Knoetschkesuchus
Knoetschkesuchus
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Kimmeridgiano | |||||||||||||||||||
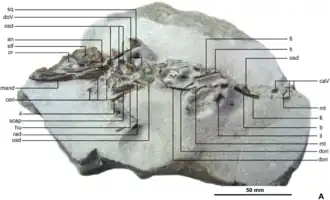 Espécime-tipo de Knoetschkesuchus langenbergensis | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Espécie-tipo | |||||||||||||||||||
| Knoetschkesuchus langenbergensis Schwarz et al., 2017 | |||||||||||||||||||
| Espécies | |||||||||||||||||||
| |||||||||||||||||||
Knoetschkesuchus é um gênero de pequeno crocodilomorfo atopossaurídeo do Jurássico Superior da Alemanha e de Portugal. Duas espécies são conhecidas: a espécie alemã K. langenbergensis, descrita por Schwarz e colegas em 2017 com base em dois esqueletos parciais e vários ossos isolados; e a espécie portuguesa K. guimarotae, nomeada a partir de mais de 400 espécimes, incluindo vários esqueletos parciais. Knoetschkesuchus era um crocodiliano pequeno e de focinho curto, medindo cerca de 55 cm de comprimento, que se alimentava principalmente de pequenas presas, incluindo invertebrados, anfíbios e mamíferos. Essa especialização em pequenas presas separava ecologicamente Knoetschkesuchus da maioria dos outros diversos crocodilianos com os quais convivia no ecossistema insular da Europa jurássica.
Ambas as espécies foram anteriormente reconhecidas como pertencentes a Theriosuchus; K. guimarotae foi inicialmente nomeada como T. guimarotae, e espécimes de K. langenbergensis foram inicialmente referidos a T. pusillus após sua descoberta. Schwarz e colegas reconheceram uma série de características que uniam esses dois táxons, excluindo outras espécies de Theriosuchus; em particular, Knoetschkesuchus possui apenas dois tipos distintos de dentes, faltando os dentes em forma de folha vistos em outros atopossaurídeos. Outras características distintivas incluem o crânio relativamente largo e a presença das fenestras antorbital e mandibular em todas as fases da vida.
Descoberta e nomeação
Knoetschkesuchus langenbergensis
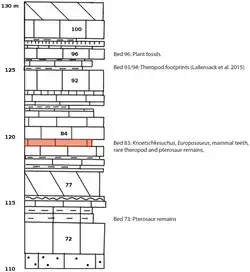
O material da espécie-tipo de Knoetschkesuchus, K. langenbergensis, provém de um leito de calcário margoso (numerado como Leito 83, não Leito 93 como relatado em algumas publicações) contendo intraclastos [en] micríticos [en] recristalizados, localizado na pedreira de Langenberg, nas montanhas Harz, perto de Goslar, Baixa Saxônia, Alemanha. Esses depósitos foram datados do estágio Kimmeridgiano Superior do Jurássico — especificamente, ao equivalente regional do Kimmeridgiano Superior conhecido como Mittleres Kimmeridge,[1] e pertencem à formação Süntel [en].[2] Embora bem preservados, os fósseis desta pedreira foram recuperados geralmente por operações regulares de detonação na pedreira.[3]
Os espécimes pertencentes a Knoetschkesuchus estão armazenados no Dinosaurier-Freilichtmuseum Münchehagen [de] (doravante denotado por DFMMh/FV) em Rehburg-Loccum, Alemanha. São eles: o espécime-tipo DFMMh/FV 200, o esqueleto parcial de um adulto com um crânio; DFMMh/FV 605, o crânio completo de um juvenil; DFMMh/FV 261, um osso angular [en] isolado; DFMMh/FV 790.12, um dentário esquerdo isolado; DFMMh/FV 279, um fêmur isolado; DFMMh/FV 790.11, um metatarso isolado; e DFMMh/FV 325, um esqueleto parcial incluindo osteodermas, vértebras e costelas. Foi utilizada a estereomicroscopia [en] para examinar os espécimes, que foram descritos em um artigo de 2017 publicado por Daniela Schwarz, Maik Raddatz e Oliver Wings.[3]
O nome do gênero Knoetschkesuchus combina o sobrenome de Nils Knötschke, um pesquisador do DFMMh responsável pela coleta, curadoria e preparação dos espécimes da pedreira de Langenberg, com o sufixo comum de crocodilianos suchus, do grego souchos ("crocodilo"). Enquanto isso, o nome da espécie langenbergensis é uma referência à proveniência desta espécie da pedreira de Langenberg.[3]
Knoetschkesuchus guimarotae
Schwarz et al. também atribuíram uma espécie adicional a Knoetschkesuchus, K. guimarotae, que foi originalmente classificada como uma espécie de Theriosuchus. Como reflete o nome específico, K. guimarotae provém das camadas de lignito da pedreira de Guimarota, localizada perto de Leiria, Portugal. Existem dois estratos de linhito principais com vertebrados dentro dos chamados estratos de Guimarota, que são separados por calcário margoso e conhecidos respectivamente como subunidades "Fundschichten" e "Ruafolge"; K. guimarotae é conhecido de ambas as camadas. Elas foram atribuídas à formação Alcobaça [en], uma subunidade da formação Abadia,[4] que foi datada do Kimmeridgiano com base em ostracodes fósseis.[5][6][7]
