Barreira de Weismann
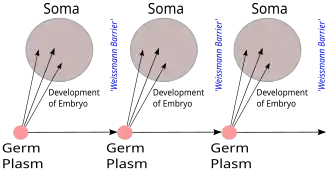
A barreira de Weismann, proposta por August Weismann, é a distinção estrita entre as linhagens de células germinativas "imortais" que produzem gametas e as células somáticas "descartáveis" nos animais (mas não nas plantas), em contraste com o mecanismo de pangênese proposto por Charles Darwin para a herança.[1][2]
Em terminologia mais precisa, a informação hereditária é copiada apenas das células da linhagem germinativa para as células somáticas. Isso significa que novas informações de mutações somáticas não são passadas para a linhagem germinativa. Este conceito de barreira implica que as mutações somáticas não são herdadas.[3][4]
Weismann expôs o conceito em seu livro de 1892, ″Das Keimplasma: eine Theorie der Vererbung″ (em alemão, O Plasma Germinativo: uma teoria da herança).[5] O uso desta teoria, comumente no contexto da teoria do plasma germinativo do final do século XIX, antes do desenvolvimento de conceitos genéticos mais bem fundamentados e sofisticados no início do século XX, é por vezes referido como Weismannismo.[6] Alguns autores distinguem o desenvolvimento Weismannista (seja preformista ou epigenético), no qual existe uma linhagem germinativa distinta, da embriogênese somática [en].[7] Este tipo de desenvolvimento está correlacionado com a evolução da morte [en] da linha somática.
A barreira de Weismann foi de grande importância em sua época e, entre outras influências, baniu efetivamente certos conceitos lamarckistas: em particular, tornaria a herança lamarckista de mudanças no corpo (o soma) difícil ou impossível.[8] Ela permanece importante, mas, no entanto, exigiu qualificação à luz da compreensão moderna da transferência horizontal de genes e de alguns outros desenvolvimentos genéticos e histológicos.[9]
Imortalidade da linhagem germinativa
O biólogo e historiador russo Zhores A. Medvedev, ao rever a teoria de Weismann um século depois, considerou que a precisão dos sistemas replicativos do genoma e outros sistemas sintéticos por si só não poderiam explicar as linhagens de células germinativas "imortais" propostas por Weismann. Em vez disso, Medvedev pensou que as características conhecidas da bioquímica e da genética da reprodução sexuada indicavam a presença de processos únicos de manutenção e restauração da informação nas diferentes fases da gametogênese. Em particular, Medvedev considerou que as oportunidades mais importantes para a manutenção da informação das células germinativas são criadas pela recombinação durante a meiose e pelo reparo do DNA; ele via estes como processos dentro das células germinativas que eram capazes de restaurar a integridade do DNA e dos cromossomos dos tipos de danos que causavam envelhecimento irreversível nas células somáticas.[10]
Animais basais
Animais basais como esponjas (Porifera) e corais (Anthozoa) contêm linhagens de células-tronco multipotentes, que dão origem tanto a células somáticas quanto a células reprodutivas. A barreira de Weismann parece ser de uma origem evolutiva mais recente entre os animais.[11]
Plantas
Nas plantas, as alterações genéticas nas linhas somáticas podem e resultam em alterações genéticas nas linhas germinativas, porque as células germinativas são produzidas por linhagens de células somáticas (meristemas vegetativos), que podem ser suficientemente antigas para terem acumulado múltiplas mutações desde a germinação da semente, algumas delas sujeitas à seleção natural.[12] É digno de nota neste contexto que, de um modo geral, as plantas adultas e em reprodução tendem a produzir um número muito maior de descendentes do que os organismos animais.
Ver também
- Alternativas à evolução Darwiniana - Lista de alternativas à seleção natural darwiniana
- Efeito Baldwin - Efeito do comportamento aprendido na evolução
- Lamarquismo - Hipótese científica sobre hereditariedade
- Pangênese - O mecanismo proposto por Darwin para a hereditariedade
Referências
- ↑ Geison, G. L. (1969). «Darwin and heredity: The evolution of his hypothesis of pangenesis». J Hist Med Allied Sci. XXIV (4): 375–411. PMID 4908353. doi:10.1093/jhmas/XXIV.4.375
- ↑ You, Yawen (26 de Janeiro de 2015). «The Germ-Plasm: a Theory of Heredity (1893), by August Weismann». The Embryo Project Encyclopedia (Arizona State University). Consultado em 7 de Setembro de 2018
- ↑ Gauthier, Peter (Março–Maio de 1990). «Does Weismann's Experiment Constitute a Refutation of the Lamarckian Hypothesis?». BIOS. 61 (1/2): 6–8. JSTOR 4608123
- ↑ Isso não se refere ao dogma central da biologia molecular, que afirma que nenhuma informação sequencial pode viajar da proteína para o DNA ou RNA, mas ambas as hipóteses se relacionam com uma visão da vida centrada nos genes.. De Tiege, Alexis; Tanghe, Koen; Braeckman, Johan; Van de Peer, Yves (Janeiro de 2014). «From DNA- to NA-centrism and the conditions for gene-centrism revisited». Biology & Philosophy. 29 (1): 55–69. doi:10.1007/s10539-013-9393-z
- ↑ Weismann, August (1892). Das Keimplasma: eine Theorie der Vererbung. Jena: Fischer
- ↑ Romanes, George John (1893). An examination of Weismannism. Chicago: Open court. OL 23380098M
- ↑ Ridley, Mark (2004). Evolution 3rd ed. [S.l.]: Blackwell. pp. 295–297
- ↑ Romanes, George John (1893). An examination of Weismannism. [S.l.]: Open Court. OL 23380098M
- ↑ Lindley, Robyn A. «How Mutational and Epigenetic Changes Enable Adaptive Evolution». G. I. T. Laboratory Journal
- ↑ Medvedev, Zhores A. (1981). «On the immortality of the germ line: Genetic and biochemical mechanisms. A review». Mechanisms of Ageing and Development. 17 (4): 331–359. ISSN 0047-6374. PMID 6173551. doi:10.1016/0047-6374(81)90052-X
- ↑ Radzvilavicius, Arunas L.; Hadjivasiliou, Zena; Pomiankowski, Andrew; Lane, Nick (20 de dezembro de 2016). «Selection for Mitochondrial Quality Drives Evolution of the Germline». PLOS Biology. 14 (12): e2000410. ISSN 1545-7885. PMC 5172535
 . PMID 27997535. doi:10.1371/journal.pbio.2000410
. PMID 27997535. doi:10.1371/journal.pbio.2000410
- ↑ Whitham, T.G.; Slobodchikoff, C.N. (1981). «Evolution by individuals, plant-herbivore interactions, and mosaics of genetic variability: The adaptive significance of somatic mutations in plants». Oecologia. 49 (3): 287–292. Bibcode:1981Oecol..49..287W. PMID 28309985. doi:10.1007/BF00347587