Halszkaraptorinae
Halszkaraptorinae
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Cretáceo Superior, 75–71 Ma
Possível registro do Cretáceo Inferior[1] | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Género-tipo | |||||||||||||||||
| †Halszkaraptor escuilliei Cau et al., 2017 | |||||||||||||||||
| Gêneros | |||||||||||||||||
| |||||||||||||||||
| Sinónimos | |||||||||||||||||
| |||||||||||||||||
Halszkaraptorinae[2] é uma subfamília extinta de Dromaeosauridae basal (ou possivelmente Unenlagiidae), que inclui os gêneros Halszkaraptor, Natovenator [en], Mahakala [en] e Hulsanpes. Os membros de Halszkaraptorinae são conhecidos definitivamente apenas de estratos do Cretáceo Superior na Ásia, especificamente na Mongólia. Há debates entre pesquisadores sobre se o grupo tinha um estilo de vida semiaquático.
História da descoberta

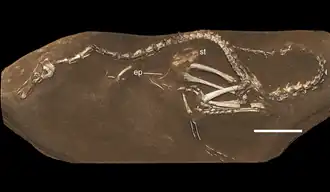
Os primeiros restos conhecidos de membros de Halszkaraptorinae foram encontrados em sedimentos de arenito em 1970, durante uma expedição polonesa-mongol na formação Barun Goyot, no deserto de Gobi. Em 1982, esses restos foram descritos pela paleontóloga polonesa Halszka Osmólska e usados como holótipo para o novo gênero e espécie Hulsanpes perlei, em homenagem ao paleontólogo mongol Altangerel Perle [en]. Embora as afinidades desses fósseis não fossem totalmente compreendidas, eles foram interpretados provisoriamente como pertencentes a Deinonychosauria.[2] Em 1992, fósseis semelhantes aos de Dromaeosauridae foram recuperados da localidade de Tugriken Shire, na formação Djadokhta, e descritos como uma nova espécie em 2007 por Alan H. Turner e equipe. Esse novo táxon, Mahakala omnogovae, foi considerado um membro de romaeosauridaep rimitivo, principalmente com base em suas proporções corporais.[3] Em 2015, o Instituto Real Belga de Ciências Naturais recebeu um pequeno fóssil de terópode, proveniente de coleções particulares da Inglaterra e do Japão. Após negociações entre o instituto, a Eldonia e as autoridades mongóis, o espécime foi devolvido ao Instituto de Paleontologia e Geologia da Academia de Ciências da Mongólia [en]. Segundo documentos associados, o espécime foi extraído ilegalmente da formação Djadokhta (localidade de Ukhaa Tolgod; no entanto, os sedimentos sugerem que, alternativamente, pode ter sido recuperado da localidade próxima de Bayn Dzak) em um ano desconhecido, acabando em coleções privadas. O espécime foi formalmente descrito em 2017 por Andrea Cau e colegas, que nomearam o gênero e a espécie Halszkaraptor escuilliei, em homenagem a Halszka Osmólska e François Escuillié, paleontólogos que viabilizaram as negociações para a devolução do espécime saqueado à Mongólia. Com a nomeação de Halszkaraptor, a subfamília Halszkaraptorinae também foi criada para incluir este táxon e seus parentes, nomeadamente Hulsanpes e Mahakala.[4]
Descrição

Os membros de Halszkaraptorinae eram terópodes relativamente pequenos, com comprimentos semelhantes aos de patos modernos.[3][4] Eles são caracterizados por pescoços longos, vértebras caudais [en] proximais com processos articulares orientados e lâminas zigapofisárias proeminentes (placas ósseas que formam as paredes posteriores de cada vértebra), uma ulna achatada com uma margem posterior afiada, ílio com um processo supratrocantérico em forma de prateleira (acima do trocanter do fêmur), haste do metacarpo III tão espessa transversalmente quanto a do metacarpo II, uma superfície posterodistal na haste do fêmur com uma fossa alongada delimitada por uma crista lateral e a metade proximal do metatarso III não constrita e marcadamente convexa anteriormente.[4]