Os restos de K. guimarotae estão armazenados no Instituto de Ciências Geológicas da Universidade Livre de Berlim (doravante IPFUB). Juntamente com o espécime-tipo IPFUB Gui Croc 7308 — que consiste em um crânio parcial com mandíbulas, uma vértebra do sacro e duas osteodermas — são conhecidos mais de 400 espécimes adicionais, a maioria consistindo em ossos isolados. Entre estes, os espécimes mais completos são IPFUB Gui Gui Croc 7352 (vértebras da cauda, fêmur, osteodermas); 7441 (osteodermas e ulna); 7545 (vértebras dorsais, ísquios, osteodermas); 7564 (fêmur, úmero, osteodermas); 7634 (vértebras dorsais, costela, osteodermas); e 8037 (vértebras cervicais e dorsais, e osteodermas). Mais espécimes são conhecidos, mas permanecem por preparar. Alguns espécimes mostram marcas de carniça.[5]
Descrição
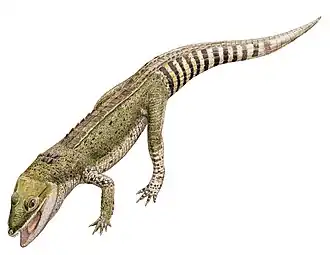
Como outros membros de Atoposauridae,[3] Knoetschkesuchus é muito pequeno, com Knoetschkesuchus guimarotae medindo apenas 55 cm de comprimento no máximo.[5] Típico do grupo, ambas as espécies eram quadrúpedes, possuindo membros longos e delgados.[8] As costas de ambas as espécies conhecidas são cobertas por duas fileiras de osteodermas ósseas retangulares, centradas na linha média, que são mais largas do que longas.[3] Cada osteoderma possui uma quilha que corre longitudinalmente, embora as quilhas sejam menos desenvolvidas em K. langenbergensis.[3] Em pelo menos K. guimarotae, as osteodermas na cauda têm quilhas mais afiadas e mais altas; as próximas à parte de trás da cauda tendem a ser mais longas do que largas, ao contrário das outras osteodermas, e também são abobadadas. As vértebras de K. guimarotae são adicionalmente anficélicas, ou côncavas em ambas as extremidades;[5] embora inicialmente proposta como uma característica única, T. pusillus também possui vértebras anficélicas.[3][9] O crânio e as mandíbulas, que exibem uma série de características que separam Knoetschkesuchus de outros atopossaurídeos, são descritos em mais detalhe abaixo.
Focinho
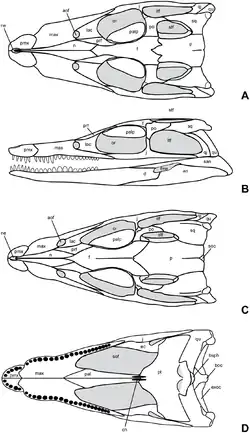
O crânio de Knoetschesuchus é relativamente curto, com o focinho ocupando 47% do comprimento do crânio em K. langenbergensis e 42% em K. guimarotae,[5] o que permite que sejam classificados como crocodilianos brevirrostrinos.[10] Ao longo do lado do focinho há duas ondulações, uma menor na pré-maxila e uma maior e mais larga na maxila. A sutura irregular entre a pré-maxila e a maxila é angulada para a frente do crânio em K. langenbergensis e para trás em K. guimarotae.[5] Ao longo da linha média do focinho estão os nasais finos e em forma de cunha; as narinas, que se voltam para cima, são claramente separadas pelos nasais em K. guimarotae,[5] mas não está claro se este é o caso em K. langenbergensis.[3]
Na parte de trás, os nasais são separados pelo osso frontal em K. langenbergensis, cujo terço posterior é um tanto abobadado. A porção do frontal entre os olhos tem um terço da largura de todo o crânio em ambas as espécies, mas alarga-se mais para trás para formar a frente do teto do crânio. O osso pré-frontal [en] é reto ao longo de seu contato com o frontal e o nasal (cerca de metade do osso está em contato com cada um), mas forma um ângulo entre a margem da órbita ocular e o lacrimal do outro lado. Este ângulo é arredondado em K. guimarotae de modo que o osso tem formato oval,[5] mas muito pontiagudo em K. langenbergensis de modo que o osso é triangular.[3]
Órbita ocular e teto do crânio
O corpo principal do lacrimal é um quadrado arredondado com ambas as faces do osso côncavas. Seu contato com o nasal é bastante limitado em ambas as espécies. A fenestra antorbital de formato oval é pequena, tendo apenas 9% do comprimento da órbita ocular; sua presença é única para ambas as espécies do gênero entre os atopossaurídeos. Enquanto isso, a órbita ocular é grande e oval, sendo 54% mais longa do que alta. Os ossos palpebrais em forma de gota projetam-se do topo das órbitas oculares. Em ambas as espécies, os dois terços posteriores da superfície interna do palpebral são ligeiramente côncavos. Ambas as espécies têm um osso esquamosal em que o terço posterior é chanfrado; em K. langenbergensis, a margem externa é um tanto convexa.[3]
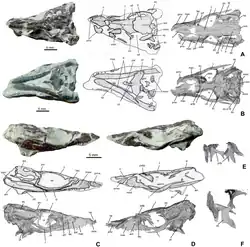
