Aristonectes
Aristonectes
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Cretáceo Superior, Maastrichtiano, ~70–66 Ma | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Espécie-tipo | |||||||||||||||||||
| Aristonectes parvidens Cabrera, 1941 | |||||||||||||||||||
| Outras espécies | |||||||||||||||||||
| |||||||||||||||||||
| Sinónimos | |||||||||||||||||||
Lista
| |||||||||||||||||||

Aristonectes (que significa "melhor nadador") é um gênero extinto de grandes plesiossauros elasmossaurídeos que viveram durante o estágio Maastrichtiano do Cretáceo Superior. Duas espécies são conhecidas, Aristonectes parvidens e Aristonectes quiriquinensis, cujos restos fósseis foram descobertos no que são hoje a Patagônia e a Antártida. Ao longo do século XX, Aristonectes foi um animal difícil de analisar pelos cientistas devido à má preparação dos fósseis, e suas relações com outros gêneros eram incertas. Após revisões e descobertas subsequentes realizadas a partir do início do século XXI, Aristonectes é agora reconhecido como o gênero-tipo [en] da subfamília Aristonectinae, uma linhagem de elasmossaurídeos caracterizada por um crânio aumentado e um comprimento reduzido do pescoço.
Medindo mais de 10 m de comprimento, Aristonectes é um plesiossauro notavelmente imponente. Um espécime referido, descoberto na Antártida, tem um tamanho estimado de mais de 11 m de comprimento, o que tornaria este gênero um dos maiores plesiossauros conhecidos [en]. Os membros em forma de remo de Aristonectes são invulgarmente grandes para um elasmossaurídeo, atingindo quase 3 m de comprimento, sugerindo uma envergadura total de cerca de 7 m para o animal. O crânio tem formato ogival e é achatado, possuindo dentes afiados voltados para a frente. De acordo com as estimativas, haveria 13 dentes em cada pré-maxila, 50 dentes na maxila e de 50 a 63 ou mais dentes na mandíbula inferior.
De acordo com sua morfologia, principalmente craniana, Aristonectes se alimentava misturando presas e sedimentos nas zonas bentônicas, como a baleia-cinzenta moderna. Como em outros plesiossauros, gastrólitos (pedras estomacais) teriam sido usados por Aristonectes para ajudar a digerir sua comida ou para auxiliar na flutuabilidade, embora haja pouco apoio para a primeira hipótese. De acordo com sua distribuição geográfica no registro fóssil, Aristonectes teria migrado regularmente entre a Patagônia e a Antártida.
História da pesquisa
Aristonectes parvidens
O primeiro fóssil de Aristonectes foi descoberto muito antes de o gênero ser nomeado por Ángel Cabrera em 1941.[1] Em 1848, o naturalista franco-chileno Claude Gay descreveu o primeiro plesiossauro conhecido da América do Sul, Plesiosaurus [en] chilensis, com base em uma única vértebra caudal descoberta na ilha Quiriquina, na província de Concepción, Chile.[2] Em um livro publicado em 1889, Richard Lydekker colocou esta espécie no gênero Cimoliosaurus.[3] Em 1895, Wilhelm Deecke a moveu para o gênero de pliosaurídeo Pliosaurus como Pliosaurus chilensis, e atribuiu outros fósseis que foram descobertos na mesma localidade a esta espécie. Entre esses fósseis estão algumas vértebras, um fêmur, fragmentos de um ísquio e costelas.[4] Em 1918, o paleontólogo russo Pavel A. Pravoslavlev a moveu para o gênero relacionado Elasmosaurus, como Elasmosaurus chilensis.[5] Em 1941, Cabrera descreveu um espécime diferente da província de Chubut, Argentina, como o novo gênero e espécie Aristonectes parvidens. Na mesma publicação, ele listou Elasmosaurus chilensis como "Plesiosaurus" chilensis, expressando incerteza sobre suas afinidades.[6] Em 1949, Edwin H. Colbert descobriu que o espécime holótipo de "P." chilensis (a vértebra caudal originalmente descrita por Gay) pertence a um pliossauroide, mas também estava incerto sobre sua colocação genérica. Também no artigo, ele considera que o outro material fóssil anteriormente atribuído a "P." chilensis poderia vir de elasmossaurídeos.[7][8] Em uma análise publicada em 2013 por José P. O'Gorman e colegas, o espécime holótipo de "P." chilensis foi reconhecido como potencialmente pertencente a Aristonectes parvidens. Como este espécime consiste apenas em uma única vértebra caudal, foi referido como A. cf. parvidens para indicar incerteza em sua atribuição.[1]
