Brancasaurus
Brancasaurus
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Berriasiano | |||||||||||||||||
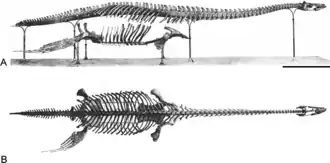 Espécime holótipo | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Nome binomial | |||||||||||||||||
| †Brancasaurus brancai Wegner, 1914 | |||||||||||||||||
| Distribuição geográfica | |||||||||||||||||
 | |||||||||||||||||
| Sinónimos | |||||||||||||||||
| |||||||||||||||||
Brancasaurus (que significa "lagarto de Branca") é um gênero de plesiossauro que viveu num lago de água doce no Cretáceo Inferior do que é hoje a Renânia do Norte-Vestfália, na Alemanha. Com um pescoço longo, possuindo vértebras com espinhas neurais distintamente em forma de "barbatana de tubarão", e uma cabeça relativamente pequena e pontiaguda, Brancasaurus é superficialmente semelhante a Elasmosaurus, embora menor em tamanho, com 3,26 m de comprimento como subadulto.
A espécie-tipo deste gênero é Brancasaurus brancai, nomeada pela primeira vez por Theodor Wegner em 1914 [en] em homenagem ao paleontólogo alemão Wilhelm von Branca [en].[1] Outro plesiossauro nomeado da mesma região, Gronausaurus wegneri, muito provavelmente representa um sinônimo deste gênero. Embora tradicionalmente considerado como um membro basal de Elasmosauridae, Brancasaurus foi mais recentemente recuperado como um membro, ou parente próximo, de Leptocleididae, um grupo que contém muitos outros plesiossauros de água doce.
Descoberta e nomeação
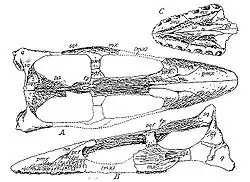
O espécime holótipo de Brancasaurus brancai é o GPMM A3.B4, armazenado na Universidade de Münster. Ele provém de uma mina de argila perto da cidade de Gronau, na Alemanha. O espécime foi descoberto em julho de 1910 por trabalhadores na mina de argila, que o escavaram com picaretas; ao fazerem isso, danificaram o espécime (em particular, o púbis foi quebrado em 176 pedaços) e deixaram para trás vários pequenos fragmentos que foram posteriormente coletados pessoalmente pelo paleontólogo Theodor Wegner, que em 1914 descreveu o espécime em detalhe. O esqueleto está razoavelmente completo, consistindo de várias partes do crânio, a maioria das vértebras, várias costelas isoladas e gastralia, partes das cinturas peitoral e pélvica, ambos os úmeros, um fêmur e vários ossos dos pés das nadadeiras. Com o tempo, várias partes foram perdidas, incluindo várias peças do crânio, dentes, gastralia e vértebras caudais, um segundo fêmur e um rádio, tíbia e fíbula. Um molde de cera do endocrânio do espécime-tipo está armazenado como SMF R4076 no Museu de História Natural Senckenberg.[2]
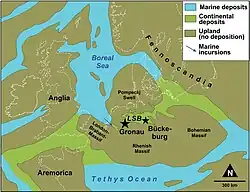
A mina de argila de onde provém o espécime-tipo faz parte da formação Isterberg no Grupo Bückeberg [en],[3] também conhecido no passado como a "fácies Wealden alemã".[4] O Grupo Bückeberg, que é dividido em seis zonas,[5] pertence ao Berriasiano do Cretáceo, com o limite entre o Berriasiano e o Valanginiano no topo do grupo.[6] As partes da formação Isterberg expostas em Gronau pertencem às zonas "Wealden 5" e "Wealden 6", que correspondem ao Berriasiano mais superior. Um segundo indivíduo subadulto, mais fragmentário, GZG.BA.0079, consiste no púbis, ísquio e vários componentes vertebrais; provém da formação Deister, ligeiramente mais baixa ("Wealden 3"[5]), no Grupo Bückeberg, e só pode ser atribuído a Brancasaurus sp., pois é relativamente incompleto e difere em várias características vertebrais menores do tipo de Brancasaurus brancai. Outros elementos prováveis, mas isolados, de Brancasaurus provêm de afloramentos das formações Isterberg e Fuhse na Baixa Saxônia; esta última formação também está no Grupo Bückeberg.[2]