Visto de cima, o osso parietal aumenta de largura na parte de trás; o aumento é pequeno em K. langenbergensis, de modo que o osso é retangular no geral, mas a diferença é maior em K. guimarotae. A parte de trás do osso apresenta um pequeno entalhe em K. langenbergensis e uma concavidade geral em K. guimarotae que expõe ligeiramente o osso supraoccipital subjacente.[5] O osso pós-orbital [en] possui dois ramos que se unem com uma curva suave, separados por um ângulo de 130° em K. langenbergensis. A fenestra supratemporal é aproximadamente quadrada em K. guimarotae,[5] mas tem uma extremidade posterior mais fina em K. langenbergensis. Em ambas as espécies, a distância máxima entre as fenestras supratemporais é cerca de um terço da largura total do topo do crânio. A fenestra infratemporal trapezoidal é 1,5 vezes mais longa do que larga em K. langenbergensis.[3]
Palato e caixa craniana
Na parte inferior do crânio, o osso pterigoide é cerca de duas vezes mais largo do que longo. Na frente do pterigoide há uma pequena projeção que se estende para trás para formar uma crista, de cada lado da qual há uma depressão em forma de sulco (o sulco coanal) contendo as coanas. Em Theriosuchus, ao contrário de Knoetschkesuchus, as coanas estão inseridas em uma depressão mais ampla em forma de tigela. Além disso, em ambas as espécies, a superfície inferior do pterigoide é um tanto côncava. Estendendo-se para a frente de cada lado do pterigoide está o ectopterigoide; em ambas as espécies, este osso é constrito perto do meio para formar uma forma de ampulheta, mas em K. langenbergensis também é um tanto torcido para o lado. A parte de trás do osso é muito côncava em ambas as espécies.[3][5]
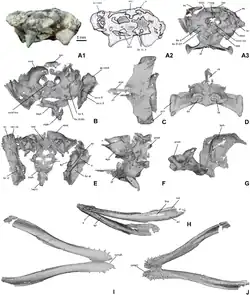
Formando os lados da parte de trás do crânio está o exoccipital, que circunda a maior parte do forame magno. A trompa de Eustáquio estende-se para baixo através do basioccipital e do esfenoide; esses ossos são espessados de cada lado da trompa em K. langenbergensis. K. guimarotae tem um pequeno forame arredondado ao lado da trompa no basioccipital e uma tuberosidade com uma crista acima.[5] Na frente do basioccipital em ambas as espécies, há duas depressões arredondadas perto da base. O esfenoide se assemelha a um machado triangular quando visto de lado.[3]
Mandíbula
Em K. guimarotae, as duas metades do dentário divergem uma da outra em um ângulo de 20° perto da frente e 40° perto de trás.[5] Também possui duas convexidades na parte inferior da mandíbula, uma no terceiro e quarto dentes e outra do oitavo ao décimo dentes. A última convexidade é substituída por uma concavidade em K. langenbergensis. Em ambas as espécies, a margem superior da mandíbula atrás da fileira de dentes inclina-se para cima em linha reta. O lado do dentário é pontuado, embora muito mais densamente em K. langenbergensis. Perto da parte de trás do dentário em ambas as espécies, os pontos são substituídos por sulcos longitudinais. No interior da mandíbula, o esplenial possui um forame oval atrás do nível da sínfise mandibular em ambas as espécies, e o topo do osso possui uma crista baixa e áspera em K. guimarotae.[3][5]
A ponta do angular está situada perto do ponto médio do osso em K. langenbergensis, em vez de estar na parte de trás como em K. guimarotae.[5] Em ambas as espécies, a parte de trás do angular contribui para o processo retroarticular. A superfície interna do angular é áspera em K. guimarotae, e a margem superior da parede interna faz a transição de uma ponta projetada para cima na frente para uma crista baixa e arredondada na parte de trás;[5] a mesma margem mantém sua altura ao longo do angular em K. langenbergensis. O surangular possui um processo fino e projetado para a frente que, em K. langenbergensis, se estende para a frente até a parte de trás da fileira de dentes e possui um sulco na parte inferior.[5] A presença da fenestra mandibular externa de formato oval[5] é única para o gênero entre os atopossaurídeos.[3]
Dentes

Ambas as espécies de Knoetschkesuchus possuem apenas dois tipos distintos de dentes, unicamente entre os atopossaurídeos. Outros atopossaurídeos têm um terceiro tipo de dentes em forma de folha. O primeiro tipo de dentes, encontrado perto da frente da mandíbula, é cônico, ligeiramente constrito na base e curvado ligeiramente para dentro.[5] Um subtipo é representado pelos dentes semelhantes a caninos ou pseudocaniniformes. Eles são o quarto e o quinto dentes maxilares e são aumentados (cerca de duas vezes o tamanho dos outros dentes maxilares), mais pontiagudos e mais constritos na base. O segundo tipo de dentes, que constitui o resto dos dentes, tem a forma de finas pontas de lança, com uma base larga e uma ponta mais estreita; em K. guimarotae todas as pontas são afiadas,[5] mas em Knoetschesuchus langenbergensis elas gradualmente se tornam mais rombas.[3]