Classificação
Halszkaraptorinae é definida como o clado mais inclusivo que contém Halszkaraptor escuilliei, mas não Dromaeosaurus albertensis, Unenlagia comahuensis, Saurornithoides mongoliensis ou condor-dos-andes.[4] O cladograma abaixo é baseado na análise filogenética conduzida em 2022 por Lee et al. na descrição de Natovenator [en]:[5]
| Dromaeosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||


Em 2025, Motta et al. recuperaram Halszkaraptorinae (elevada ao nível de família) junto com Unenlagiidae, Anchiornithidae [en] e Archaeopteryx como pertencentes ao clado Avialae.[6] As duas árvores recuperadas são reproduzidas abaixo:
- Consenso Melhorado (ponderação igual)
| Pennaraptora |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- Consenso Melhorado (ponderação implícita)
| Pennaraptora |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia

Em 2017, uma comparação dos fósseis de Halszkaraptor com os ossos de crocodilianos atuais e aves aquáticas revelou evidências de um estilo de vida semiaquático. Cau e colegas sugeriram que os membros de Halszkaraptorinae eram bípedes obrigatórios em terra, mas também nadadores que usavam seus membros anteriores para se impulsionar na água e seus pescoços longos para forragear. Tais características permitiam que explorassem paleoecossistemas terrestres e aquáticos.[4] Um estudo de 2024 por Tse, Miller e Pittman, focado na morfologia craniana e nas forças de mordida de vários membros de Dromaeosauridae, descobriu que Halszkaraptor tinha uma mordida rápida inadequada para alimentação piscívora, como previamente hipotetizado com base na morfologia do crânio, e sugeriu que era um insetívoro que caçava pequenos invertebrados, possivelmente em condições de pouca luz (à noite ou em águas turvas), já que provavelmente tinha uma visão excepcional em baixa luminosidade entre os membros de Dromaeosauridae, com base no tamanho relativamente grande de sua órbita.[7]
Outros pesquisadores discordaram ou apenas seguiram a interpretação de Cau de um dinossauro semiaquático. Em 2019, Brownstein argumentou que as características notadas para Halszkaraptor não suportam diretamente sua capacidade de nadar. Ele também sugeriu que este dinossauro pode ser um membro de Dromaeosauridae basal com características transicionais,[8] embora Cau tenha refutado suas alegações um ano depois.[9] Em 2021, Hone e Holtz observaram que, como Halszkaraptor e muitas aves aquáticas modernas sem unhas achatadas são considerados semiaquáticos, ter unhas achatadas como Spinosaurus não sugere necessariamente que o animal pudesse nadar; eles não propuseram sua própria visão sobre a potencial capacidade de nado deste dinossauro.[10] Em 2022, Fabbri e seus colegas argumentaram contra uma ecologia semiaquática para Halszkaraptor, observando que ele tinha baixa densidade óssea, uma característica não observada em animais semiaquáticos.[11] Em resposta, Cau apontou em seu blog que os cisnes também têm baixa densidade óssea, mas possuem adaptações para se alimentarem em ambientes semiaquáticos.[12]
Referências
