Calostoma cinnabarinum
Calostoma cinnabarinum
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Nome binomial | |||||||||||||||||
| Calostoma cinnabarinum Desv. (1809)[1] | |||||||||||||||||
| Sinónimos | |||||||||||||||||
| |||||||||||||||||
Calostoma cinnabarinum
| |
|---|---|
| Himênio glebal | |
| Estipe é nua | |
|
A cor do esporo é amarelo
a castanho-escuro |
| A relação ecológica é micorrízica | |
| Comestibilidade: não comestível | |
Calostoma cinnabarinum é uma espécie de fungo gasteroide da família Sclerodermataceae. É a espécie-tipo do gênero Calostoma. Apesar da aparência, C. cinnabarinum não está relacionada aos verdadeiros cogumelos puffballs, membros do gênero Podaxis, Geastrales ou fungos fedorentos da família Phallaceae. No entanto, durante sua complexa história taxonômica, foi confundida com cada um desses grupos antes do advento da filogenética molecular.
O basidioma apresenta coloração e aparência características, com uma camada de gelatina amarelada envolvendo um píleo esférico vermelho-vivo com aproximadamente 2 cm de diâmetro, situado no topo de um estipe esponjoso vermelho ou marrom-amarelado com 1,5 a 4 cm de altura. A camada mais interna do píleo é a gleba, que contém esporos elípticos claros ou ligeiramente amarelados, medindo 14–20 μm de comprimento por 6–9 μm de largura. A superfície dos esporos apresenta um padrão de pequenas depressões, conferindo aparência reticulada. A espécie distribui-se pelas Américas e pelo leste da Ásia, onde cresce no solo em florestas decíduas, formando associações micorrízicas com carvalhos. Embora seja consumida ou utilizada na medicina tradicional em algumas regiões, geralmente é considerada não comestível.
Taxonomia

Calostoma cinnabarinum possui uma história taxonômica complexa. Leonard Plukenet ilustrou um "fungo empoeirado da Virgínia, um trabalho elegante retorcido com estipe vermelho-coral"[Note 1] em sua Phytographia de 1692,[2] posteriormente reconhecida como esta espécie.[3] Em 1809, Christiaan Persoon forneceu a primeira descrição científica moderna, como Scleroderma callostoma, e sugeriu que a espécie poderia ser suficientemente distinta para justificar a criação de um novo gênero.[4] Ainda naquele ano, Nicaise Desvaux criou o gênero Calostoma.[5] Para evitar um nome tautônimo, renomeou a espécie-tipo como C. cinnabarinum.[1]
Em 1811, Louis Bosc descreveu-a como Lycoperdon heterogeneum sem mencionar os trabalhos anteriores, embora também tenha sugerido colocá-la em gênero próprio.[6] Jean Poiret transferiu S. callostoma de Persoon para Lycoperdon em 1817, mantendo separadamente L. heterogeneum de Bosc.[7] No mesmo ano, Nees von Esenbeck observou a opinião de Bosc e criou o gênero Mitremyces, sem referenciar a atribuição anterior de Desvaux a Calostoma.[8] Um artigo de 1825 de Edward Hitchcock usou o nome binomial novo Gyropodium coccineum; embora afirmasse que o nome fora estabelecido por Lewis Schweinitz, admitiu que não havia descrição publicada,[9] sendo o nome e sua origem considerados duvidosos.[10]
Schweinitz atribuiu Lycoperdon heterogeneum de Bosc a Mitremyces como M. lutescens em 1822.[11] Uma década depois, descreveu M. cinnabarinum como espécie nova,[12] mas descrições incompletas e espécimes mal identificados geraram confusão.[13] August Corda separou-as com maior clareza, fornecendo novas descrições e atribuindo cinnabarinum a Calostoma com base em Desvaux e Persoon, mantendo lutescens em Mitremyces.[14] A monografia de George Massee de 1888 sobre Calostoma descartou a distinção, argumentando que as duas espécies de Schweinitz eram a mesma em diferentes estágios de desenvolvimento.[15] Em 1897, Charles Edward Burnap publicou nova descrição de C. lutescens, estabelecendo divisão clara entre as espécies semelhantes[13] que permanece substancialmente inalterada. Referências como "C. cinnabarina" são comuns, mas incorretas.[16]