O primeiro espécime formalmente identificado como Aristonectes parvidens foi coletado por Cristian S. Petersen em colaboração com o residente local Victor Saldivia, no Cañadón de los Loros, perto do município de Paso del Sapo [en], na província de Chubut, Argentina.[6] O espécime foi descoberto na formação Lefipán [en] e foi datado do Maastrichtiano.[9][10][11] Em setembro de 1940, Pablo Groeber enviou este espécime ao Museu de La Plata como uma doação para a Dirección de Minas y Geología del Ministerio de Agricultura. Uma vértebra e falanges incompletas da mesma região haviam sido doadas anteriormente ao Museu de La Plata por Mario Reguiló, e foram subsequentemente atribuídas ao holótipo, catalogado como MLP 40-XI-14-6, pois muito provavelmente provinham do mesmo espécime.[6] O holótipo consiste em um crânio parcial ligado à mandíbula, o atlas e o áxis, 21 vértebras cervicais (incluindo 16 cervicais anteriores articuladas), 8 vértebras caudais e um membro anterior [en] esquerdo incompleto. O espécime foi interpretado como um adulto, ou mesmo um indivíduo idoso.[9][12] Cabrera descreveu o animal como Aristonectes parvidens em seu artigo publicado em 1941 pelo museu que abriga o material fóssil.[6] O nome do gênero Aristonectes vem das palavras gregas antigas ἄριστος (áristos, "melhor", "superior") e νηκτός (nêktós, "nadador"), e pode ser traduzido como "melhor nadador", assim nomeado para indicar o espécime mais bem preservado de plesiossauro da Argentina conhecido na época. O epíteto específico parvidens significa "dentes pequenos", em referência ao tamanho bastante pequeno dos alvéolos dentários.[13]
Como as partes desconhecidas do espécime original foram fortemente reconstruídas, a anatomia precisa de Aristonectes permaneceu incerta ao longo do século XX.[14][15][16] Em 2003, a paleontóloga argentina Zulma Gasparini e colegas prepararam novamente o crânio holótipo, juntamente com o atlas e o áxis, na tentativa de esclarecer sua anatomia.[9] Em uma tese de doutorado publicada em 2013, O'Gorman atribui informalmente várias vértebras caudais a Aristonectes parvidens.[10] Em 2016, o mesmo autor revisou novamente o holótipo.[11]
Aristonectes quiriquinensis
A primeira descoberta da segunda espécie conhecida, Aristonectes quiriquinensis, data do final da década de 1950, quando o paleontólogo argentino Rodolfo Casamiquela [en] descobriu um esqueleto parcial na baía de Las Tablas, localizada ao norte da ilha Quiriquina, Chile. O espécime, catalogado como SGO.PV.260, foi encontrado em camadas maastrichtianas da formação Quiriquina [en]; inicialmente, cinco vértebras, um fragmento de úmero e a extremidade distal de um membro eram visíveis. A dureza das rochas que continham os fósseis dificultou a preparação subsequente. A preparação revelou partes adicionais do corpo, com o espécime incluindo todos os quatro membros, uma porção posterior do pescoço, a maior parte do tronco e uma cauda completa. É interpretado como um filhote. Este espécime foi analisado pela primeira vez em 2012 e identificado como um aristonectíneo indeterminado,[17] antes de ser atribuído ao gênero Aristonectes em 2013.[18]
Em 2001, o paleontólogo chileno Mario E. Suárez coletou um crânio parcial, fragmentos mandibulares e 12 vértebras cervicais anteriores de uma praia perto de Cocholgüe, localizada ao norte de Tomé. Este material foi descrito no ano seguinte e atribuído a Aristonectes, mas a identificação como uma espécie separada era então impossível devido à falta de preparação.[19] No início de 2009, uma segunda escavação foi realizada independentemente no mesmo local por uma equipe da Universidade de Concepción do Chile e da Universidade de Heidelberg da Alemanha. Esta escavação recuperou 119 blocos de arenito, a maioria com material ósseo, mas alguns foram degradados pela imersão periódica da água do mar, que causou superfícies quebradiças nas partes mais delicadas. Além disso, vários contatos anatômicos foram perdidos quando blocos de arenito foram cortados da praia com uma serra de rocha na maré baixa. A localização precisa do achado e a distribuição tafonômica dos fósseis mostram que eles pertencem ao mesmo espécime que foi descoberto em 2001. Os ossos foram enviados ao Museu Nacional de História Natural do Chile para preparação e análise científica. O fóssil completamente preparado consiste no crânio, no áxis e no atlas, 12 vértebras cervicais anteriores, 23 vértebras cervicais médias a posteriores, a maior parte do tronco, os dois membros anteriores quase completos e uma parte significativa do membro posterior direito. O espécime era provavelmente um jovem adulto e está catalogado como SGO.PV.957. Em 2014, Rodrigo Otero e colegas o tornaram o holótipo da nova espécie Aristonectes quiriquinensis; esses pesquisadores também atribuíram o espécime anterior, SGO.PV.260, a esta espécie. O epíteto específico é uma referência à formação de Quiriquina, que é a localidade-tipo deste táxon.[12]