Sinônimos

O espécime GPMM A3.B2 consiste em dentes, partes das mandíbulas, a caixa craniana e outras partes fragmentárias do crânio, vértebras, pedaços de costelas, parte da cintura peitoral, a cintura pélvica inteira, um úmero completo e um parcial, uma ulna, dois fêmures, uma fíbula e vários ossos dos pés. Embora este espécime tenha sido originalmente atribuído a Brancasaurus, Hampe (2013) o atribuiu a um novo gênero e espécie, Gronausaurus wegneri.[7] Foi descoberto cerca de 8 m mais alto na coluna estratigráfica do que o espécime-tipo de Brancasaurus. Análises posteriores descobriram que este espécime, que era maduro, era virtualmente indistinguível do tipo de Brancasaurus, com exceção do comprimento do ísquio, da altura das espinhas neurais cervicais, da largura dos centros cervicais e do fato de as espinhas neurais dorsais serem ou não constritas na base. Essas pequenas diferenças podem provavelmente ser atribuídas a variações individuais ou baseadas na idade, apoiando Gronausaurus wegneri como um sinônimo júnior de Brancasaurus brancai.[2]
E. Koken nomeou Plesiosaurus limnophilus em 1887 com base em vértebras cervicais isoladas de afloramentos do Grupo Bückeberg na Baixa Saxônia. Da mesma localidade, Koken posteriormente nomeou duas outras espécies de Plesiosaurus, Plesiosaurus degenhardti e Plesiosaurus kanzleri, e também atribuiu algum material a Plesiosaurus valdensis. Todo esse material não é particularmente diagnóstico e foi parcialmente perdido; assim, foram considerados nomina dubia. Sachs et al. consideraram todos esses como sendo restos de Brancasaurus, com exceção de Plesiosaurus degenhardti, que foi mantido como um nomen dubium por não possuir as espinhas neurais cervicais distintas de Brancasaurus.[2]
Descrição
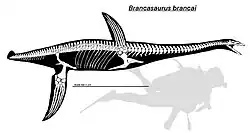
Brancasaurus era um plesiossauro de tamanho médio, com o espécime holótipo medindo 3,26 m de comprimento; este espécime provavelmente pertence a um subadulto, a julgar pelas suturas não fundidas nas vértebras, bem como pelo desenvolvimento de processos nos membros e no púbis.[2]
Crânio

O crânio do holótipo, que mede 23,7 cm de comprimento, é longo e estreito, com um focinho afilado que se inclina para baixo num ângulo de 15°. As órbitas oculares tinham aproximadamente o mesmo tamanho das aberturas temporais imediatamente atrás delas. Uma crista estreita e arredondada ao longo do meio da superfície superior do crânio estende-se desde perto da frente da pré-maxila até a parte de trás das órbitas oculares. Os ossos frontais formam uma barra retangular que separa as órbitas oculares pelo meio. Uma crista que atravessa a barra cruza com a crista que se estende para a frente para produzir uma protuberância em forma de adaga. O osso jugal, que se estende da parte inferior da órbita ocular até o nível das aberturas temporais, é inteiramente delimitado na sua parte inferior pela maxila. Os ossos esquamosais arqueiam-se para formar a parte de trás curvada do crânio e possuem uma crista no topo para a fixação dos músculos do pescoço. Há também uma crista no ponto onde os dois ossos se fundem. Um molde da caixa craniana mostra impressões dos canais semicirculares e do ouvido interno membranoso, bem como dos canais dos nervos hipoglosso, acessório, glossofaríngeo e vago, que também podem ser observados no exoccipital-opistótico ósseo da caixa craniana. Na mandíbula, imperfeitamente preservada, a eminência coronoide [en] parece ser relativamente baixa, a julgar pela borda superior estreita e ligeiramente curvada do osso surangular. Embora os dentes tenham sido perdidos, foram inicialmente descritos como longos, delgados e em forma de sovela, com cristas ásperas nas superfícies externas. Embora tenha sido sugerido que Brancasaurus tinha alvéolos dentários muito reduzidos na pré-maxila, como em Leptocleidus [en],[8] isso é impossível de verificar devido aos danos nesta porção do crânio.[2]
