Tsintaosaurus
Tsintaosaurus
| |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Campaniano | |||||||||||||||||||||
 Crânio holótipo com crista quebrada | |||||||||||||||||||||
| Classificação científica | |||||||||||||||||||||
| |||||||||||||||||||||
| Espécie-tipo | |||||||||||||||||||||
| †Tsintaosaurus spinorhinus Young, 1958[1] | |||||||||||||||||||||
Tsintaosaurus ([sɪntaʊˈsɔːrəs]; sic para a antiga transliteração [en] "Tsingtao",[2] significando "lagarto de Qingdao") é um gênero de dinossauro hadrossaurídeo da China. Tinha cerca de 8,3 m de comprimento e pesava 2,5 toneladas.[3] A espécie-tipo é Tsintaosaurus spinorhinus, descrita pela primeira vez pelo paleontólogo chinês C. C. Young em 1958. Como um hadrossauro, Tsintaosaurus tinha o característico focinho de 'bico de pato' e uma bateria de dentes poderosos que usava para mastigar vegetação. Ele geralmente andava de quatro, mas podia se erguer nas patas traseiras para procurar predadores e fugir se avistasse algum. Como outros hadrossauros, Tsintaosaurus provavelmente vivia e se deslocava em manadas.
Descoberta e nomeação
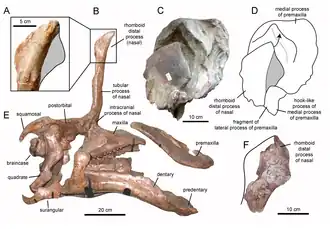
Em 1950, em Hsikou, perto de Chingkangkou, em Laiyang [en], Xantum, na parte oriental da China, vários restos de grandes hadrossaurídeos foram descobertos. Em 1958, estes foram descritos pelo paleontólogo chinês Yang Zhongjian ("C.C. Young") como a espécie-tipo Tsintaosaurus spinorhinus. O nome genérico é derivado da cidade de Qingdao, anteriormente muitas vezes transliterada como "Tsintao". O nome específico significa "com um espinho nasal", do latim spina, e do grego ῥίς, rhis, "nariz", em referência à crista distinta no focinho.[1]
O holótipo, IVPP AS V725, foi descoberto em uma camada da formação Jingangkou, parte do grupo Wangshi, datada do Campaniano. Consiste em um esqueleto parcial com crânio. O parátipo é o espécime IVPP V818, um teto craniano. Na mesma área, foram encontrados alguns esqueletos parciais adicionais e um grande número de elementos esqueléticos desarticulados. Alguns destes foram por Yang referidos a Tsintaosaurus, outros foram nomeados como Tanius chingkankouensis Yang 1958; também existe um Tanius laiyangensis Zhen 1976. As duas últimas espécies são hoje consideradas sinônimos juniores ou nomina dubia. Pesquisadores posteriores refeririam uma parte maior do material a Tsintaosaurus.
Descrição
Crista

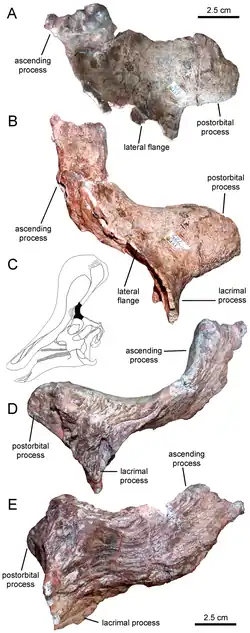
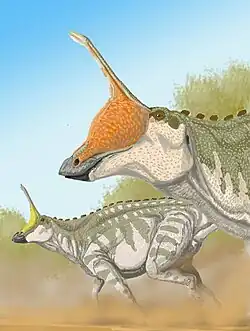
Tsintaosaurus foi originalmente reconstruído com uma crista semelhante à de um unicórnio em seu crânio. A crista, como preservada, consiste em um processo de cerca de quarenta centímetros de comprimento, protuberante quase verticalmente do topo da parte traseira do focinho. A estrutura é oca e parece ter uma extremidade superior bifurcada. Estruturas comparáveis com espécies relacionadas são desconhecidas: elas possuem cristas mais parecidas com lobos. Em 1990, David B. Weishampel [en] e Jack Horner puseram em dúvida a presença da crista, sugerindo que era na verdade um osso nasal quebrado do topo do focinho, distorcido para cima por um esmagamento do fóssil. Seu estudo sugeriu ainda que, sem a crista distinta para distingui-lo, Tsintaosaurus era na verdade um sinônimo do hadrossauro semelhante, mas sem crista, Tanius. No entanto, em 1993, Éric Buffetaut [en] e outros, após uma nova investigação dos próprios ossos, concluíram que a crista não estava distorcida nem era um artefato de restauração; além disso, um segundo espécime com uma parte da crista ereta havia sido descoberto desde então, indicando que a crista era de fato real e Tsintaosaurus é provavelmente um gênero distinto.[4]
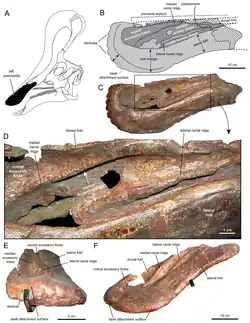