Em 2015, outros dois espécimes da ilha Quiriquina foram atribuídos a Aristonectes quiriquinensis. Estes são compostos por um fêmur esquerdo completo e uma parte proximal de um úmero, catalogados respectivamente como SGO.PV.135 e SGO.PV.169, que foram anteriormente atribuídos ao gênero agora duvidoso Mauisaurus [en].[20] Em 2018, o espécime holótipo foi redescrito após preparação adicional. Na mesma publicação, outro espécime, SGO.PV.94, consistindo de uma série de vértebras caudais anteriores, também foi atribuído a Aristonectes quiriquinensis.[21]
Descrição
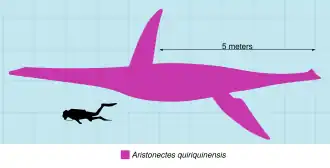
Aristonectes é um plesiossauro que foi particularmente difícil de analisar pelos cientistas devido à sua incompletude.[15][16] Suas relações permaneceram incertas até que pesquisas mais recentes realizadas em Aristonectes parvidens e a descoberta da segunda espécie, Aristonectes quiriquinensis, tornaram possível redescrever sua anatomia e classificá-lo entre os elasmossaurídeos.[9][11][12][21][14] No entanto, as comparações anatômicas entre Aristonectes parvidens e Aristonectes quiriquinensis ainda são limitadas aos elementos que são conhecidos de ambas as espécies.[12][11] Aristonectes é um grande representante dos plesiossauroides, com o holótipo de Aristonectes quiriquinensis tendo um tamanho estimado de cerca de 10 m ou mais de comprimento.[21] Um espécime referido de Aristonectes descoberto na formação López de Bertodano [en], na Antártida, catalogado como MLP 89-III-3-1, é considerado um dos maiores e mais pesados plesiossauros [en] identificados até hoje, com um tamanho estimado entre 11 e 11,9 m de comprimento e uma massa corporal de 10,7 a 13,5 toneladas.[22]
Crânio
Nas duas espécies conhecidas, o crânio de Aristonectes é achatado e em forma de ogiva.[9][11][12][21] O tamanho do crânio varia muito pouco entre as duas espécies, o do holótipo de Aristonectes quiriquinensis tendo um tamanho proposto entre 65-70 cm, enquanto o do holótipo de Aristonectes parvidens mede 73,5 cm.[12] Em Aristonectes quiriquinensis, os ossos esquamosais se estendem bem posteriormente ao côndilo occipital. Os ossos pterigoides são grandes e se projetam posteriormente ao neurocrânio. As extensões posteriores dos ossos pterigoides circundam o áxis e o atlas, o que reduz o espaço entre o crânio e as vértebras e, portanto, limitaria o movimento lateral em suas articulações.[21] Cerca de 13 dentes estavam presentes na pré-maxila e 50 na maxila.[12][11] Entre as características únicas do crânio de Aristonectes quiriquinensis está a presença de uma protuberância mentoniana (uma crista ao longo da sínfise mandibular), uma característica não observada no holótipo de Aristonectes parvidens.[12]
A sínfise mandibular de Aristonectes é curta.[6][9][11][12][21] Os ossos dentários, embora parcialmente preservados em ambas as espécies, teriam um total estimado de 50 dentes em Aristonectes quiriquinensis e 63 ou mais em Aristonectes parvidens.[9][12][23][11] A sínfise mandibular dos ossos dentários em Aristonectes quiriquinensis é comparativamente mais espessa e não possui o sulco profundo visto em vista ventral em Aristonectes parvidens. Os dentes mais posteriores de A. parvidens estavam cerca de 60 mm rostralmente ao processo coronoide [en]. Em Aristonectes quiriquinensis, não há alvéolos dentários presentes, embora 130 mm da mandíbula estejam preservados rostralmente ao processo coronoide. Portanto, é provável que A. quiriquinensis tivesse menos dentes que Aristonectes parvidens. Em A. parvidens, o contato labial entre o osso angular [en] e o surangular, anterior à fossa glenoide, apresenta uma cavidade profunda que não está presente em Aristonectes quiriquinensis.[12]