Existem 5 dentes pré-maxilares em ambas as espécies,[5] o quarto dos quais é cerca de um terço maior em relação aos outros. A maxila tem 12 alvéolos distintos em K. langenbergensis. Excluindo os pseudocaniniformes, o tamanho do dente maxilar permanece aproximadamente constante, mas diminui gradualmente após os pseudocaniniformes em ambas as espécies; todos os alvéolos dentários após o décimo segundo são substituídos por um sulco contínuo. No total, K. guimarotae tinha pelo menos 15 dentes maxilares,[5] e K. langenbergensis 17 ou 18. Enquanto isso, o dentário exibe 21 dentes em K. langenbergensis e pelo menos 20 em K. guimarotae.[5] Assim como na maxila, os alvéolos distintos para os dentes do dentário são substituídos por um sulco a partir do décimo primeiro dente para trás em K. langenbergensis.[3]
Classificação

Quando os espécimes conhecidos de K. langenbergensis foram descritos pela primeira vez de forma preliminar num artigo de 2006 de Hans-Volker Karl et al., eles foram referidos ao gênero Theriosuchus. Isso se baseou no crânio curto, narinas divididas, órbitas oculares grandes em comparação com as fenestras supratemporais e o lado chanfrado do escamosal. Especificamente, eles reconheceram sua semelhança com Theriosuchus pusillus com base em seus osteodermos e dentes, embora tenham observado que a orientação da parte posterior do crânio era diferente e que os dentes em forma de folha estavam ausentes. Essas diferenças foram atribuídas a K. langenbergensis, provavelmente representando um estágio de vida diferente de T. pusillus em comparação com o espécime-tipo.[11][12] Uma análise de 2016 das relações de Atoposauridae, de Jonathan Tennant et al., apoiou provisoriamente a afinidade desses espécimes com T. pusillus devido aos dentes estarem situados em uma ranhura (uma característica que une T. pusillus e “T.” guimarotae em sua análise), à presença de dentes pseudocaniniformes e aos dentes em forma de ponta de lança perto da parte posterior da mandíbula. No entanto, eles observaram que os ossos nasais dividem essencialmente as narinas de forma completa, excluindo outros ossos, o que não é observado no gênero Theriosuchus.[12]
No entanto, a taxonomia de Theriosuchus em si é um pouco complicada. Muitos estudos taxonômicos recentes não forneceram um conjunto explícito de características que unam as espécies de Theriosuchus e as separem de outros atopossaurídeos;[13][14][15] os únicos diagnósticos desse tipo foram feitos por Steve Salisbury e Darren Naish [en] em 2011,[16] e por Jeremy Martin et al. em 2010.[17] Em 2016, Mark Young et al. criticaram esses diagnósticos, observando que muitas características eram mais amplamente distribuídas entre Atoposauridae, difíceis de avaliar ou, no caso do último diagnóstico, totalmente ausentes. Young et al. forneceram um diagnóstico alternativo contendo nove características, envolvendo principalmente os dentes; no entanto, eles também observaram que havia variação entre a expressão dessas características, o que coloca em questão a monofilia de Theriosuchus.[13] K. langenbergensis difere desse diagnóstico em quatro das nove características,[13] que Schwarz et al. citaram como base tanto para a separação genéricado Knoetschkesuchus quanto para a necessidade de revisar ainda mais o diagnóstico.[3]
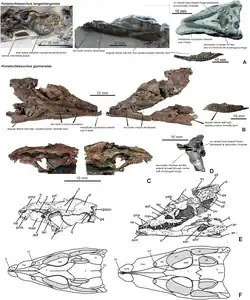
Segundo Schwarz et al., sete características unem K. langenbergensis e K. guimarotae, e as separam de outras espécies referidas a Theriosuchus. Estas incluem a presença de apenas dois morfótipos dentários únicos; as coanas estarem localizadas em sulcos rasos em vez de uma depressão em forma de tigela; o topo do crânio relativamente largo; a presença de fenestras antorbitais e mandibulares em todas as fases da vida; e o contato relativamente limitado entre o lacrimal e o nasal. Além disso, K. langenbergensis difere de outras espécies por não ter dentes com coroas baixas; ter uma sínfise maxilar mais longa; ter uma crista na lateral do processo descendente do pós-orbital; ter sobreposição entre o pós-orbital e a frente do esquamosal; e um parietal retangular que não faz parte da fenestra supratemporal.[3]