Uma nova reconstrução em 2013, por Albert Prieto-Márquez e Jonathan Wagner e baseada na identificação do espécime IVPP V829, uma pré-maxila, como um elemento de Tsintaosaurus, chegou à conclusão de que o osso semelhante ao de um unicórnio era apenas a parte traseira de uma crista craniana maior que começava na ponta do focinho. A frente da crista teria sido formada por processos ascendentes das pré-maxilas. Estes tinham facetas de contato romboides expandidas com as partes superiores expandidas dos processos da crista dos ossos nasais, formando a parte traseira da crista. A base traseira da crista era coberta por crescimentos dos pré-frontais. Os ossos nasais fundidos teriam formado uma estrutura tubular oca. A altura da crista teria excedido a da parte traseira do crânio, medida ao longo dos quadrados. Embora em grande parte vertical, a crista é direcionada ligeiramente para trás; a inclinação para a frente da crista do holótipo seria o resultado de uma distorção do fóssil.[5]
A nova reconstrução por Prieto-Márquez e Wagner também levou a uma nova hipótese sobre as passagens de ar internas da crista. Yang havia assumido que a cavidade tubular na parte preservada do holótipo teria servido como a principal entrada de ar. Isso foi rejeitado por Prieto-Márquez e Wagner, que apontaram que o tubo era fechado em sua extremidade inferior e que, com os lambeossaurinos em geral, as passagens de ar estão localizadas em uma posição mais avançada, com as narinas ósseas sendo completamente fechadas pelas pré-maxilas. Eles assumiram que Tsintaosaurus teria um arranjo lambeossaurino padrão no focinho, com o ar, ao inalar, entrando no crânio através das pseudo-narinas pareadas, as "falsas narinas" das pré-maxilas atrás do bico superior. De lá, o ar teria sido transportado através de passagens pareadas abaixo dos processos medianos das pré-maxilas para o topo da crista, subsequentemente entrando em uma câmara mediana comum dentro do lobo. A parte traseira da câmara era formada pelos ossos nasais e provavelmente homóloga à cavidade nasal. A câmara era dividida em duas cavidades menores, uma na frente e outra atrás, por processos medianos curvos das pré-maxilas, formando ganchos ao redor de uma passagem entre as cavidades. Da cavidade traseira, o ar era transportado para baixo, em direção à cavidade craniana interna. Embora geralmente se assuma que uma única passagem servia a esse propósito, Prieto-Márquez e Wagner viram indicações na forma do nasal de que havia passagens descendentes pareadas, para o interior dos processos laterais das pré-maxilas. Disso, eles concluíram que todo o fluxo de ar era provavelmente separado, com a câmara mediana comum provavelmente sendo dividida em uma seção esquerda e direita por um septo cartilaginoso.[5]

A conclusão de que a estrutura tubular nos ossos nasais traseiros não era uma passagem de ar forçou Prieto-Márquez e Wagner a encontrar uma explicação alternativa para sua função. Eles sugeriram que ela teria servido para diminuir o peso da crista, com tal tubo combinando resistência relativa com baixa massa óssea. Tsintaosaurus teria diferido nisso dos lambeossaurinos mais derivados, que têm uma extensão frontal do osso frontal, na forma de uma lâmina óssea, suportando a crista.[5]
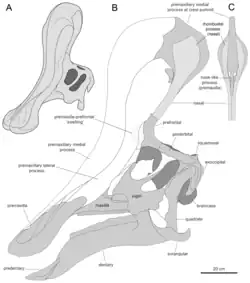
Outras características distintivas