- ↑ Kubota, K.; Kobayashi, Y.; Ikeda, T. (2024). «Early Cretaceous troodontine troodontid (Dinosauria: Theropoda) from the Ohyamashimo Formation of Japan reveals the early evolution of Troodontinae». Scientific Reports. 14 (1). 16392. Bibcode:2024NatSR..1416392K. PMC 11272788
 . PMID 39054320. doi:10.1038/s41598-024-66815-2
. PMID 39054320. doi:10.1038/s41598-024-66815-2
- ↑ a b Osmólska, H. (1982). «Hulsanpes perlei n.g. n.sp. (Deinonychosauria, Saurischia, Dinosauria) from the Upper Cretaceous Barun Goyot Formation of Mongolia». Neues Jahrbuch für Geologie und Paläontologie, Monatshefte. 1982 (7): 440–448. doi:10.1127/njgpm/1982/1982/440
- ↑ a b Turner, A.H.; Pol, D.; Clarke, J.A.; Erickson, G.M.; Norell, M.A. (2007). «A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight». Science. 317 (5843): 1378–1381. Bibcode:2007Sci...317.1378T. PMID 17823350. doi:10.1126/science.1144066
- ↑ a b c d e Cau, A.; Beyrand, V.; Voeten, D. F. A. E.; Fernandez, V.; Tafforeau, P.; Stein, K.; Barsbold, R.; Tsogtbaatar, K.; Currie, P. J.; Godefroit, P. (2017). «Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs». Nature. 552 (7685): 395–399. Bibcode:2017Natur.552..395C. PMID 29211712. doi:10.1038/nature24679 Supplementary Information
- ↑ Lee, S.; Lee, Y.-N.; Currie, P. J.; Sissons, R.; Park, J.-Y.; Kim, S.-H.; Barsbold, R.; Tsogtbaatar, K. (2022). «A non-avian dinosaur with a streamlined body exhibits potential adaptations for swimming». Communications Biology. 5 (1185): 1185. ISSN 2399-3642. PMC 9715538. PMID 36456823. doi:10.1038/s42003-022-04119-9
- ↑ Motta, Matías J.; Agnolína, Federico L.; Eglia, Federico Brissón; Rozadillaa, Sebastián; Novas, Fernando E. (22 de agosto de 2025). «Phylogenetic relationships of Unenlagiidae among Paraves (Dinosauria)». Journal of Systematic Palaeontology. 23 (1). doi:10.1080/14772019.2025.2529608. Consultado em 22 de agosto de 2025
- ↑ Tse, Y. T.; Miller, C. V.; Pittman, M. (2024). «Morphological disparity and structural performance of the dromaeosaurid skull informs ecology and evolutionary history». BMC Ecology and Evolution. 24 (1). 39. Bibcode:2024BMCEE..24...39T. PMC 11020771. PMID 38622512. doi:10.1186/s12862-024-02222-5
- ↑ Brownstein, C.D. (2019). «Halszkaraptor escuilliei and the evolution of the paravian bauplan». Nature. 9 (1): 16455. Bibcode:2019NatSR...916455B. PMC 6848195. PMID 31712644. doi:10.1038/s41598-019-52867-2
- ↑ Cau, A. (2020). «The body plan of Halszkaraptor escuilliei (Dinosauria, Theropoda) is not a transitional form along the evolution of dromaeosaurid hypercarnivory». PeerJ. 8. PMC 7047864. PMID 32140312. doi:10.7717/peerj.8672
- ↑ Hone; Holtz, Thomas R. Jr. (2021). «Evaluating the ecology of Spinosaurus: Shoreline generalist or aquatic pursuit specialist?». Palaeontologia Electronica. 24 (1): a03. doi:10.26879/1110. hdl:1903/28570
- ↑ Fabbri, Matteo; Navalón, Guillermo; Benson, Roger B. J.; Pol, Diego; O'Connor, Jingmai; Bhullar, Bhart-Anjan S.; Erickson, Gregory M.; Norell, Mark A.; Orkney, Andrew; Lamanna, Matthew C.; Zouhri, Samir; Becker, Justine; Emke, Amanda; Dal Sasso, Cristiano; Bindellini, Gabriele; Maganuco, Simone; Auditore, Marco; Ibrahim, Nizar (23 de março de 2022). «Subaqueous foraging among carnivorous dinosaurs». Nature. 603 (7903): 852–857. Bibcode:2022Natur.603..852F. PMID 35322229. doi:10.1038/s41586-022-04528-0
- ↑ Cau, Andrea (26 de março de 2022). «Theropoda: L'impatto della densità ossea di Halszkaraptor sulla sua ecologia». Consultado em 28 de março de 2023