Os dentes de A. quiriquinensis têm uma secção transversal oval, sendo muito semelhantes aos de elasmossaurídeos do Hemisfério Norte e da província Weddelliana (uma área geográfica que surgiu após o isolamento da Antártida), como em Kaiwhekea [en]. A raiz dos dentes completos é ligeiramente mais longa que a coroa. A dentição de A. quiriquinensis, e muito provavelmente de Aristonectes parvidens, é homodonte, o que significa que os dentes são todos de formato semelhante. O maior dente conhecido é muito fino e pontiagudo, com cristas no lado lingual da coroa. O dente menor é um dente de substituição de raiz curta, com morfologia semelhante ao maior.[12] Todos os dentes são inclinados para a frente e não se encaixariam quando as mandíbulas estivessem fechadas.[21]
Esqueleto pós-craniano
Embora restos pós-cranianos sejam conhecidos em ambas as espécies, eles são mais completamente conhecidos e, portanto, mais bem documentados, em A. quiriquinensis.[12][21] Em 2018, Otero e seus colegas estimaram que Aristonectes quiriquinensis teria um total de 109 vértebras, incluindo 43 cervicais, 3 peitorais, 24 dorsais, 3 sacrais e 35 caudais.[21] As costelas cervicais [en] anteriores são recurvadas em Aristonectes parvidens e fundidas distalmente,[6] enquanto as de Aristonectes quiriquinensis são curtas e não possuem contatos.[12] As costelas cervicais e as espinhas neurais cervicais de Aristonectes são curvadas em direção à cabeça, semelhantes a outros membros da subfamília Aristonectinae.[12][21] Essa morfologia teria permitido manter o tronco mais alto que a cabeça.[21] As espinhas neurais presentes no meio da parte cervical se parecem com lâminas e são alargadas distalmente.[12]
As vértebras dorsais são maiores e mais altas que as vértebras cervicais e também são mais altas do que largas. Os centros das vértebras dorsais têm superfícies articulares anficélicas.[12] Entre as cavidades glenoides, a cintura peitoral de Aristonectes quiriquinensis tem quase um metro de largura. As costelas parecem espessadas em suas extremidades distais. As gastralia são paquiostóticas [en] e de comprimento moderado. As vértebras caudais de A. quiriquinensis são mais largas do que altas ou longas. As espinhas neurais das caudais tornam-se progressivamente rombas e curtas e têm um ápice dorsal espesso com uma superfície plana, o que sugere a provável existência de um forte ligamento ao longo da parte dorsal da cauda.[21]
Os membros de A. quiriquinensis são muito longos para um elasmossaurídeo, com a nadadeira mais bem preservada tendo cerca de 3 m de comprimento. Combinado com a largura mencionada da cintura peitoral, isso indica uma envergadura total de 7 m. Em tetrápodes aquáticos, tais proporções de membros só são observáveis no romaleossaurídeo Meyerasaurus [en] do Jurássico Inferior e na baleia-jubarte moderna.[21] Aristonectes parvidens também teria um tamanho de nadadeira semelhante. Em ambas as espécies, as falanges de seus membros têm grandes facetas articulares, apresentando ossos alongados em forma de bobina. Em Aristonectes quiriquinensis, a ulna é ligeiramente mais curta que o rádio e a tíbia é ligeiramente mais larga que a fíbula.[12]
Classificação

Aristonectes foi reclassificado várias vezes ao longo de sua história taxonômica.[12][11] Em 1941, Cabrera o classificou dentro da Elasmosauridae com base na forma dos elementos cranianos,[6] o que se provaria correto décadas depois.[24][25][21] Em 1962, Samuel P. Welles [en] considerou Aristonectes um pliossauro indeterminado com base em sua anatomia bastante enigmática.[14] Em 1960, Per O. Persson o classificou em Cimoliasauridae, uma proposta que ele reafirmou em 1963 com base em novas observações no crânio.[15] Em 1981, David S. Brown moveu Aristonectes para Cryptoclididae com base na presença de 6 a 15 dentes na pré-maxila.[16] A partir da primeira revisão do gênero realizada em 2003, Gasparini e seus colegas reclassificaram Aristonectes na família Elasmosauridae,[9] mas essa classificação foi rejeitada no mesmo ano por F. Robin O'Keefe e William Wahl Jr., que colocaram o gênero em Cimoliasauridae.[26] Em 2009, O'Keefe e Hallie P. Street revisaram a validade de Cimoliasauridae e descobriram que este táxon é um sinônimo júnior de Elasmosauridae. Portanto, eles moveram os gêneros Aristonectes, Kaiwhekea, Kimmerosaurus [en] e Tatenectes para a recém-erigida família Aristonectidae.[27] Em 2011, dois anos depois, O'Keefe e colegas estavam céticos sobre a classificação de Aristonectes e Kaiwhekea com Tatenectes, porque suas morfologias não pareciam corresponder.[28] Otero e colegas, em 2012, erigiram Aristonectinae como uma subfamília dentro da Elasmosauridae. Este grupo é caracterizado por um crânio muito aumentado em comparação com a largura do corpo, um pescoço moderadamente curto e mais de 25 dentes na maxila,[17] um número maior do que em outros elasmossaurídeos.[23] Juntamente com Elasmosaurinae, Aristonectinae representa uma das duas subfamílias reconhecidas dentro dos elasmossaurídeos.[29] Em muitas análises filogenéticas, Aristonectes é o gênero mais derivado dentro de Aristonectinae.[24][25][29][30]