Filogenia
| ||||||||||||||||||||||||||||||
| Cladograma mostrando a filogenia e relações de Calostoma cinnabarinum dentro de Sclerodermatineae.[17] |
As relações e origens evolutivas de Calostoma foram intensamente debatidas. Com base em características morfológicas, micologistas do século XIX a relacionaram alternadamente a Scleroderma,[18] Clathrus,[19] Geastrum,[15] ou Tulostoma.[13] A filogenética molecular no final do século XX demonstrou que Gasteromycetales era polifilética, pois fungos gasteroides não formam um clado único. Estudos de sequenciamento de DNA nuclear e mitocondrial revelaram que Calostoma cinnabarinum não é próxima de Lycoperdaceae (verdadeiros cogumelos puffballs), Phallales (fungos fedorentos), maioria dos Geastrales, nem ágaricos gasteroides como Tulostoma ou Podaxis, pertencendo à ordem Boletales.[20] Pesquisas posteriores agruparam fungos majoritariamente gasteroides, incluindo Calostoma, na subordem Sclerodermatineae. Confirmou-se que C. cinnabarinum e C. ravenelii são espécies distintas, e identificaram suas parentes mais próximas fora do gênero como Gyroporus, Astraeus [en] e Scleroderma.[21] Estudo multigênico posterior ajustou o cladograma de Sclerodermatineae, posicionando Pisolithus como parente mais próxima de Calostoma.[17]
A grande dissimilaridade morfológica de Calostoma cinnabarinum em relação a outras Boletales corresponde a taxa de deriva genética acima da média da ordem.[20] Essa característica é compartilhada por outras Sclerodermatineae, que sofreram evolução mais rápida que a ordem como um todo.[21]
Quimiotaxonomia
A inclusão do gênero Calostoma em Boletales inseriu-o em uma ordem cuja bioquímica é bem estudada. A maioria dos membros da família Boletales é caracterizada por compostos produzidos pela via do chiquimato-corismato, incluindo pigmentos característicos.[22][23][24]
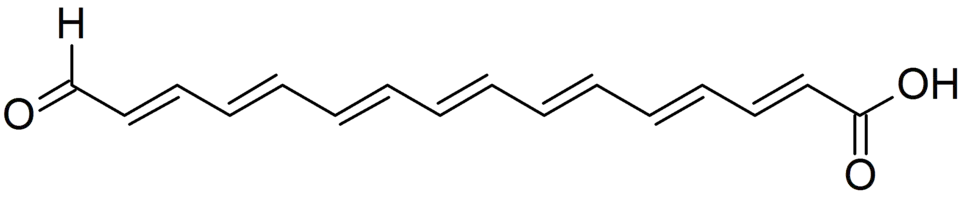
Gertraud Gruber e Wolfgang Steglich não detectaram esses compostos em C. cinnabarinum, mas isolaram um novo pigmento polieno denominado calostomal, responsável pela cor laranja-avermelhada dos basidiomas. O éster metílico do calostomal foi analisado por espectroscopia de NMR e identificado como ácido al-trans-16-oxohexadeca-2,4,6,8,10,12,14-heptaenoico.[25] Pigmentos relacionados, as boletocrocinas, foram isolados de Boletus laetissimus e B. rufoaureus.[26]
Etimologia
O epíteto específico cinnabarinum deriva do grego antigo kinnábari (κιννάβαρι), referindo-se à cor "vermelho-cinábrio", semelhante ao sangue de dragão.[27][28]
Nomes vernáculos em inglês incluem "stalked puffball-in-aspic",[16][25][29] "red slimy-stalked puffball",[30] "aspic puffball",[31] "gelatinous-stalked puffball"[27][20] e "hot lips".[27] No centro do México é chamado em espanhol de "hongo orquídea" (fungo orquídea) e em náuatle de "huang noono".[32]
Descrição

A aparência dos basidiomas foi comparada a ovos de anfíbios[33] ou "pequenos tomates vermelhos cercados por gelatina".[34] Consistem em um píleo globoso (esférico) vermelho-vivo sobre um estipe reticulado, coberto por espessa camada gelatinosa.[31] Inicialmente hipógeos,[30] emergem do solo à medida que o estipe continua a se expandir.[33]