Coluna vertebral

O pescoço inteiro possui 37 vértebras cervicais e tem aproximadamente 1,18 m de comprimento. Os centros das vértebras são mais largos do que altos ou longos. Ambas as extremidades de cada vértebra são ligeiramente côncavas, o que significa que as vértebras são anficélicas. Os lados das vértebras são igualmente fracamente côncavos; ao contrário de muitos outros plesiossauros de pescoço longo, eles não possuíam uma crista na lateral (embora isso possa ser afetado pela idade). As espinhas neurais das vértebras têm uma forma distintiva de barbatana de tubarão, sendo altas e triangulares. Existem três vértebras peitorais na transição pescoço-corpo, que são fracamente côncavas, mais altas do que longas, e têm espinhas neurais de formato retangular que são direcionadas ligeiramente para trás. As vértebras cervicais e peitorais têm reentrâncias profundas através das quais a corda dorsal passava.[2]
As 19 vértebras dorsais são semelhantes às vértebras peitorais, sendo fracamente côncavas e mais altas do que longas, mas as espinhas neurais são proporcionalmente mais altas que os centros. As costelas dorsais de cabeça única são arredondadas, mas ligeiramente achatadas em secção transversal, e algumas têm uma projeção em forma de dente na extremidade superior; suas superfícies articulares são ligeiramente côncavas. Por baixo, há pelo menos dez pares de gastralia, cada um dos quais se afunila para os lados e tem um sulco central na superfície inferior. As três vértebras sacrais são semelhantes, mas têm costelas muito menores, mais rombas e de formato mais oval. A primeira costela sacral, comparativamente menor, é direcionada mais para fora e para trás do que as outras duas costelas. Inicialmente, havia 25 vértebras caudais preservadas, das quais 22 ainda estão presentes. As últimas vértebras caudais estão parcialmente fundidas numa estrutura semelhante a um pigóstilo. As costelas caudais preservadas são achatadas, triangulares e afunilam-se em direção à ponta da cauda.[2]
Membros e cinturas
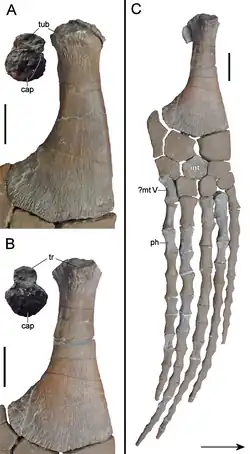

A interclavícula é uma grande placa com uma superfície superior lisa e um sulco proeminente na superfície inferior. Também possui uma pequena projeção pontiaguda na sua extremidade posterior. As escápulas têm prateleiras proeminentes de cada lado (diagnósticas de leptocleidídeos e policotilídeos, mas não fortemente diferenciadas em elasmossaurídeos), e seus glenoides são claramente côncavos, com fixações ásperas para cartilagem. Os dois coracoides curvam-se para fora no meio e entram em contato nas extremidades, formando um buraco no meio, embora a morfologia exata desse buraco seja incerta. As regiões onde os coracoides entram em contato são abobadadas e espessadas para formar uma projeção fraca em forma de crista, comparável, mas provavelmente adquirida convergentemente, aos elasmossaurídeos. Os púbis formam um prato um tanto retangular, com uma borda frontal convexa e uma borda externa côncava, enquanto os ísquios são planos, triangulares e em forma de placa. As bordas dos púbis, onde encontram os ísquios, curvam-se para dentro da linha média para cada lado. As bordas correspondentes dos ísquios têm formato semelhante, com as bordas curvas dos ossos formando coletivamente duas fenestras arredondadas que são conectadas no centro por uma pequena abertura em forma de losango, como também visto em Futabasaurus [en].[9] Os ílios são em forma de haste e dobrados, com projeções rombas a meio caminho de suas bordas externas; na extremidade superior, são achatados em forma de leque.[2]