O cladograma a seguir é modificado de Otero & Acuña, (2020):[30]
| Elasmosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia
Gastrólitos
Como na maioria dos outros plesiossauros, gastrólitos foram encontrados em alguns espécimes de Aristonectes. Entre estes estão 5 elementos descobertos durante a segunda exumação do holótipo de Aristonectes quiriquinensis,[12] e mais de 793 no maior espécime conhecido do gênero, MLP 89-III-3-1, da Antártida. No entanto, como este último espécime foi encontrado desarticulado (ou seja, os ossos se moveram de sua posição anatômica original), é provável que nem todos os seus gastrólitos tenham sido encontrados.[31] A função dos gastrólitos em plesiossauros ainda é controversa.[32] As duas hipóteses mais frequentemente citadas são seu uso no controle da flutuabilidade e seu uso como auxílio na digestão; esta última sendo a mais amplamente aceita em estudos recentes.[33][34][35] Em 2014, O'Gorman e colegas analisaram os gastrólitos do espécime da Antártida e concluíram que Aristonectes não selecionava pedras individuais para ingestão, mas engolia lotes de sedimento aleatoriamente que continham pedras de vários tamanhos. Isso é indicado pela falta de seleção de tamanho: a distribuição de tamanho dos gastrólitos é semelhante à esperada de uma amostra aleatória de sedimento.[31]
Alimentação
As nadadeiras e a cauda de Aristonectes quiriquinensis se assemelham às de vários cetáceos modernos, e os movimentos laterais da cabeça teriam sido limitados. Além disso, o crânio do animal tem uma boca aumentada que teria permitido o engolfamento de um grande volume de água. Isso indica que A. quiriquinensis se alimentava em zonas bentônicas, misturando presas e sedimentos ao mesmo tempo. Este tipo de padrão de alimentação também é documentado em baleias-cinzentas modernas. A presença de crustáceos decápodes e peixes na formação Quiriquina reforça ainda mais essa afirmação.[21]
Crescimento
Após a descoberta de A. quiriquinensis em 2014, vários espécimes foram subsequentemente atribuídos a esta espécie. Em um estudo publicado em 2015 por Otero e seus colegas, foi notado que, durante sua ontogenia, o úmero e o fêmur mudam de um capítulo [en] plano em juvenis para cabeças de formato hemisférico na idade adulta.[20]
Paleoecologia

Aristonectes é conhecido de várias formações geológicas da Patagônia e da Antártida que datam do Maastrichtiano. É conhecido das formações Quiriquina [en][12][21] e Dorotea [en] no Chile,[36] das formações Allen, Jagüel e Lefipán na Argentina[11] e da formação López de Bertodano na Antártida.[37][22] A presença de Aristonectes nessas formações mostra que ele teria sido endêmico da província Weddelliana, uma área geográfica que surgiu após o isolamento da Antártida.[36] Isso também indica que o gênero poderia ter migrado regularmente entre a Antártida e a Patagônia, como muitos cetáceos atuais.[21] Muitos peixes ósseos, peixes cartilaginosos, crustáceos e moluscos são conhecidos da maioria das localidades de onde Aristonectes é listado.[38][39][40][36][21][41]
A diversificação de plesiossauros parece variar de formação para formação. Na formação Lefipán, apenas Aristonectes é conhecido,[9][11] enquanto na formação Quiriquina, ele era contemporâneo de seu parente próximo Wunyelfia [en].[30] Nas formações Dorotea e López de Bertodano, além de Aristonectes, existem muitos elasmossaurídeos indeterminados.[36][37] Exclusivamente na formação López de Bertodano, numerosos gêneros de mosassauros contemporâneos foram identificados. Estes incluem Kaikaifilu [en], Moanasaurus [en], Mosasaurus, Liodon [en] e Plioplatecarpus, embora a validade de alguns desses gêneros seja disputada, pois são baseados principalmente em dentes isolados.[42][43] Alguns desses mosassauros poderiam ter atacado os plesiossauros contemporâneos da formação, incluindo Aristonectes.[44]
Referências
- ↑ a b José P. O'Gorman; Zulma Gasparini; Leonardo Salgado (2013). «Postcranial morphology of Aristonectes (Plesiosauria, Elasmosauridae) from the Upper Cretaceous of Patagonia and Antarctica». Antarctic Science. 25 (1): 71–82. Bibcode:2013AntSc..25...71O. doi:10.1017/S0954102012000673. hdl:11336/11188

