas de lama, siltitos e conglomerados arenosos, junto com estratificação cruzada em massa, também são rochas comuns. Também há arenitos disformes, médios, finos e extremamente finos são predominantes em sedimentos da formação.[16][17]
A geologia geral da formação indica que os sedimentos foram depositados em climas [[árido]s ou semiáridos em paleoambientes de planície aluvial, lago e erosão eólica, com outros corpos d'água de vida curta.[16][17]
Fauna contemporânea

A Formação Baruungoyot é similar á próxima Formação Djadokhta no quesito dos depósitos eólicos, que preservam animais pequenos e médios. Vários gêneros de lagartos são conhecidos dessas formações, incluíndo Gobiderma e Estesia. Mamíferos também eram abundantes na região, sendo representados por multituberculados como Catopsbaatar e Nemegtbaatar, metatérios como Deltatheridium, e eutérios como Asioryctes e Barunlestes.[18][19]
Dinossauros são o grupo mais bem-representado de vertebrados, com fósseis de anquilossaurídeos como Saichania, Tarchia, Zaraapelta e uma espécie indescrita de Hermiin Tsav.[20] Um crânio de saurópode também foi encontrado e assinalado como Quaesitosaurus.[21] marginocéfalos também estão presentes, sendo representados pelo paquicéfalossauro Tylocephale e pelo ceratópsio Bagaceratops, sendo este a espécie mais comum na formação.[22]
Os dinossauros com mais espécies são, definitivamente, os maniraptoranos. Há pelo menos quatro alvaressauroídeos (Ceratonykus, Khulsanurus, Ondogurvel e Parvicursor)[23] e vários oviraptorossauros, como Nemegtomaia, que também foi encontrada na formação Nemegt.[24] dromeossauros também eram comuns e incluíam Shri devi, Natovenator, Kuru kulla e um táxon não-descrito.[1][25] Os menores maniraptores presentes eram as aves, que incluíam a enigmática Hollanda, o mais bem-descrito Gobipteryx, e vários ovos de pássaro com embriões fossilizados.[26][27]
Referências
- ↑ a b c d e f g h i j k l m n o p Napoli, J. G.; Ruebenstahl, A. A.; Bhullar, B.-A. S.; Turner, A. H.; Norell, M. A. (2021). «A New Dromaeosaurid (Dinosauria: Coelurosauria) from Khulsan, Central Mongolia» (PDF). American Museum Novitates (3982): 1–47. ISSN 0003-0082. doi:10.1206/3982.1. hdl:2246/7286
- ↑ Norell, M. A.; Clark, J. M.; Perle, A. (1992). «New dromaeosaur material from the Late Cretaceous of Mongolia». Journal of Vertebrate Paleontology. 12 (3): 45A
- ↑ Perle, A.; Norell, M. A.; Clark, J. M. (1999). «A new maniraptoran Theropod – Achillobator giganticus (Dromaeosauridae) – from the Upper Cretaceous of Burkhant, Mongolia». Contribution No. 101 of the Mongolian-American Paleontological Project: 1–105
- ↑ Norell, M.A.; P.J. Makovicky (1997). «Important features of the dromaeosaur skeleton: information from a new specimen». American Museum Novitates (3215): 1–45
- ↑ Norell, M.A.; Clark, James Matthew; Turner, Alan Hamilton; Makovicky, Peter J.; Barsbold, Rinchin; Rowe, Timothy (2006). «A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia)». American Museum Novitates (3545): 1–51. doi:10.1206/0003-0082(2006)3545[1:ANDTFU]2.0.CO;2
- ↑ Turner, A.H.; P.J. Makovicky; M.A. Norell (2007). «Feather quill knobs in the dinosaur Velociraptor» (PDF). Science. 317 (5845). 1721 páginas. Bibcode:2007Sci...317.1721T. PMID 17885130. doi:10.1126/science.1145076