Os úmeros, que têm um comprimento de cerca de 24 cm, são ovais em secção transversal e cerca de metade da largura em relação ao comprimento no ponto mais largo. Suas bordas dianteiras são curvas em forma de S, uma característica também vista em Leptocleidus [en], Hastanectes [en], policotilídeos e no elasmossaurídeo Wapuskanectes [en], mas não em Nichollssaura.[8][10] O único fêmur atualmente disponível tem 21,5 cm de comprimento; é côncavo em uma borda, enquanto a outra borda é reta perto do topo, mas curva-se acentuadamente perto da base. Os restantes ossos longos do membro foram perdidos. Alegadamente, o rádio era semelhante, mas menor e mais reto que a tíbia, e havia um buraco presente entre a tíbia e a fíbula. As 14 falanges preservadas, que provavelmente incluem elementos tanto dos membros anteriores quanto dos posteriores, são longas e em forma de ampulheta.[2]
Possível tecido mole
Aparentemente, tecido mole foi preservado com o espécime, mas foi subsequentemente removido durante a preparação. Cobrindo os membros e o resto do corpo havia uma camada de calcita lisa e multicamada, que foi originalmente interpretada como a preservação de pele em decomposição. Além disso, uma acumulação de sedimento na região abdominal pode ter representado o conteúdo intestinal, com gastrólitos e ossos digeridos. No entanto, como ambas as amostras do alegado tecido mole não estão mais disponíveis, é impossível verificar essas interpretações.[2]
Classificação
Inicialmente, Brancasaurus foi atribuído à Elasmosauridae por Wegner. Ele observou, no entanto, que tinha um pescoço mais curto e uma cabeça mais estreita, bem como várias morfologias distintas do teto do crânio, dentes e vértebras (especialmente as espinhas neurais em forma de "barbatana de tubarão" das vértebras cervicais) em comparação com outros membros do grupo conhecidos na época. Vários estudos subsequentes consideraram Brancasaurus como um membro basal de Elasmosauridae,[11][12][13][14] com alguns até usando Brancasaurus para definir o clado.[12] No entanto, várias opiniões taxonômicas contrárias foram expressas; em particular, Theodore E. White criou uma nova família, Brancasauridae, para conter Brancasaurus, Seeleyosaurus [en] e "Thaumatosaurus", um gênero extinto com espécies que agora pertencem a Rhomaleosaurus [en] e Meyerasaurus [en].[15][2]
Uma hipótese filogenética alternativa que ganhou tração substancial coloca Brancasaurus no clado Leptocleididae,[16][8][17] juntamente com outros leptocleidídeos, incluindo o próprio Leptocleidus [en], Vectocleidus [en], Umoonasaurus [en], Nichollssaura, e também possivelmente Hastanectes [en].[17] Este resultado foi recuperado pelas filogenias de Benson et al., que também notaram uma série de características morfológicas que aliam Brancasaurus ao clado mais geral Leptocleidia.[8][2]