- ↑ Claude Gay (1848). «Répteis Fósseis». Zoologia, Vol. 2. Col: Historia Física y Política de Chile [História Física e Política do Chile] (em espanhol). 2. Paris: Imprenta Maulde y Renou. pp. 130–136. Consultado em 30 de agosto de 2023. Cópia arquivada em 29 de agosto de 2023
- ↑ Richard Lydekker (1889). Catalogue of the fossil Reptilia and Amphibia in the British Museum. Part II. Londres: British Museum. p. 222
- ↑ Wilhelm Deecke (1895). «Ueber Saurierreste aus den Quiriquina−Schichten» [Sobre Restos de Dinossauros das Camadas de Quiriquina]. Beiträge zur Geologie und Palaeontologie von Südamerika (em alemão). 14: 32–63
- ↑ Pavel Pravoslavlev (1918). «Геологическое распространенiе эласмозавровъ» [Distribuição geológica de Elasmosaurus]. Bulletin of the Russian Academy of Sciences. VI (em russo). 12 (17): 1955–1978. Consultado em 18 de novembro de 2017. Cópia arquivada em 1 de dezembro de 2017
- ↑ a b c d e f g Ángel Cabrera (1941). «Un Plesiosaurio nuevo del Cretáceo del Chubut» [Um novo Plesiossauro do Cretáceo de Chubut]. Revista del Museo de La Plata (em espanhol). 2 (8): 113–130
- ↑ Edwin H. Colbert (1949). «A new Cretaceous plesiosaur from Venezuela». American Museum Novitates (1420): 1–22. CiteSeerX 10.1.1.1033.3285. hdl:2246/2347
- ↑ Rodrigo A. Otero; Sergio Soto-Acuña; David Rubilar-Rogers (2010). «Presence of Mauisaurus in the Maastrichtian (Late Cretaceous) of central Chile». Acta Palaeontologica Polonica. 55 (2): 361–364. doi:10.4202/app.2009.0065
- ↑ a b c d e f g h i Zulma Gasparini; Nathalie Bardet; James E. Martin; Marta Fernandez (2003). «The elasmosaurid plesiosaur Aristonectes Cabrera from the latest Cretaceous of South America and Antarctica». Journal of Vertebrate Paleontology. 23 (1): 104–115. JSTOR 4524298. doi:10.1671/0272-4634(2003)23[104:TEPACF]2.0.CO;2
- ↑ a b José P. O'Gorman (2013). Plesiosaurios del Cretácico Superior de Patagonia y la península Antártica [Plesiossauros do Cretáceo Superior da Patagônia e da Península Antártica] (PhD) (em espanhol). Universidad Nacional de La Plata. hdl:10915/26140
- ↑ a b c d e f g h i j k José P. O'Gorman (2016). «New Insights on the Aristonectes parvidens (Plesiosauria, Elasmosauridae) Holotype: News on an Old Specimen». Ameghiniana. 52 (4): 397–417. doi:10.5710/AMGH.24.11.2015.2921. hdl:11336/54715
- ↑ a b c d e f g h i j k l m n o p q r s t u Rodrigo A. Otero; Sergio Soto-Acuña; Frank Robin O'Keefe; José P. O'Gorman; Wolfgang Stinnesbeck; Mario E. Suárez; David Rubilar-Rogers; Christian Salazar; Luis Arturo Quinzio-Sinn (2014). «Aristonectes quiriquinensis, sp. nov., a new highly derived elasmosaurid from the upper Maastrichtian of central Chile». Journal of Vertebrate Paleontology. 34 (1): 100–125. Bibcode:2014JVPal..34..100O. JSTOR 24523254. doi:10.1080/02724634.2013.780953. hdl:10915/128448
- ↑ Ben Creisler (2012). «Ben Creisler's Plesiosaur Pronunciation Guide». Oceans of Kansas. Consultado em 26 de junho de 2021. Cópia arquivada em 16 de maio de 2021
- ↑ a b c Samuel P. Welles (1962). «A new species of elasmosaur from the Aptian of Colombia and a review of the Cretaceous plesiosaurs» (PDF). University of California Publications in Geological Sciences. 44 (1): 1–96. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 6 de janeiro de 2024
- ↑ a b c Per Ove Persson (1963). «A revision of the classification of the Plesiosauria with a synopsis of the stratigraphical and geographical distribution of the group» (PDF). Lunds Universitets Arsskrift. 59 (1): 1–59
- ↑ a b c David S. Brown (1981). «The English Late Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria». Bulletin of the British Museum (Natural History), Geology. 35 (4): 253–347
- ↑ a b Rodrigo A. Otero; Sergio Soto-Acuña; David Rubilar-Rogers (2012). «A postcranial skeleton of an elasmosaurid plesiosaur from the Maastrichtian of central Chile, with comments on the affinities of Late Cretaceous plesiosauroids from the Weddellian Biogeographic Province». Cretaceous Research. 37: 89–99. Bibcode:2012CrRes..37...89O. doi:10.1016/j.cretres.2012.03.010
- ↑ Rodrigo A. Otero; José P. O'Gorman (2013). «Identification of the first postcranial skeleton of Aristonectes Cabrera (Plesiosauroidea, Elasmosauridae) from the upper Maastrichtian of the south-eastern Pacific, based on a bivariate graphic analysis». Cretaceous Research. 41: 86–89. Bibcode:2013CrRes..41...86O. doi:10.1016/j.cretres.2012.11.001