- ↑ Brusatte, Stephen L.; Lloyd, Graeme T.; Wang, Steve C.; Norell, Mark A. (2014). «Gradual Assembly of Avian Body Plan Culminated in Rapid Rates of Evolution across the Dinosaur-Bird Transition» (PDF). Current Biology. 24 (20): 2386–2392. Bibcode:2014CBio...24.2386B. PMID 25264248. doi:10.1016/j.cub.2014.08.034
- ↑ Pei, Rui; Pittman, Michael; Goloboff, Pablo A.; Dececchi, T. Alexander; Habib, Michael B.; Kaye, Thomas G.; Larsson, Hans C.E.; Norell, Mark A.; Brusatte, Stephen L.; Xu, Xing (2020). «Potential for Powered Flight Neared by Most Close Avialan Relatives, but Few Crossed Its Thresholds» (PDF). Current Biology. 30 (20): 4033–4046.e8. Bibcode:2020CBio...30E4033P. PMID 32763170. doi:10.1016/j.cub.2020.06.105
- ↑ Powers, Mark J.; Fabbri, Matteo; Doschak, Michael R.; Bhullar, Bhart-Anjan S.; Evans, David C.; Norell, Mark A.; Currie, Philip J. (2021). «A new hypothesis of eudromaeosaurian evolution: CT scans assist in testing and constructing morphological characters». Journal of Vertebrate Paleontology. 41 (5). Bibcode:2021JVPal..41E0087P. doi:10.1080/02724634.2021.2010087 Verifique o valor de
|url-access=subscription(ajuda) - ↑ a b Czepiński, Łukasz (2023). «Skull of a dromaeosaurid Shri devi from the Upper Cretaceous of the Gobi Desert suggests convergence to the North American forms». Acta Palaeontologica Polonica. 68. doi:10.4202/app.01065.2023

- ↑ Cau, Andrea (2024). «A Unified Framework for Predatory Dinosaur Macroevolution» (PDF). Bollettino della Società Paleontologica Italiana. 63 (1): 1–19. ISSN 0375-7633. doi:10.4435/BSPI.2024.08 (inativo 6 July 2025) Verifique data em:
|doi-incorrecto=(ajuda) - ↑ Moutrille, Léa; Cau, Andrea; Chinzorig, Tsogtbaatar; Escuillié, François; Tsogtbaatar, Khishigjav; Ganzorig, Bayasgaa; Mallet, Christophe; Godefroit, Pascal (13 de julho de 2025). «A new bird-like dinosaur from the Upper Cretaceous of Mongolia with extremely robust hands supports niche partitioning among velociraptorines». Historical Biology (em inglês): 1–32. ISSN 0891-2963. doi:10.1080/08912963.2025.2530148
- ↑ a b Ruebenstahl, Alexander; Napoli, James; Turner, Alan; Bhullar, Bhart-Anjan; Norell, Mark A. (2021). «Two New Eudromaeosaurs from Khulsan (Central Mongolia) Reveal Modern-Like Faunal Predatory Structure Amoung [sic] Non-Avian Dinosaurs». Geological Society of America. Geological Society of America Abstracts with Programs. doi:10.1130/abs/2021AM-368751 Verifique o valor de
|url-access=subscription(ajuda) - ↑ Jerzykiewicz, Tomasz; Currie, Philip J.; Fanti, Federico; Lefeld, Jerzy (2021). «Lithobiotopes of the Nemegt Gobi Basin1». Canadian Journal of Earth Sciences. 58 (9): 829–851. Bibcode:2021CaJES..58..829J. doi:10.1139/cjes-2020-0148 Verifique o valor de
|url-access=subscription(ajuda) - ↑ Eberth, D. A. (2018). «Stratigraphy and paleoenvironmental evolution of the dinosaur-rich Baruungoyot-Nemegt succession (Upper Cretaceous), Nemegt Basin, southern Mongolia». Palaeogeography, Palaeoclimatology, Palaeoecology. 494: 29–50. Bibcode:2018PPP...494...29E. doi:10.1016/j.palaeo.2017.11.018 Verifique o valor de
|url-access=subscription(ajuda) - ↑ a b c Saneyoshi, Mototaka; Watabe, Mahito; Suzuki, Shigeru; Tsogtbaatar, Khishigjav (2011). «Trace fossils on dinosaur bones from Upper Cretaceous eolian deposits in Mongolia: Taphonomic interpretation of paleoecosystems in ancient desert environments». Palaeogeography, Palaeoclimatology, Palaeoecology. 311 cissue=1–2: 38–47. Bibcode:2011PPP...311...38S. doi:10.1016/j.palaeo.2011.07.024 Verifique o valor de