O píleo atinge até 2 cm de diâmetro, geralmente quase esférico,[16][35] embora em algumas populações seja oval e ligeiramente menor[36] ou maior.[37] Sua estrutura interna é complexa, descrita às vezes como exoperídio e endoperídio com subcamadas,[30] outras como camadas distintas.[13] A camada mais externa é um revestimento amarelado translúcido gelatinoso de 4 a 9 mm de espessura,[37] semelhante a um véu universal gelatinoso.[13][30] Abaixo há membrana fina vermelho-cinábrio.[30][37] Com o amadurecimento, essas camadas externas se desfazem. Fragmentos da membrana vermelha incorporam-se ao gel remanescente, parecendo pequenas sementes vermelhas.[35][36] Revela-se então o endoperídio, camada resistente não gelatinosa que não se fragmenta. Inicialmente tem superfície empoeirada vermelho-viva que envelhece para laranja ou amarelo pálido.[30][37] Cristas ou raios apicais vermelho-vivos formam o peristoma. Os espécimes norte-americanos geralmente apresentam quatro a cinco dessas cristas,[30][36] enquanto populações asiáticas podem ter até sete.[37] Dentro do endoperídio está a gleba, massa de esporos branca quando jovem e fulva ou amarela em indivíduos mais velhos.[16]
O estipe também é coberto por camada gelatinosa externa[35] que consiste em filamentos gelatinosos anastomosados,[27] conferindo aspecto reticulado[16] ou esponjoso.[35] A coloração varia de vermelho a marrom-amarelado, desbotando com a idade.[30] Tem de 1 a 2 cm de espessura e 1,5 a 4 cm de comprimento, com uma parte ou o todo podendo permanecer enterrado.[16][35]
Características microscópicas
Na esporada, os esporos geralmente aparecem amarelos,[33][38] embora um população coreana com esporada rosa claro tenha sido observada.[37] Ao microscópio óptico são hialinos[27] ou amarelo-pálidos,[10] elípticos e visivelmente com depressões. Microscopia eletrônica ou de força atômica revela poros formando um retículo elaborado, com dois a três poros por micrômetro, cada um com cerca de 400 nm de profundidade.[39] A maioria mede 14–20 por 6–9 μm,[27] mas alguns chegam a 24[10] ou 28 μm;[16] espécimes coreanos apresentaram esporos ligeiramente menores. Diferentemente de outras espécies do gênero, C. cinnabarinum não utiliza células nutridoras para nutrir esporos.[39] Os basídios medem 40–50 por 15–20 μm, são amplamente obovados (formato de lágrima),[15] claviformes ou às vezes cilíndricos, com cinco a doze esporos distribuídos uniformemente[13] ou irregularmente[37] na superfície. A gleba contém hifas ramificadas de 3–4 μm de espessura com fíbulas frequentes.[13] O capilício formado por essas fíbulas [37] está presente apenas em indivíduos jovens, desintegrando-se posteriormente.[30]
Espécies semelhantes


Pelo menos na América do Norte, C. cinnabarinum é distinta e facilmente reconhecível.[16] Duas outras espécies de Calostoma também ocorrem no leste dos Estados Unidos. C. lutescens possui camada gelatinosa mais fina e camada média (mesoperídio) predominantemente amarela, com a cor vermelha restrita ao peristoma.[10] Apresenta também um colar bem definido na base do píleo,[27] estipe mais longo e esporos globosos com depressões.[16] C. ravenelii não é gelatinosa, mas possui verrugas adornando o píleo[10] e é menor que C. cinnabarinum.[27] Também possui peristoma avermelhado, mas é de cor argilosa no restante.[40] Diferentemente de C. lutescens, os esporos de C. ravenelii não podem ser distinguidos dos de C. cinnabarinum exceto pelo uso de microscopia de força atômica.[39]