- ↑ Mario E. Suárez; Omar Fritis (2002). «Nuevo registro de Aristonectes sp. (Plesiosauroidea incertae sedis) del Cretácico Tardío de la Formación Quiriquina, Cocholgüe, Chile» [Novo registro de Aristonectes sp. (Plesiosauroidea incertae sedis) do Cretáceo Superior da Formação Quiriquina, Cocholgüe, Chile]. Boletín de la Sociedad de Biología de Concepción (em espanhol). 73: 87–93
- ↑ a b Rodrigo A. Oterio; José P. O'Gorman; Norton Hiller (2015). «Reassessment of the upper Maastrichtian material from Chile referred to Mauisaurus Hector, 1874 (Plesiosauroidea: Elasmosauridae) and the taxonomical value of the hemispherical propodial head among austral elasmosaurids». New Zealand Journal of Geology and Geophysics. 58 (3): 252–261. Bibcode:2015NZJGG..58..252O. doi:10.1080/00288306.2015.1037775. hdl:11336/53493
- ↑ a b c d e f g h i j k l m n o p q r Rodrigo A. Otero; Sergio Soto-Acuña; Frank R. O'keefe (2018). «Osteology of Aristonectes quiriquinensis (Elasmosauridae, Aristonectinae) from the upper Maastrichtian of central Chile». Journal of Vertebrate Paleontology. 38 (1). Bibcode:2018JVPal..38E8638O. JSTOR 44865847. doi:10.1080/02724634.2017.1408638
- ↑ a b José P. O'Gorman; Sergio N. Santillana; Rodrigo A. Otero; Marcelo A. Reguero (2019). «A giant elasmosaurid (Sauropterygia; Plesiosauria) from Antarctica: New information on elasmosaurid body size diversity and aristonectine evolutionary scenarios» (PDF). Cretaceous Research. 102: 37–58. Bibcode:2019CrRes.102...37O. doi:10.1016/j.cretres.2019.05.004. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 29 de agosto de 2023
- ↑ a b Sven Sachs; Benjamin P. Kear (2015). «Fossil Focus: Elasmosaurs». www.palaeontologyonline.com. Palaeontology Online. pp. 1–8. Consultado em 18 de janeiro de 2018. Cópia arquivada em 1 de fevereiro de 2018
- ↑ a b R. Araújo; Michael J. Polcyn; Johan Lindgren; L. L. Jacobs; A. S. Schulp; O. Mateus; A. Olimpio Gonçalves; M-L. Morais (2015). «New aristonectine elasmosaurid plesiosaur specimens from the Early Maastrichtian of Angola and comments on paedomorphism in plesiosaurs.» (PDF). Netherlands Journal of Geosciences. 94 (1): 93–108. Bibcode:2015NJGeo..94...93A. doi:10.1017/NJG.2014.43. Cópia arquivada (PDF) em 11 de julho de 2020
- ↑ a b Rodrigo A. Otero (2016). «Taxonomic reassessment of Hydralmosaurus as Styxosaurus: new insights on the elasmosaurid neck evolution throughout the Cretaceous». PeerJ. 4. PMC 4806632. PMID 27019781. doi:10.7717/peerj.1777
- ↑ F. Robin O'Keefe; William Wahl Jr. (2003). «Preliminary report on the osteology and relationships of a new aberrant cryptocleidoid plesiosaur from the Sundance Formation, Wyoming». Paludicola. 4 (2): 48–68. Consultado em 30 de agosto de 2023. Cópia arquivada em 21 de novembro de 2023
- ↑ F. Robin O'Keefe; Hallie P. Street (2009). «Osteology Of The cryptoclidoid plesiosaur Tatenectes laramiensis, with comments on the taxonomic status of the Cimoliasauridae» (PDF). Journal of Vertebrate Paleontology. 29 (1): 48–57. Bibcode:2009JVPal..29...48O. doi:10.1671/039.029.0118. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 3 de março de 2012
- ↑ F. Robin O'Keefe; Hallie P. Street; Benjamin C. Wilhelm; Courtney D. Richards; Helen Zhu (2011). «A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape among plesiosaurs». Journal of Vertebrate Paleontology. 31 (2): 330–339. Bibcode:2011JVPal..31..330O. doi:10.1080/02724634.2011.550365. Consultado em 30 de agosto de 2023. Cópia arquivada em 21 de novembro de 2023
- ↑ a b José P. O'Gorman (2020). «Elasmosaurid phylogeny and paleobiogeography, with a reappraisal of Aphrosaurus furlongi from the Maastrichtian of the Moreno Formation». Journal of Vertebrate Paleontology. 39 (5). Bibcode:2019JVPal..39E2025O. doi:10.1080/02724634.2019.1692025. Consultado em 14 de setembro de 2023. Cópia arquivada em 29 de setembro de 2023
- ↑ a b c Rodrigo A. Otero; Sergio Soto-Acuña (2020). «Wunyelfia maulensis gen. et sp. nov., a new basal aristonectine (Plesiosauria, Elasmosauridae) from the Upper Cretaceous of central Chile». Cretaceous Research. 188. Bibcode:2021CrRes.11804651O. doi:10.1016/j.cretres.2020.104651