Uma análise filogenética de 2016 conduzida por Sachs et al. encontrou duas colocações alternativas igualmente fortes para Brancasaurus (incluindo Gronausaurus): dentro de Leptocleididae; ou como o táxon irmão de um clado contendo tanto Leptocleididae quanto Polycotylidae, com o clado contendo todos os táxons mencionados sendo o táxon irmão de Elasmosauridae. O estudo concluiu que, atualmente, nenhum conjunto de dados filogenéticos é suficiente para resolver as relações de Brancasaurus. Além do fato de que o espécime-tipo é um subadulto, essa inconsistência nos resultados pode ser atribuída à mistura de características de leptocleidídeos, policotilídeos e elasmossaurídeos que é vista em Brancasaurus.[17] Os cladogramas abaixo ilustram os arranjos alternativos.[2]
Topologia A: ’'Brancasaurus'‘ na família Leptocleididae, com base em Benson et al. (2013)[8]
|
Topologia B: ’'Brancasaurus'‘ fora da família Leptocleididae, com base em Benson & Druckenmiller (2014)[17]
|
|
Paleoecologia
O Grupo Bückeberg, de onde provém Brancasaurus, provavelmente representava um grande lago continental de água doce para o qual as terras altas circundantes drenavam. Por sua vez, o próprio lago estava temporariamente conectado ao Mar Boreal através de uma passagem a oeste. Durante o tempo em que as camadas de "Wealden 5" e "Wealden 6" foram depositadas, o lago expandiu-se e tornou-se mais salobro como resultado da transgressão marinha.[18] Os sedimentos depositados provavelmente representam a porção inferior do lago, pobre em oxigênio, com os plesiossauros do Grupo Bückeberg sendo presumivelmente preservados após afundarem na coluna de água até o fundo.[2]
Além de Brancasaurus, outros constituintes do Grupo Bückeberg são invertebrados bentônicos, incluindo bivalves neomiodontídeos;[2] Hybodontiformes [en], incluindo Hybodus, Egertonodus [en], Lonchidion [en] e Lissodus [en]; os peixes actinopterígios Caturus [en], Lepidotes, Coelodus [en], Sphaerodus, Ionoscopus [en] e Callopterus [en],[7] dos quais Brancasaurus teria se alimentado em águas superficiais;[19] a tartaruga Desmemys;[7] crocodilianos, incluindo Goniopholis, Pholidosaurus e Theriosuchus; o terópode Altispinax; o marginocéfalo Stenopelix [en]; e um anquilossauro atribuído a Hylaeosaurus.[20][21] Outros restos indeterminados foram atribuídos a pterossauros; aos clados de crocodilianos Hylaeochampsidae [en] e Eusuchia; e aos clados de dinossauros Dryosauridae, Ankylopollexia, Troodontidae e Macronaria.[21]
Referências
- ↑ Wegner, T.H. 1914. "Brancasaurus brancai n. g. n. sp., ein Elasmosauride aus dem Wealden Westfalens". Festschrift für Wilhelm Branca zum 70. Geburtstage. Borntraeger; Leipzig: pp. 235–305
- ↑ a b c d e f g h i j k l m n o p Sachs, S.; Hornung, J.J.; Kear, B.P. (2016). «Reappraisal of Europe's most complete Early Cretaceous plesiosaurian: Brancasaurus brancai Wegner, 1914 from the "Wealden facies" of Germany». PeerJ. 4: e2813. PMC 5183163
 . PMID 28028478. doi:10.7717/peerj.2813
. PMID 28028478. doi:10.7717/peerj.2813
- ↑ Casey, R.; Allen, P.; Dörhöfer, G.; Gramann, F.; Hughes, N. F.; Kemper, E.; Rawson, P. F.; Surlyk, F. (1975). «Stratigraphical subdivision of the Jurassic-Cretaceous boundary beds in NW Germany». Newsletters on Stratigraphy. 4 (1): 4–5. doi:10.1127/nos/4/1975/4
- ↑ Allen, P. (1955). «Age of the Wealden in North-Western Europe». Geological Magazine. 92 (4): 265–281. Bibcode:1955GeoM...92..265A. doi:10.1017/S0016756800064311
- ↑ a b Elstner, F.; Mutterlose, J. (1996). «The Lower Cretaceous (Berriasian and Valanginian) in NW Germany». Cretaceous Research. 17 (1): 119–133. doi:10.1006/cres.1996.0010
- ↑ Mutterlose, J.; Bodin, S.; Fahnrich, L. (2014). «Strontium-isotope stratigraphy of the Early Cretaceous (Valanginian–Barremian): Implications for Boreal–Tethys correlation and paleoclimate». Cretaceous Research. 50 (4): 252–263. doi:10.1016/j.cretres.2014.03.027