Na descrição de 2017 de K. langenbergensis, Schwarz et al. usaram o conjunto de dados filogenéticos de 2015 de Alan Turner,[18] que foi revisado para remover características irrelevantes, adicionar K. langenbergensis e Theriosuchus grandinaris, e corrigir falhas nas características codificadas de K. guimarotae (devido a imagens de baixa resolução, imprecisões na reconstrução original e aquisição de novos dados). As árvores filogenéticas recuperadas por esta análise consistentemente descobriram que a Atoposauridae, representada por Knoetschkesuchus, Theriosuchus e Alligatorium, forma um clado monofilético. Dentro deste clado, uma relação próxima entre K. langenbergensis e K. guimarotae, excluindo outras espécies de Theriosuchus, foi fortemente apoiada, fornecendo mais evidências de que essas duas espécies formam um gênero separado. Em algumas árvores, T. grandinaris também estava próximo de Knoetschkesuchus. O arranjo mais parcimonioso é reproduzido abaixo.[3]
| Eusuchia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoecologia
Sendo relativamente pequeno, Knoetschkesuchus teria se alimentado principalmente de invertebrados e possivelmente de pequenos vertebrados (incluindo anfíbios e mamíferos). A mesma dieta é observada em crocodilianos juvenis modernos da mesma classe de tamanho.[5]
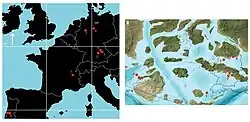
Durante o Kimmeridgiano, a Europa era coberta pelo mar interior raso do Oceano Tétis, com as massas de terra expostas sendo uma série de arquipélagos dispersos.[19][20] Níveis do mar altamente variáveis[21] sustentaram uma fauna de crocodilianos insulares altamente diversificada, com a biodiversidade (a dos atopossaurídeos em particular) sendo impulsionada fortemente pela especiação alopátrica.[22] As duas espécies de Knoetschkesuchus faziam parte de faunas semelhantes, em ambas as quais funcionavam como pequenos predadores ecologicamente particionados dos outros crocodilianos contemporâneos; é provável que essas faunas se tenham originado através da dispersão por massas de terra maiores.[3] Uma troca faunística semelhante ocorreu com a bacia de Wessex-Weald [en], na Inglaterra.[23]
Knoetschkesuchus langenbergensis
A pedreira de Langenberg, de onde K. langenbergensis é conhecido, faz parte da bacia da Baixa Saxônia, que teria feito parte da massa de terra associada aos maciços Renano, Boêmio e Londres-Brabante. Durante o Kimmeridgiano, teria sido um ambiente marinho raso; no entanto, isso não implica que K. langenbergensis fosse marinho, uma vez que os animais e plantas da pedreira de Langenberg foram provavelmente transportados aloctonamente (embora apenas por uma curta distância) das ilhas circundantes. Sedimentos de água salobra e doce também estão presentes na pedreira, o que implica que havia influxo ocasional de água doce.[3] Galhos e cones de coníferas indicam que a araucária Brachyphyllum estava presente no local.[24]
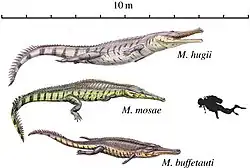
Além de K. langenbergensis, outros crocodilianos da pedreira de Langenberg incluem os marinhos Machimosaurus hugii e Steneosaurus aff. brevirostris [en],[11] que teriam vivido ao largo da costa e se alimentado de tartarugas e peixes; e o generalista anfíbio Goniopholis simus,[11] que subsistia de uma dieta de presas com e sem concha, como os aligátores modernos.[3] A pedreira é mais conhecida como a localidade-tipo do dinossauro saurópode braquiossaurídeo Europasaurus holgeri, que era um anão insular.[25] Dentes isolados mostram que havia pelo menos quatro tipos diferentes de terópodes presentes na localidade, incluindo o megalossaurídeo Torvosaurus sp. bem como um megalossaurídeo adicional e membros indeterminados de Allosauridae e Ceratosauria;[26] pegadas de terópodes de duas espécies diferentes (não identificáveis a nível de família) também são conhecidas.[1] Restos atribuíveis a diplodocídeos (possivelmente também anões) e estegossauros também foram encontrados,[24] juntamente com um pterossauro dsungaripterídeo indeterminado.[27]
Não-arcossauros também estão presentes na pedreira de Langenberg. Estes incluem um lagarto paramacelodídeo [en];[28] as tartarugas Thalassemys sp. [en],[29] Plesiochelys etalloni [en],[30] e um eucriptodiro juvenil;[31] mamíferos plagiaulacídeos multituberculados,[32] driolestídeos [en] e docodontes conhecidos a partir de dentes;[24] o mamaliaforme Storchodon [en];[33] uma fauna diversificada de peixes actinopterígios dominada por Lepidotes sp., mas que também inclui Macromesodon sp., Proscinetes sp. [en], Coelodus sp. [en], Macrosemius sp. [en] (ou Notagogus sp. [en]), Histionotus sp. [en], Ionoscopus sp. [en], Callopterus sp. [en], Caturus sp. [en], Sauropsis sp. [en], Belonostomus sp. [en] e Thrissops subovatus [en];[34] e cinco morfótipos de hibodontiformes [en], além dos neosseláquios Palaeoscyllium sp., dois distintos Synechodus sp. [en] e dois distintos Asterodermus sp. [en][35]