- ↑ a b José P. O'Gorman; Eduardo B. Olivero; Sergio Santillana; Michael J. Everhart; Marcelo Reguero (2014). «Gastroliths associated with an Aristonectes specimen (Plesiosauria, Elasmosauridae), López de Bertodano Formation (upper Maastrichtian) Seymour Island (Is. Marambio), Antarctic Peninsula». Cretaceous Research. 50: 228–237. Bibcode:2014CrRes..50..228O. doi:10.1016/j.cretres.2014.03.011. hdl:11336/7400
- ↑ Oliver Wings (2007). «A review of gastrolith function with implications for fossil vertebrates and a revised classification». Acta Palaeontologica Polonica. 52 (1): 1–16. Consultado em 30 de agosto de 2023. Cópia arquivada em 14 de abril de 2021
- ↑ Michael J. Everhart (2000). «Gastroliths Associated with Plesiosaur Remains in the Sharon Springs Member of the Pierre Shale (Late Cretaceous), Western Kansas». Transactions of the Kansas Academy of Science. 103 (1–2): 64–75. JSTOR 3627940. doi:10.2307/3627940
- ↑ David J. Cicimurri; Michael J. Everhart (2001). «An Elasmosaur with Stomach Contents and Gastroliths from the Pierre Shale (Late Cretaceous) of Kansas». Transactions of the Kansas Academy of Science. 104 (3–4): 129–143. JSTOR 3627807. doi:10.1660/0022-8443(2001)104[0129:AEWSCA]2.0.CO;2. Consultado em 30 de agosto de 2023. Cópia arquivada em 29 de agosto de 2023
- ↑ Ignacio A. Cerda; Leonardo Salgado (2008). «Gastrolitos en un plesiosaurio (Sauropterygia) de la Formación Allen (Campaniano-Maastrichtiano), provincia de Río Negro, Patagonia, Argentina». Ameghiniana (em espanhol). 45 (8): 529–536. ISSN 0002-7014
- ↑ a b c d Rodrigo A. Otero; Sergio Soto-Acuña; Christian Salazar; José L. Oyarzún (2015). «New elasmosaurids (Sauropterygia, Plesiosauria) from the Late Cretaceous of the Magallanes Basin, Chilean Patagonia: Evidence of a faunal turnover during the Maastrichtian along the Weddellian Biogeographic Province». Andean Geology. 42 (2): 237–267. doi:10.5027/andgeoV42n1-a05. hdl:11336/21148
- ↑ a b José P. O'Gorman; Karen M. Panzeri; Marta S. Fernández; Sergio Santillana; Juan J. Moly; Marcelo Reguero (2018). «A new elasmosaurid from the upper Maastrichtian López de Bertodano Formation: new data on weddellonectian diversity». Alcheringa: An Australasian Journal of Palaeontology. 42 (4): 575–586. Bibcode:2018Alch...42..575O. doi:10.1080/03115518.2017.1339233. hdl:11336/49635
- ↑ Wolfgang Stinnesbeck (1986). «Zu den faunistischen und palökologischen Verhältnissen in der Quiriquina Formation (Maastrichtium) Zentral-Chiles» [Sobre as condições faunísticas e paleoecológicas na Formação Quiriquina (Maastrichtiano) do Chile central]. Palaeontographica Abteilung A (em alemão). 194 (4–6): 99–237
- ↑ Agustín Martinelli; Analía Forasiepi (2004). «Late Cretaceous vertebrates from Bajo de Santa Rosa (Allen Formation), Rio Negro province, Argentina, with the description of a new sauropod dinosaur (Titanosauridae)». Revista del Museo Argentino de Ciencias Naturales. 6 (2): 257–305. doi:10.22179/revmacn.6.88
- ↑ María F. Rodríguez; Héctor A. Leanza; Matías Salvarredy Aranguren (2007). «Provincias del Neuquén, Río Negro y La Pampa» [Províncias de Neuquén, Río Negro e La Pampa]. Servicio Geológico Minero Argentino - Instituto de Geología y Recursos Minerales (em espanhol) (370): 32-35
- ↑ Alberto L. Cione; Sergio Santillana; Soledad Gouiric-Cavalli; Carolina Acosta Hospitaleche; Javier N. Gelfo; Guillermo M. Lopez; Marcelo Reguero (2018). «Before and after the K/Pg extinction in West Antarctica: New marine fish records from Marambio (Seymour) Island». Cretaceous Research. 85: 250–265. Bibcode:2018CrRes..85..250C. doi:10.1016/j.cretres.2018.01.004. hdl:10915/147537
- ↑ Rodrigo A. Otero; Sergio Soto-Acuña; David Rubilar-Rogers; Carolina S. Gutstein (2017). «Kaikaifilu hervei gen. et sp. nov., a new large mosasaur (Squamata, Mosasauridae) from the upper Maastrichtian of Antarctica». Cretaceous Research. 70: 209–225. Bibcode:2017CrRes..70..209O. doi:10.1016/j.cretres.2016.11.002
- ↑ Pablo Gonzalez Ruiz; Marta S. Fernandez; Marianella Talevi; Juan M. Leardi; Marcelo A. Reguero (2019). «A new Plotosaurini mosasaur skull from the upper Maastrichtian of Antarctica. Plotosaurini paleogeographic occurrences». Cretaceous Research. 103: 104166. Bibcode:2019CrRes.10304166G. doi:10.1016/j.cretres.2019.06.012. hdl:11336/125124
- ↑ Universidade do Chile (7 de novembro de 2016). «Um lagarto predador gigante nadava nos mares da Antártida perto do fim da era dos dinossauros». ScienceDaily
Ligações externas
- Informações e fotos de Aristonectes, The Plesiosaur Directory
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||






.png)