- ↑ a b c Hampe, O. (2013). «The forgotten remains of a leptocleidid plesiosaur (Sauropterygia: Plesiosauroidea) from the Early Cretaceous of Gronau (Münsterland, Westphalia, Germany)». Paläontologische Zeitschrift. 78 (4): 473–491. doi:10.1007/s12542-013-0175-3
- ↑ a b c d e Benson, R.B.J.; Ketchum, H.F.; Naish, D.; Turner, L.E. (2013). «A new leptocleidid (Sauropterygia, Plesiosauria) from the Vectis Formation (Early Barremian–early Aptian; Early Cretaceous) of the Isle of Wight and the evolution of Leptocleididae, a controversial clade». Journal of Systematic Palaeontology. 11 (2): 233–250. doi:10.1080/14772019.2011.634444
- ↑ Sato, Tamaki; Hasegawa, Y.; Manabe, M. (2006). «A new elasmosaurid plesiosaur from the Upper Cretaceous of Fukushima, Japan». Palaeontology. 49 (3): 467–484. doi:10.1111/j.1475-4983.2006.00554.x
- ↑ Albright, L.B.; Gillette, D.D.; Titus, A.L. (2007). «Plesiosaurs from the Upper Cretaceous (Cenomanian–Turonian) Tropic Shale of Southern Utah, part 2: Polycotylidae» (PDF). Journal of Vertebrate Paleontology. 27 (1): 41–58. JSTOR 4524666. doi:10.1671/0272-4634(2007)27[41:pftucc]2.0.co;2
- ↑ Brown, D.S. (1981). «The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria». Bulletin of the British Museum. 35: 253–347
- ↑ a b O'Keefe, F.R. (2001). «A Cladistic Analysis and Taxonomic Revision of the Plesiosauria (Reptilia: Sauropterygia)». Acta Zoologica Fennica. 213: 1–63
- ↑ O'Keefe, F.R. (2004). «Preliminary description and phylogenetic position of a new plesiosaur (Reptilia: Sauropterygia) from the Toarcian of Holzmaden, Germany» (PDF). Journal of Paleontology. 78 (5): 973–988. doi:10.1666/0022-3360(2004)078<0973:PDAPPO>2.0.CO;2
- ↑ Großman, F. (2007). «The taxonomic and phylogenetic position of the Plesiosauroidea from the Lower Jurassic Posidonia Shale of south-west Germany». Palaeontology. 50 (3): 545–564. doi:10.1111/j.1475-4983.2007.00654.x
- ↑ White, T.E. (1940). «Holotype of Plesiosaurus longirostris Blake and Classification of the Plesiosaurs». Journal of Paleontology. 14 (5): 451–467. JSTOR 1298550
- ↑ Ketchum, H.F.; Benson, R.B.J. (2010). «Global interrelationships of Plesiosauria (Reptilia, Sauropterygia) and the pivotal role of taxon sampling in determining the outcome of phylogenetic analyses». Biological Reviews. 85 (2): 361–392. PMID 20002391. doi:10.1111/j.1469-185X.2009.00107.x
- ↑ a b c d Benson, R.B.J.; Druckenmiller, P.S. (2014). «Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition». Biological Reviews. 89 (1): 1–23. PMID 23581455. doi:10.1111/brv.12038
- ↑ Mutterlose, J.; Bornemann, A. (2000). «Distribution and facies patterns of Lower Cretaceous sediments in northern Germany: a review». Cretaceous Research. 21 (6): 733–759. doi:10.1006/cres.2000.0232
- ↑ Halstead, L.B. (1989). «Plesiosaur locomotion». Journal of the Geological Society. 146 (1): 37–40. Bibcode:1989JGSoc.146...37H. doi:10.1144/gsjgs.146.1.0037
- ↑ Sachs, S.; Hornung, J.J. (2013). «Ankylosaur Remains from the Early Cretaceous (Valanginian) of Northwestern Germany». PLOS ONE. 8 (4): e60571. Bibcode:2013PLoSO...860571S. PMC 3616133. PMID 23560099. doi:10.1371/journal.pone.0060571
- ↑ a b Hornung, J.J. (2013). Contributions to the Palaeobiology of the Archosaurs (Reptilia: Diapsida) from the Bückeberg Formation ('Northwest German Wealden'– Berriasian-Valanginian, Lower Cretaceous) of northern Germany (Dr. rer. nat.). Georg-August University School of Science. pp. 318–351. hdl:11858/00-1735-0000-0001-BACB-5
Ligações externas
- Brancasaurus no Diretório de Plesiossauros
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||






.png)