Mais representantes do gênero estão presentes na Ásia. Pelo menos nove espécies foram registradas no continente indiano, algumas das quais também se sobrepõem à distribuição de C. cinnabarinum na Indonésia, Taiwan ou Japão.[41] Muitas dessas espécies podem ser distinguidas facilmente por características macroscópicas. C. japonicum é laranja-rosado e carece de camada externa gelatinosa,[37] enquanto tanto C. jiangii[42] quanto C. junghuhnii[38] são marrons. No entanto, outras exigem características microscópicas de forma e ornamentação dos esporos para identificação. Diferentemente dos esporos uniformemente alongados de C. cinnabarinum, C. guizhouense possui esporos elípticos e globosos.[42] C. pengii difere principalmente no padrão de ornamentação na superfície dos esporos.[43]
Distribuição, habitat e ecologia
Amplamente distribuída, C. cinnabarinum pode ser encontrada desde Massachusetts até a Flórida nos Estados Unidos. Sua distribuição se estende pelo menos até o Texas,[44] com populações possíveis no Sudoeste,[16] mas é mais comum nas Montanhas Apalaches, onde se torna mais frequente com o aumento da altitude.[10] Também está presente no leste do México, onde cresce nas florestas nubladas subtropicais de Veracruz[45] e Hidalgo.[46] Na América Central, é conhecido do Parque Nacional Chiquibul em Belize,[47] das florestas nubladas[48] de Baja Verapaz e El Quiché[49] na Guatemala, e do Panamá.[50] A espécie também foi registrada na América do Sul, desde a Colômbia[51] até o sudeste do Brasil, onde é descrita como rara.[52] Foi coletada ainda em uma população disjunta na Ásia, registrada em sete províncias da China continental, principalmente no sudeste,[37] incluindo Taiwan,[38] além da Indonésia,[53] Japão,[54] e Jirisan na Coreia do Sul.[39]
O cogumelo Calostoma cinnabarinum era considerado saprófito e descrito dessa forma tanto em trabalhos acadêmicos[40] quanto em discussões populares.[27] No entanto, essa classificação resultou de sua história taxonômica e comparações com fungos saprófitos não relacionados.[20] Após sua atribuição à subordem Sclerodermatineae,[21] cujos membros são ectomicorrízicos,[55][56][57] seu papel ecológico passou a ser questionado.[20] Em 2007, Andrew Wilson e David Hibbett, da Clark University, e Eric Hobbie, da Universidade de New Hampshire, empregaram marcação isotópica, sequenciamento de DNA e análise morfológica para determinar que essa espécie também é ectomicorrízica.[58] Como todos os fungos micorrízicos, C. cinnabarinum estabelece relação mutualística com as raízes de árvores, permitindo a troca de minerais e aminoácidos extraídos do solo por carbono fixado do hospedeiro. As hifas subterrâneas do fungo formam uma bainha de tecido ao redor das raízes da árvore. Essa associação é especialmente benéfica ao hospedeiro, pois o fungo produz enzimas que mineralizam compostos orgânicos e facilitam a transferência de nutrientes para a árvore.[59] As únicas árvores hospedeiras identificadas para C. cinnabarinum são carvalhos do gênero Quercus, embora membros relacionados de Calostoma tenham sido observados em associação com outras árvores da família Fagaceae, como faias.[58][60]
Além da associação obrigatória com carvalhos, C. cinnabarinum parece restrito a florestas mais úmidas.[60] Descrições iniciais de seu habitat o encontraram em "situações bastante úmidas"[13] e em "bosques úmidos",[61] e David Arora descreveu mais recentemente sua preferência pelas florestas úmidas do sul dos Apalaches.[30] Em contraste, não foi detectado nas florestas secas de carvalho da Califórnia[62][63] e provavelmente também está ausente das florestas tropicais secas do oeste da Costa Rica.[60] No Brasil, foi observado em solos arenosos e condições mais secas da Caatinga e do Cerrado, embora apenas após períodos de chuvas intensas.[52] Sua camada externa pode fornecer proteção contra dessecação.[64] Os cogumelos são mais comuns no final do verão e outono,[30][35] embora ocorrências na primavera sejam conhecidas.[27]