Knoetschkesuchus guimarotae

A localidade de Guimarota está localizada na bacia Lusitânica oriental, que faz parte da Meseta Ibérica.[3] Os estratos de Guimarota, que contêm vertebrados, foram depositados em uma lagoa salobra que recebia periodicamente influxos de água doce e salgada.[4] Ecologicamente, o ambiente teria sido semelhante a um manguezal moderno.[36][37] As plantas conhecidas a partir de megafósseis incluem as cavalinhas Equisetum e Schizoneura; samambaia com semente Caytonia [en]; a cicadácea Otozamites [en]; a araucária Brachyphyllum; Ginkgo; e a alga carófita Porochara. Além disso, as famílias de plantas conhecidas apenas pelo pólen incluem licópodes, samambaias bifurcadas, possíveis samambaias arbóreas escamosas (que podem ser matoniáceas ou dicksoniáceas), samambaias reais, a samambaia com semente Sphenopteris, ciprestes e pinheiros.[38] A maioria dos organismos preservados provavelmente se originou de ao redor do pântano, embora possa ter havido transporte mínimo por correntes de água.[37]
Muitos crocodilianos são conhecidos da mina de Guimarota. O marinho Machimosaurus hugii está novamente presente,[39] assim como Goniopholis (embora como uma espécie diferente, Goniopholis baryglyphaeus).[40] Além disso, Bernissartia sp., Lisboasaurus estesi [en] e Lusitanisuchus mitracostatus [en] também foram encontrados;[41] os dois últimos eram pequenos, com menos de 50 cm de comprimento, e provavelmente também se alimentavam de insetos como Knoetschkesuchus guimarotae.[3] Os dinossauros de Guimarota são conhecidos principalmente por dentes e incluem um braquiossaurídeo, que também é de tamanho bastante pequeno; terópodes, incluindo Stokesosaurus sp. (conhecido a partir de fósseis corporais), Compsognathus sp., alossaurídeos, um ceratossauro semelhante a Ceratosaurus, um táxon semelhante ao filogeneticamente problemático Richardoestesia, dromeossaurídeos, troodontídeos e arqueopterigiformes; e os ornitópodes Phyllodon henkeli [en] e um iguanodonte semelhante a Camptosaurus.[42] Outros vertebrados diversos, incluindo condrictes, osteíctes, anfíbios albanerpetontídeos, tartarugas, lagartos, pterossauros ranforrinquídeos e mamíferos docodontes e driolestídeos (incluindo Henkelotherium guimarotae) também estão presentes. Os invertebrados são representados por ostracodes e moluscos.[3]
Referências
- ↑ a b Lallensack, J.; Sander, P.M.; Knötschke, N.; Wings, O. (2015). «Dinosaur tracks from the Langenberg Quarry (Late Jurassic, Germany) reconstructed with historical photogrammetry: Evidence for large theropods soon after insular dwarfism». Palaeontologia Electronica. 18 (18.2.31A): 1–34. doi:10.26879/529

- ↑ Pieńkowski, G.; Schudack, M.E.; Bosák, P.; Enay, R.; Feldman-Olszewska, A.; Golonka, J.; Gutowski, J.; Herngreen, G.F.W.; Jordan, P.; Krobicki, M.; Lathuiliere, B.; Leinfelder, R.R.; Michalik, J.; Monnig, E.; Noe-Nygaard, N.; Palfy, J.; Pint, A.; Rasser, M.W.; Reisdorf, A.G.; Schmid, D.U.; Schweigert, G.; Surlyk, F.; Wetzel, A.; Wong, T.E. (2008). «Jurassic». In: McCann, T. The Geology of Central Europe Volume 2: Mesozoic and Cenozoic. Londres: Geological Society. pp. 823–922
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z Schwarz, D.; Raddatz, M.; Wings, O. (2017). «Knoetschkesuchus langenbergensis gen. nov. sp. nov., a new atoposaurid crocodyliform from the Upper Jurassic Langenberg Quarry (Lower Saxony, northwestern Germany), and its relationships to Theriosuchus». PLOS ONE. 12 (2). Bibcode:2017PLoSO..1260617S. PMC 5310792. PMID 28199316. doi:10.1371/journal.pone.0160617
- ↑ a b Schudack, M.E. (2000). «Geological setting and dating of the Guimarota beds». In: Martin, T.; Krebs, B. Guimarota – A Jurassic Ecosystem. Munique: Verlag Dr. Friedrich Pfeil. pp. 21–26. ISBN 978-3-931516-80-2
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x Schwarz, D.; Salisbury, S.W. (2005). «A new species of Theriosuchus (Atoposauridae, Crocodylomorpha) from the Late Jurassic (Kimmeridgian) of Guimarota, Portugal». Geobios. 38 (6): 779–802. doi:10.1016/j.geobios.2004.04.005
- ↑ Sousa, L. (1998). «Upper Jurassic (Upper Oxfordian–Tithonian) palynostratigraphy from the Lusitanian Basin (Portugal)». Memórias da Academia das Ciencias de Lisboa. 37: 49–77
- ↑ Schudack, M.E. (2000). «Ostracodes and charophytes from the Guimarota beds». In: Martin, T.; Krebs, B. Guimarota – A Jurassic Ecosystem. Munique: Verlag Dr. Friedrich Pfeil. pp. 33–36. ISBN 978-3-931516-80-2