|url-access=subscription(ajuda) - ↑ a b Erro de citação: Etiqueta
<ref>inválida; não foi fornecido texto para as "refs" nomeadasEnnerth2018 - ↑ Erro de citação: Etiqueta
<ref>inválida; não foi fornecido texto para as "refs" nomeadasnemegtbgage - ↑ Novacek, Michael J.; Hoffman, Eva A.; o'Leary, Maureen A. (2022). «First occurrence of the eutherian mammal Asioryctes nemegtensis from the Upper Cretaceous Djadokhta Formation, Gobi Desert, Mongolia, and a revised alpha taxonomy based on the skull and dentition». Journal of Vertebrate Paleontology. 42 (4). Bibcode:2022JVPal..42E6320N. doi:10.1080/02724634.2023.2196320
- ↑ Arbour, V. M.; Currie, P. J.; Badamgarav, D. (2014). «The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia». Zoological Journal of the Linnean Society. 172 (3): 631−652. doi:10.1111/zoj.12185 Verifique o valor de
|url-access=subscription(ajuda) - ↑ Wilson, J. A. (2005). «Redescription of the Mongolian Sauropod Nemegtosaurus mongoliensis Nowinski (Dinosauria: Saurischia) and Comments on Late Cretaceous Sauropod Diversity». Journal of Systematic Palaeontology. 3 (3): 283−318. Bibcode:2005JSPal...3..283W. doi:10.1017/S1477201905001628
- ↑ Czepiński, Ł. (2019). «Ontogeny and variation of a protoceratopsid dinosaur Bagaceratops rozhdestvenskyi from the Late Cretaceous of the Gobi Desert». Historical Biology. 32 (10): 1394–1421. doi:10.1080/08912963.2019.1593404
- ↑ Alexander O. Averianov; Alexey V. Lopatin (19 de fevereiro de 2022). «A new alvarezsaurid theropod dinosaur from the Upper Cretaceous of Gobi Desert, Mongolia». Cretaceous Research. 135: 105168. Bibcode:2022CrRes.13505168A. doi:10.1016/j.cretres.2022.105168 Verifique o valor de
|url-access=subscription(ajuda) - ↑ Funston, G. F.; Mendonca, S. E.; Currie, P. J.; Barsbold, R. (1 de abril de 2018). «Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin». Palaeogeography, Palaeoclimatology, Palaeoecology. THE LATE CRETACEOUS NEMEGT ECOSYSTEM: DIVERSITY, ECOLOGY, AND GEOLOGICAL SIGNATURE (em inglês). 494: 101–120. Bibcode:2018PPP...494..101F. ISSN 0031-0182. doi:10.1016/j.palaeo.2017.10.023 Verifique o valor de
|url-access=subscription(ajuda) - ↑ Lee, S.; Lee, Y.-N.; Currie, P. J.; Sissons, R.; Park, J.-Y.; Kim, S.-H.; Barsbold, R.; Tsogtbaatar, K. (2022). «A non-avian dinosaur with a streamlined body exhibits potential adaptations for swimming». Communications Biology. 5 (1185): 1185. ISSN 2399-3642. PMC 9715538. PMID 36456823. doi:10.1038/s42003-022-04119-9
- ↑ Bell, Alyssa K.; Chiappe, Luis M.; Erickson, Gregory M.; Suzuki, Shigeru; Watabe, Mahito; Barsbold, Rinchen; Tsogtbaatar, K. (2010). «Description and ecologic analysis of Hollanda luceria, a Late Cretaceous bird from the Gobi Desert (Mongolia)». Cretaceous Research. 31 (1): 16–26. Bibcode:2010CrRes..31...16B. doi:10.1016/j.cretres.2009.09.001 Verifique o valor de
|url-access=subscription(ajuda) - ↑ Varricchio, David J.; Balanoff, Amy M.; Norell, Mark A. (2015). «Reidentification of Avian Embryonic Remains from the Cretaceous of Mongolia». PLOS ONE. 10 (6): e0128458. Bibcode:2015PLoSO..1028458V. PMC 4452496. PMID 26030147. doi:10.1371/journal.pone.0128458
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||








.png)


.jpg)