Esquilos foram observados se alimentando de C. cinnabarinum,[65] embora sua cobertura gelatinosa iniba a predação por insetos.[39][40]
Usos
Assim como todos os membros de seu gênero, C. cinnabarinum é geralmente considerado não comestível pelos guias de campo.[66] Como os cogumelos começam o desenvolvimento no subsolo, tornam-se duros demais para consumo quando visíveis,[30] e sua aparência pode ser considerada pouco apetitosa.[29] Um estudo sobre práticas culturais de descendentes mestiços do povo Otomi em Tenango de Doria, México, relatou que espécimes imaturos de C. cinnabarinum, conhecidos localmente como yemitas, eram frequentemente consumidos crus no passado, especialmente por crianças. O consumo da espécie não era mais comum, com apenas cinco dos 450 entrevistados familiarizados com a prática.[65] A gleba de C. cinnabarinum foi descrita como tendo sabor suave[37] e, apesar de uma lembrança local em contrário, não é doce.[65] C. cinnabarinum também foi utilizado na medicina tradicional. Um estudo etnomicológico de 1986 sobre tradições nativas em Veracruz identificou esse uso do huang noono, que os locais assavam e consumiam em pó com água mineral para tratar distúrbios gastrointestinais.[32] Diferentemente dessas tradições mexicanas, crenças populares em Hunan consideram o cogumelo venenoso devido à sua cor brilhante.[67]
Ver também
- Agrocybe putaminum
- Cyptotrama asprata
- Deconica coprophila
- Pholiota nubigena
- Protostropharia semiglobata
- Stropharia caerulea
- Roridomyces austrororidus
Notas
- ↑ Em latim: Fungus pulverulentus virginianus caudice coralline topiario opere contorto
References
- ↑ a b «Names Record: Calostoma cinnabarinum». Index Fungorum. Index Fungorum Partnership. Consultado em 14 de janeiro de 2026
- ↑ Plukenet L. (1692). Phytographia (em Latin). London: Leonard Plukenet. pl. 184
- ↑ Reed HS. (1910). «A note on two species of the genus Calostoma». The Plant World. 13 (10): 246–248. JSTOR 43476817

- ↑ Persoon CH. (1809). «Mémoire sur les vesse-loups ou Lycoperdon». Journal de Botanique (em French). 2: 5–31
- ↑ Desvaux NA. (1809). «Observations sur quelques genres à établir dans la famille des champignons». Journal de Botanique (em French e Latin). 2: 88–105
- ↑ Bosc LAG. (1811). «Mémoire sur quelques espèces de champignons des parties méridionales de l'Amérique septentrionale». Magazin der Gesellschaft Naturforschender Freunde zu Berlin für die Neuesten Entdeckungen in der Gesammten Naturkunde (em French). 5: 83–89
- ↑ Poiret JLM. (1817). «Vesse-Loup à bouche élégante». Encyclopédie Méthodique: Botanique Supplément (em French). 5: 476
- ↑ Nees von Esenbeck CDG. (1817). Das System der Pilze und Schwämme (em German). Würzburg, Germany: In der Stahelschen buchhandlung. pl. 129
- ↑ Hitchcock E. (1825). «Physiology of the Gyropodium coccineum». American Journal of Science and Arts. 9: 56–60
- ↑ a b c d e f Coker WC, Couch J (1928). The Gasteromycetes of the Eastern United States and Canada. Chapel Hill, North Carolina: University of North Carolina Press. pp. 188–193
- ↑ Schweinitz LD de. (1822). «Synopsis fungorum Carolinae Superioris». Schriften der Naturforschenden Gesellschaft zu Leipzig (em Latin). 1: 60–61
- ↑ Schweinitz LD de. (1832). «Synopsis fungorum in America Boreali media degentium». Transactions of the American Philosophical Society (em Latin). ns–4 (2): 255