- ↑ Buscaloni, A.D.; Sanz, J.L. (1988). «Phylogenetic relationships of the Atoposauridae (Archosauria, Crocodylomorpha)». Historical Biology. 1 (3): 233–250. doi:10.1080/08912968809386477
- ↑ Salisbury, S.W.; Frey, E. (2001). «A biomechanical transformation model for the evolution of semi-spheroidal articulations between adjoining vertebral bodies in crocodilians». In: Grigg, G.C.; Seebacher, F.; Franklin, C.E. Crocodilian Biology and Evolution. Chipping Norton: Surry Beatty & Sons. pp. 85–134. ISBN 978-0-949324-89-4
- ↑ Busbey, A.B. (1997). «Structural consequences of skull flattening in crocodilians». In: Thomason, J.J. Functional Morphology in Vertebrate Paleontology. Cambridge: Cambridge University Press. pp. 173–192. ISBN 978-0-521-62921-8
- ↑ a b c Karl, H.-V.; Gröning, E.; Brauckmann, C.; Schwarz, D.; Knötschke, N. (2006). «The Late Jurassic crocodiles of the Langenberg near Oker, Lower Saxony (Germany), and description of related materials (with remarks on the history of quarrying the "Langenberg Limestone" and "Obernkirchen Sandstone")». Clausthaler Geowissenschaften. 5: 59–77
- ↑ a b Tennant, J.P.; Mannion, P.D.; Upchurch, P. (2016). «Evolutionary relationships and systematics of Atoposauridae (Crocodylomorpha: Neosuchia): implications for the rise of Eusuchia» (PDF). Zoological Journal of the Linnean Society. 177 (4): 854–936. doi:10.1111/zoj.12400
- ↑ a b c Young, M.T.; Tennant, J.P.; Brusatte, S.L.; Challands, T.J.; Fraser, N.C.; Clark, N.D.L.; Ross, D.A. (2016). «The first definitive Middle Jurassic atoposaurid (Crocodylomorpha, Neosuchia), and a discussion on the genus Theriosuchus». Zoological Journal of the Linnean Society. 176 (2): 443–462. PMC 4989461. PMID 27594716. doi:10.1111/zoj.12315
- ↑ Martin, J.E.; Rabi, M.; Csiki-Sava, Z.; Vasile, S. (2014). «Cranial morphology of Theriosuchus sympiestodon (Mesoeucrocodylia, Atoposauridae) and the widespread occurrence of Theriosuchus in the Late Cretaceous of Europe». Journal of Paleontology. 88 (3): 444–456. doi:10.1666/13-106
- ↑ Lauprasert, K.; Laojumpon, C.; Saenphala, W.; Cuny, G.; Thirakhupt, K.; Suteethorn, V. (2011). «Atoposaurid crocodyliforms from the Khorat Group of Thailand: first record of Theriosuchus from Southeast Asia». Paläontologische Zeitschrift. 85 (1): 37–47. doi:10.1007/s12542-010-0071-z
- ↑ Salisbury, S.W.; Naish, D. (2011). «Crocodilians» (PDF). In: Batten, D.; Lane, P.D. English Wealden Fossils. London: Palaeontological Association. pp. 305–369. ISBN 978-1-4443-6711-9
- ↑ Martin, J.E.; Rabi, M.; Cziki, Z. (2010). «Survival of Theriosuchus (Mesoeucrocodylia: Atoposauridae) in a Late Cretaceous archipelago: a new species from the Maastrichtian of Romania». Naturwissenschaften. 97 (9): 845–854. Bibcode:2010NW.....97..845M. PMID 20711558. doi:10.1007/s00114-010-0702-y
- ↑ Turner, A.H. (2015). «A Review of Shamosuchus and Paralligator (Crocodyliformes, Neosuchia) from the Cretaceous of Asia». PLOS ONE. 10 (2). Bibcode:2015PLoSO..1018116T. PMC 4340866. PMID 25714338. doi:10.1371/journal.pone.0118116
- ↑ Fourcade, E.; Azéma, J.; Bassoullet, J.-P.; Cecca, F.; Dercourt, J.; Enay, R.; Guiraud, R. (1996). «Paleogeography and Paleoenvironment of the Tethyan Realm During the Jurassic Breakup of Pangea». In: Nairn, A.E.M.; Ricou, L.-E.; Vrielynck, B.; Dercourt, J. The Tethys Ocean. [S.l.]: Springer US. pp. 191–214. ISBN 978-1-4899-1560-3. doi:10.1007/978-1-4899-1558-0_6
- ↑ Smith, A.G.; Smith, D.G.; Funnell, B.M. (2004). Atlas of Mesozoic and Cenozoic Coastlines. Cambridge: Cambridge University Press. ISBN 978-0-521-60287-7
- ↑ Miller, K.G.; Kominz, M.A.; Browning, J.V.; Wright, J.D.; Mountain, G.S.; Katz, M.E.; Sugarman, P.J.; Cramer, B.S.; Christie-Blick, N.; Pekar, S.F. (2005). «The Phanerozoic Record of Global Sea-Level Change». Science. 310 (5752): 1293–1298. Bibcode:2005Sci...310.1293M. PMID 16311326. doi:10.1126/science.1116412
- ↑ Tennant, J.P.; Mannion, P.D. (2014). «Revision of the Late Jurassic crocodyliform Alligatorellus, and evidence for allopatric speciation driving high diversity in western European atoposaurids». PeerJ. 2. PMC 4179893. PMID 25279270. doi:10.7717/peerj.599
- ↑ Salisbury, S.W.; Willis, P.M.A.; Peitz, S.; Sander, P.M. (1999). «The crocodilian Goniopholis simus from the Lower Cretaceous of north-western Germany». Special Papers in Palaeontology. 60: 121–148
- ↑ a b c Wings, O. (2015). The Langenberg Quarry near Goslar: Unique window into a terrestrial Late Jurassic ecosystem in Northern Germany. 12th Symposium on Mesozoic Terrestrial Ecosystems. Shenyang: Paleontological Museum of Liaoning. pp. 99–100