Além da crista, Prieto-Márquez e Wagner identificaram várias outras características distintivas (autapomorfias) de Tsintaosaurus. A borda do bico superior é arredondada e espessa, mais larga que a largura transversal da depressão frontal ao redor das narinas. Na medida em que essa depressão está situada nas pré-maxilas, ela é dividida longitudinalmente em cada lado por duas cristas que continuam obliquamente para baixo e para os lados. Internamente, os ossos nasais fundidos formam um bloco ósseo na frente da caixa craniana. A parte traseira do osso nasal é cortada por extensões frontais do osso frontal, a mais alta das quais é elevada em relação ao teto do crânio. Os ramos ascendentes das pré-maxilas têm processos internos apontando para trás, para baixo e ligeiramente para dentro, dividindo uma câmara compartilhada na linha média. O pré-frontal possui um flange, continuando da parte inferior do osso lacrimal até a parte inferior do processo ascendente do pré-frontal, e conectando-se a um processo na lateral da pré-maxila para formar uma elevação na base lateral da crista. A lateral e a parte inferior do pré-frontal mostram sulcos verticais profundos. A fenestra supratemporal é, transversalmente, mais larga do que longa.[5]
Classificação
Tsintaosaurus pode formar um clado em Lambeosaurinae com os gêneros europeus Pararhabdodon [en] e Koutalisaurus [en] (provável sinônimo de Pararhabdodon).[6]
A posição de Tsintaosaurus na árvore evolutiva, de acordo com um estudo de 2013 por Prieto-Márquez et al., é indicada por este cladograma:[7]
| Lambeosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecologia
Um estudo de ovos de dinossauro em camadas sucessivas da série Wangshi da província de Shandong, da qual a formação Jingangkou é a camada mais recente, mostra que a região tinha uma alta diversidade de dinossauros e que o clima havia se tornado mais seco desde a formação Jiangjunding precedente.[8]
Referências
- ↑ a b Young, C.-C. (1958). «The dinosaurian remains of Laiyang, Shantung». Whole Number. Palaeontologia Sinica, New Series C. 42 (16): 1–138
- ↑ Creisler, B. (2002). «Dinosauria Translation and Pronunciation Guide T». DOL Dinosaur Omnipedia. Consultado em 24 de fevereiro de 2010. Cópia arquivada em 31 de dezembro de 2005
- ↑ Gregory S. Paul (2010). The Princeton Field Guide to Dinosaurs. Estados Unidos: Princeton University Press. pp. 308. ISBN 978-0-691-13720-9
- ↑ Buffetaut, E.; Tong, H. (1993). «Tsintaosaurus spinorhinus Young and Tanius sinensis Wiman: a preliminary comparative study of two hadrosaurs (Dinosauria) from the Upper Cretaceous of China». C.R. Academy of Science Paris. 2. 317: 1255–1261
- ↑ a b c d Prieto-Márquez, A.; Wagner, J. R. (2013). «The 'Unicorn' Dinosaur That Wasn't: A New Reconstruction of the Crest of Tsintaosaurus and the Early Evolution of the Lambeosaurine Crest and Rostrum». PLOS ONE. 8 (11). Bibcode:2013PLoSO...882268P. PMC 3838384
 . PMID 24278478. doi:10.1371/journal.pone.0082268
. PMID 24278478. doi:10.1371/journal.pone.0082268
- ↑ Prieto-Márquez, A.; Wagner, J. R. (2009). «Pararhabdodon isonensis and Tsintaosaurus spinorhinus: a new clade of lambeosaurine hadrosaurids from Eurasia». Cretaceous Research. online preprint (5): 1238–1246. Bibcode:2009CrRes..30.1238P. doi:10.1016/j.cretres.2009.06.005. hdl:2152/41080
- ↑ Prieto-Marquez, A.; Vecchia, F. M. D.; Gaete, R.; Galobart, A. (2013). «Diversity, relationships, and biogeography of the Lambeosaurine dinosaurs from the European archipelago, with description of the new aralosaurin Canardia garonnensis». PLOS ONE. 8 (7). Bibcode:2013PLoSO...869835P. PMC 3724916. PMID 23922815. doi:10.1371/journal.pone.0069835
- ↑ Zhao, ZiKui; Zhang, ShuKang; Wang, Qiang; Wang, XiaoLin (2013). «Dinosaur diversity during the transition between the middle and late parts of the Late Cretaceous in eastern Shandong Province, China: Evidence from dinosaur eggshells». Chinese Science Bulletin. 58 (36): 4663–4669. Bibcode:2013ChSBu..58.4663Z. doi:10.1007/s11434-013-6059-9
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