- ↑ a b c d e f g h Burnap CE. (1897). «Contributions from the cryptogamic laboratory of Harvard University XXXVIII. Notes on the genus Calostoma». Botanical Gazette. 23 (3): 180–192. doi:10.1086/327486
- ↑ Corda AKJ. (1842). Anleitung Zum Studium Der Mycologie: Nebst Kritischer Beschreibung Aller Bekannten Gattungen, Und Einer Kurzen Geschichte Der Systematik (em German e Latin). Prague: Friedrich Ehrlich. pp. 97, 102
- ↑ a b c Massee G. (1888). «A monograph of the genus Calostoma, Desv. (Mitremyces, Nees)». Annals of Botany. os–2 (1): 25–45. doi:10.1093/aob/os-2.1.25
- ↑ a b c d e f g h i j Kuo M. (2011). «Calostoma cinnabarinum». MushroomExpert.com. Consultado em 14 de janeiro de 2026
- ↑ a b Binder M, Hibbett DS (2006). «Molecular systematics and biological diversification of Boletales» (PDF). Mycologia. 98 (6): 971–981. PMID 17486973. doi:10.1080/15572536.2006.11832626
- ↑ Fischer E. (1884). «Zur entwickelungsgeschichte der gastromyceten». Botanische Zeitung (em German). 42 (30). cols. 465–475
- ↑ de Bary A. (1887). Comparative Morphology and Biology of the Fungi, Mycetozoa and Bacteria. Trans. Garnsey HEF. London: Oxford University Press. pp. 312, 326
- ↑ a b c d e Hughey B, Adams GC, Bruns T, Hibbett DS (2000). «Phylogeny of Calostoma, the gelatinous-stalked puffball, based on nuclear and mitochondrial ribosomal DNA sequences». Mycologia. 92 (1): 94–104. JSTOR 3761453. doi:10.2307/3761453
- ↑ a b c Binder M, Bresinsky A (2002). «Derivation of a polymorphic lineage of Gasteromycetes from boletoid ancestors» (PDF). Mycologia. 94 (1): 85–98. JSTOR 3761848. PMID 21156480. doi:10.2307/3761848
- ↑ Prillinger H, Lopandic K, Schweigkofler W, Deak R, Aarts HJ, Bauer R, Sterflinger K, Kraus GF, Maraz A (2002). «Phylogeny and systematics of the fungi with special reference to the Ascomycota and Basidiomycota». Fungal Allergy and Pathogenicity. Col: Chemical Immunology and Allergy. 81. [S.l.]: Chemical Immunology. pp. 207–295 (see p. 269). ISBN 978-3-8055-7391-7. PMID 12102002. doi:10.1159/000058868
- ↑ Høiland K. (1987). «A new approach to the phylogeny of the order Boletales (Basidiomycotina)». Nordic Journal of Botany. 7 (6): 705–718. Bibcode:1987NorJB...7..705H. doi:10.1111/j.1756-1051.1987.tb02038.x
- ↑ Zhou Z, Liu JK (2010). «Pigments of fungi (macromycetes)». Natural Product Reports. 27 (11): 1531–1570. PMID 20694228. doi:10.1039/C004593D
- ↑ a b Gruber G, Steglich W (2007). «Calostomal, a polyene pigment from the gasteromycete Calostoma cinnabarinum (Boletales)» (PDF). Zeitschrift für Naturforschung B. 62 (1): 129–131. doi:10.1515/znb-2007-0120
- ↑ Kahner L, Dasenbrock J, Spiteller P, Steglich W, Marumoto R, Spiteller M (1998). «Polyene pigments from fruit-bodies of Boletus laetissimus and B. rufo-aureus (Basidiomycetes)». Phytochemistry. 49 (6): 1693–1697. Bibcode:1998PChem..49.1693K. PMID 11711083. doi:10.1016/S0031-9422(98)00319-7
- ↑ a b c d e f g h i j Roody WC. (2003). Mushrooms of West Virginia and the Central Appalachians. Lexington, Kentucky: University Press of Kentucky. p. 439. ISBN 978-0-8131-9039-6
- ↑ Dallas EM, Burgin CA (1900). Among the Mushrooms: A Guide for Beginners. Philadelphia, Pennsylvania: Drexel Biddle. pp. 20–21
- ↑ a b Loewer HP. (1996). Thoreau's Garden. Mechanicsburg, Pennsylvania: Stackpole Books. p. 156. ISBN 978-0-8117-1728-1
- ↑ a b c d e f g h i j k l Arora D. (1986). Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi 2nd ed. Berkeley, California: Ten Speed Press. pp. 718–19. ISBN 978-0-89815-170-1
- ↑ a b McKnight KH, McKnight VB (1998). A Field Guide to Mushrooms: North America. New York: Houghton Mifflin. p. 344. ISBN 978-0-395-91090-0