- ↑ Sander, P.M.; Mateus, O.; Laven, T.; Knötschke, N. (2006). «Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur». Nature. 441 (7094): 739–741. Bibcode:2006Natur.441..739M. PMID 16760975. doi:10.1038/nature04633
- ↑ Gerke, O.; Wings, O. (2016). «Multivariate and Cladistic Analyses of Isolated Teeth Reveal Sympatry of Theropod Dinosaurs in the Late Jurassic of Northern Germany». PLOS ONE. 11 (7). Bibcode:2016PLoSO..1158334G. PMC 4934775. PMID 27383054. doi:10.1371/journal.pone.0158334
- ↑ Fastnacht, M. (2005). «The first dsungaripterid pterosaur from the Kimmeridgian of Germany and the biomechanics of pterosaur long bones». Acta Palaeontologica Polonica. 50 (2): 273–288
- ↑ Richter, A.; Knötschke, N.; Kosma, R.; Sobral, G.; Wings, O. (2013). The first Mesozoic lizard from northern Germany (Paramacellodidae, Late Jurassic, Langenberg Quarry) and its taphonomy. 73rd Annual Meeting of the Society of Vertebrate Paleontology. Los Angeles: Society of Vertebrate Paleontology. p. 198. Consultado em 20 de março de 2017. Cópia arquivada em 5 de março de 2016
- ↑ Marinheiro, J.; Mateus, O. (2011). Occurrence of the marine turtle Thalassemys in the Kimmeridgian of Oker, Germany (PDF). 71st Annual Meeting of the Society of Vertebrate Paleontology. Las Vegas: Society of Vertebrate Paleontology. p. 151
- ↑ Karl, H.-V. (2012). «Bite Traces in a Turtle Shell Fragment from the Kimmeridgian (Upper Jurassic) of Northern Germany». Studia Geologica Salmanticensia. 9: 25–30. ISSN 0211-8327
- ↑ Jansen, M.; Klein, N. (2014). «A juvenile turtle (Testudines, Eucryptodira) from the Upper Jurassic of Langenberg Quarry, Oker, Northern Germany». Palaeontology. 57 (4): 743–756. doi:10.1111/pala.12085
- ↑ Martin, M.; Schultz, J.A.; Schwermann, A.H.; Wings, O. «First Jurassic mammals of Germany: Multituberculate teeth from the Late Jurassic Langenberg Quarry near Goslar (Lower Saxony)» (PDF). Palaeontologia Polonica. 67: 171–179
- ↑ Martin, T.; Averianov, A.O.; Jäger, K.R.K.; Schwermann, A.H.; Wings, O. (2019). «A large morganucodontan mammaliaform from the Late Jurassic of Germany». Fossil Imprint. 75 (3–4): 504–509. doi:10.2478/if-2019-0030
- ↑ Mudroch, A.; Thies, D. (1996). «Knochenfischzähne (Osteichthyeso Actinopterygii) aus dem Oberjura (Kimmeridgium) des Langenbergs bei Oker (Norddeutschland)» [Peixes ósseos (Osteichthyes, Actinopterygii) do Jurássico Superior (Kimmeridgiano) da Pedreira de Langenberg em Oker (Norte da Alemanha)]. Geologica et Palaeontologica (em alemão). 30: 239–265
- ↑ Thies, D. (1995). «Placoid scales (Chondrichthyes: Elasmobranchii) from the Late Jurassic (Kimmeridgian) of northern Germany». Journal of Vertebrate Paleontology. 15 (3): 463–481. doi:10.1080/02724634.1995.10011242
- ↑ Martin, T. (2000). «Overview over the Guimarota ecosystem». In: Martin, T.; Krebs, B. Guimarota – A Jurassic Ecosystem. Munique: Verlag Dr. Friedrich Pfeil. pp. 143–146. ISBN 978-3-931516-80-2
- ↑ a b Gloy, G. (2000). «Taphonomy of the fossil lagerstatte Guimarota». In: Martin, T.; Krebs, B. Guimarota – A Jurassic Ecosystem. Munique: Verlag Dr. Friedrich Pfeil. pp. 129–136. ISBN 978-3-931516-80-2
- ↑ van Erve, A.; Mohr, B. (1988). «Palynological investigation of the Late Jurassic microflora from the vertebrate locality Guimarota coal mine (Leiria, Central Portugal)». Neues Jahrbuch für Geologie und Paläontologie - Monatshefte. 1988 (4): 246–262. doi:10.1127/njgpm/1988/1988/246
- ↑ Krebs, B.; Schwarz, D. (2000). «The crocodiles from the Guimarota mine». In: Martin, T.; Krebs, B. Guimarota – A Jurassic Ecosystem. Munique: Verlag Dr. Friedrich Pfeil. pp. 69–74. ISBN 978-3-931516-80-2
- ↑ Schwarz, D. (2002). «A New Species of Goniopholis from the Upper Jurassic of Portugal» (PDF). Palaeontology. 45 (1): 185–208. doi:10.1111/1475-4983.00233
- ↑ Schwarz, D.; Fechner, R. (2004). «Lusitanisuchus, a new generic name for Lisboasaurus mitracostatus (Crocodylomorpha: Mesoeucrocodylia), with a description of new remains from the Upper Jurassic (Kimmeridgian) and Lower Cretaceous (Berriasian) of Portugal». Canadian Journal of Earth Sciences. 41 (10): 1259–1271. Bibcode:2004CaJES..41.1259S. doi:10.1139/e04-059
- ↑ Rauhut, O.W.M. (2000). «The dinosaur fauna from the Guimarota mine». In: Martin, T.; Krebs, B. Guimarota – A Jurassic Ecosystem. Munique: Verlag Dr. Friedrich Pfeil. pp. 75–82. ISBN 978-3-931516-80-2
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||