- ↑ a b Alatorre E. (1996). Etnomicologia en la Sierra de Santa Marta (PDF) (em Spanish). Xalapa, Mexico: CONABIO. pp. 26, 65
- ↑ a b c Kuo M, Methven A (2010). 100 Cool Mushrooms. Ann Arbor, Michigan: University of Michigan Press. p. 35. ISBN 978-0-472-03417-8
- ↑ Phillips R. «Calostoma cinnabarina». Rogers Mushrooms. Consultado em 14 de janeiro de 2026. Arquivado do original em 6 de novembro de 2011
- ↑ a b c d e f Bessette AE, Fischer DW, Bessette AR (1996). Mushrooms of Northeastern North America. Syracuse, New York: Syracuse University Press. p. 452. ISBN 978-0-8156-0388-7
- ↑ a b c Miller OK, Miller H (2006). North American Mushrooms: A Field Guide to Edible and Inedible Fungi. Guilford, Connecticut: Globe Pequot Press. p. 469. ISBN 978-0-7627-3109-1
- ↑ a b c d e f g h i j k Zhishu B, Guoyang Z, Taihui L (1993). The Macrofungus Flora of China's Guangdong Province. Hong Kong: The Chinese University Press. pp. 575–576. ISBN 978-962-201-556-2
- ↑ a b c Chen ZC, Yeh KY (1975). «Notes on new Formosan forest fungi III. The genus Calostoma Desv» (PDF). Taiwania. 20 (2): 229–233. doi:10.6165/tai.1975.20.229
- ↑ a b c d e Kim M, Kim KW, Jung HS (2007). «Morphological discretion of basidiospores of the puffball mushroom Calostoma by electron and atomic force microscopy» (PDF). Journal of Microbiology and Biotechnology. 17 (10): 1721–1726. PMID 18156793
- ↑ a b c Miller OK, Miller H (1988). Gasteromycetes: Morphological and Developmental Features with Keys to the Orders, Families, and Genera. Eureka, California: Mad River Press. pp. 59–60. ISBN 978-0-916422-74-5
- ↑ Fan L, Liu P, Liu YH (1994). The Gasteromycetes of China. Berlin: Lubrecht & Cramer. pp. 50–53. ISBN 978-3-443-51030-5
- ↑ a b Liu B, Jiang SZ, Liu YH (1985). «Two new Calostoma species from Guizhou». Acta Mycologica Sinica. 4 (1): 51–54
- ↑ Li LJ, Liu B, Liu YH (1984). «Two new species of the genus Calostoma from China». Acta Mycologica Sinica (em Chinese). 3 (2): 92–95
- ↑ Johnson MM. (1929). «The Gasteromycetae of Ohio: Puffballs, birds'-nest fungi and stinkhorns». Ohio Biological Survey Bulletin 22. 4 (7): 271–352 (see p. 325)
- ↑ Munguia P, Guzmán G, Ramírez-Guillén F (2006). «Seasonal community structure of macromycetes in Veracruz, Mexico». Ecography. 29 (1): 57–65. Bibcode:2006Ecogr..29...57M. doi:10.1111/j.2005.0906-7590.04252.x
- ↑ Varela L, Cifuentes J (1979). «Distribución de algunos macromicetos en el norte del estado de Hidalgo». Boletín de la Sociedad Mexicana de Micología (em Spanish). 13: 75–88
- ↑ Bridgewater S. (2012). A Natural History of Belize: Inside the Maya Forest. Austin, Texas: University of Texas Press. p. 115. ISBN 978-0-292-72671-0
- ↑ Morales O, Garciá E, Cáceres R, Bran MC, Gurriarán N, Flores R (2009). «Gasteromycetes de Guatemala: Especies citades en el período de 1948 a 2008». Revista Científica (em Spanish). 4 (1 special issue): 27–33. ISSN 2070-8246. doi:10.54495/Rev.Cientifica.EdicionEspecial2009.177
- ↑ Flores AR, Comandini O, Rinaldi AC (2012). «A preliminary checklist of macrofungi of Guatemala, with notes on edibility and traditional knowledge». Mycosphere. 3 (1): 1–21. doi:10.5943/mycosphere/3/1/1
- ↑ Gube M, Piepenbring M (2009). «Preliminary annotated checklist of Gasteromycetes in Panama». Nova Hedwigia. 89 (3–4): 519–543. Bibcode:2009NovaH..89..519G. doi:10.1127/0029-5035/2009/0089-0519
- ↑ Dumont KP, Umaña MI (1978). «Los hongos de Colombia, 5: Laternera triscapa y Calostoma cinnabarina en Colombia». Caldasia (em Spanish). 12 (58): 349–352
- ↑ a b Baseia IG, Silva BD, Leite AG, Maia LC (2007). «O gênero Calostoma (Boletales, Agaricomycetidae) em áreas de cerrado e semi-árido no Brasil» (PDF). Acta Botanica Brasilica (em Portuguese). 21 (2): 277–280. Bibcode:2007AcBBr..21..277B. doi:10.1590/S0102-33062007000200003
- ↑ Boedijn KB. (1938). «The genus Calostoma in the Netherlands Indies». Bulletin du Jardin Botanique de Buitenzorg. 16 (3): 64–75
- ↑ Castro-Mendoza E; Miller OK Jr.; Stetler DA. (1983). «Basidiospore wall ultrastructure and tissue system morphology in the genus Calostoma in North America». Mycologia. 75 (1): 36–45. JSTOR 3792921. doi:10.2307/3792921
- ↑ Godbout C, Fortin JA (1983). «Morphological features of synthesized ectomycorrhizae of Alnus crispa and A. rugosa». New Phytologist. 94 (2): 249–262. Bibcode:1983NewPh..94..249G. doi:10.1111/j.1469-8137.1983.tb04498.x
- ↑ Danielson RM. (1984). «Ectomycorrhizal associations in jack pine stands in northeastern Alberta». Canadian Journal of Botany. 62 (5): 932–939. Bibcode:1984CaJB...62..932D. doi:10.1139/b84-132
- ↑ Moyersoen B, Beever RE (2004). «Abundance and characteristics of Pisolithus ectomycorrhizas in New Zealand geothermal areas». Mycologia. 96 (6): 1225–1232. JSTOR 3762138. PMID 21148945. doi:10.2307/3762138
- ↑ a b Wilson AW, Hobbie EA, Hibbett DS (2007). «The ectomycorrhizal status of Calostoma cinnabarinum determined using isotopic, molecular, and morphological methods». Canadian Journal of Botany. 85 (4): 385–393. Bibcode:2007Botan..85..385W. doi:10.1139/B07-026
- ↑ Näsholm T, Ekblad A, Nordin A, Giesler R, Högberg M, Högberg P (1998). «Boreal forest plants take up organic nitrogen». Nature. 392 (6679): 914–916. Bibcode:1998Natur.392..914N. doi:10.1038/31921
- ↑ a b c Wilson A. (2012). «MycoDigest: "Hotlips" on the beech» (PDF). Mycena News. 63 (5): 1, 4–5
- ↑ Atkinson GF. (1911). Studies of American Fungi: Mushrooms Edible, Poisonous, Etc 3rd ed. New York: Henry Holt and Company. pp. 212–213
- ↑ Morris MH, Smith ME, Rizzo DM, Rejmánek M, Bledsoe CS (2008). «Contrasting ectomycorrhizal fungal communities on the roots of co-occurring oaks (Quercus spp.) in a California woodland». New Phytologist. 178 (1): 167–176. Bibcode:2008NewPh.178..167M. PMID 18194145. doi:10.1111/j.1469-8137.2007.02348.x
- ↑ Smith ME, Douhan GW, Rizzo DM (2007). «Ectomycorrhizal community structure in a xeric Quercus woodland based on rDNA sequence analysis of sporocarps and pooled roots». New Phytologist. 174 (4): 847–863. Bibcode:2007NewPh.174..847S. PMID 17504467. doi:10.1111/j.1469-8137.2007.02040.x
- ↑ Roberts P, Evans S (2011). The Book of Fungi. Chicago, Illinois: University of Chicago Press. p. 511. ISBN 978-0-226-72117-0
- ↑ a b c Bautista-Nava E, Moreno-Fuentes A (2009). «Primer registro de Calostoma cinnabarina (Sclerodermatales) como especie comestible» (PDF). Revista Mexicana de Biodiversidad (em Spanish). 80 (2): 561–564. doi:10.22201/ib.20078706e.2009.002.629. Consultado em 14 de janeiro de 2026. Arquivado do original (PDF) em 16 de julho de 2011
- ↑ Rubel W, Arora D (2008). «A study of cultural bias in field guide determinations of mushroom edibility using the iconic mushroom, Amanita muscaria, as an example» (PDF). Economic Botany. 62 (3): 223–243. Bibcode:2008EcBot..62..223R. doi:10.1007/s12231-008-9040-9. Arquivado do original (PDF) em 15 de abril de 2012
- ↑ Härkönen M. (2002). «Mushroom Collecting in Tanzania and Hunan (Southern China): Inherited Wisdom and Folklore of Two Different Cultures». In: Watling R, Frankland JC, Ainsworth AM, Isaac S, Robinson CH. Tropical Mycology: Volume 1: Macromycetes. Wallingford, United Kingdom: CAB International. pp. 149–166. ISBN 978-0-85199-542-7