Alternativas à evolução Darwiniana

Alternativas à evolução darwiniana foram propostas por estudiosos que investigam a biologia para explicar evidências de evolução e a relação entre diferentes grupos de seres vivos. Essas alternativas não negam que mudanças evolutivas ao longo do tempo sejam a origem da diversidade da vida, nem que os organismos vivos hoje compartilhem um ancestral comum de um passado distante (ou ancestrais, em algumas propostas); em vez disso, propõem mecanismos alternativos de mudança evolutiva ao longo do tempo, argumentando contra mutações influenciadas pela seleção natural como o principal motor da evolução.
Isso as distingue de certos tipos de argumentos que negam que a evolução em larga escala tenha ocorrido, como em algumas formas de criacionismo, que não propõem mecanismos alternativos de mudança evolutiva, mas negam completamente que tal mudança tenha acontecido. Nem todas as formas de criacionismo negam a ocorrência de mudanças evolutivas; notavelmente, proponentes da evolução teísta, como o biólogo Asa Gray, afirmam que a mudança evolutiva ocorre e é responsável pela história da vida na Terra, com a ressalva de que esse processo foi influenciado por um deus ou deuses de maneira significativa.
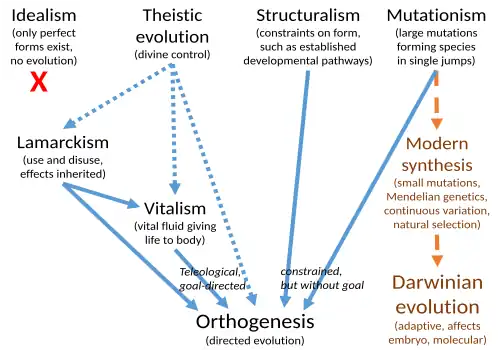
Apesar da ocorrência de mudanças evolutivas ter sido aceita, o mecanismo proposto por Charles Darwin, a seleção natural, foi negado e explicações de evolução como lamarckismo, catastrofismo, ortogênese, vitalismo, estruturalismo e mutacionismo (chamado saltacionismo antes de 1900) foram consideradas. Diferentes fatores motivaram pessoas a propor mecanismos evolutivos não darwinistas. A seleção natural, com sua ênfase em morte e competição, não atraía alguns naturalistas por considerarem-na imoral, deixando pouco espaço para a teleologia ou o conceito de progresso (ortogênese) no desenvolvimento da vida. Alguns que aceitaram a evolução, mas rejeitaram a seleção natural, levantaram objeções religiosas. Outros acreditavam que a evolução era um processo inerentemente progressivo que a seleção natural sozinha não podia explicar. Outros ainda sentiam que a natureza, incluindo o desenvolvimento da vida, seguia padrões ordenados que a seleção natural não explicava.
No início do século XX, a evolução era geralmente aceita pelos biólogos, mas a seleção natural estava em declínio.[2] Muitas teorias alternativas foram propostas, mas os biólogos rapidamente descartaram teorias como ortogênese, vitalismo e lamarckismo, que não ofereciam mecanismos para a evolução. O mutacionismo propôs um mecanismo, mas não foi amplamente aceito. A síntese moderna, uma geração depois, afirmou ter eliminado todas as alternativas à evolução darwiniana, embora algumas tenham sido revividas à medida que mecanismos moleculares para elas foram descobertos.
Formas imutáveis
Aristóteles não abraçou nem a criação divina nem a evolução, argumentando em sua biologia que cada espécie (eidos) era imutável, reproduzindo-se fielmente à sua forma eterna ideal (não idêntica à Teoria das Formas de Platão).[3][4] A sugestão de Aristóteles em Da Geração dos Animais de uma hierarquia fixa na natureza – uma scala naturae ("escada da natureza") – forneceu uma explicação inicial para a continuidade dos seres vivos.[5][6][7] Aristóteles observou que os animais eram teleológicos (direcionados funcionalmente ao propósito) e tinham partes homólogas com as de outros animais, mas não conectou essas ideias a um conceito de progresso evolutivo.[8]
Na Idade Média, a escolástica desenvolveu a visão de Aristóteles na ideia de uma grande cadeia do ser.[1] A imagem de uma escada sugere inerentemente a possibilidade de ascensão, mas tanto os gregos antigos quanto os escolásticos medievais, como Ramon Llull,[1] sustentavam que cada espécie permanecia fixa desde o momento de sua criação.[9][8]
Em 1818, no entanto, Étienne Geoffroy Saint-Hilaire argumentou em sua Philosophie anatomique que a cadeia era "uma série progressiva", onde animais como moluscos, situados na base da cadeia, poderiam "ascender, pela adição de partes, da simplicidade das primeiras formações à complexidade das criaturas no topo da escala", dado tempo suficiente. Assim, Geoffroy e biólogos posteriores buscaram explicações para tal mudança evolutiva.[10]
O Recherches sur les Ossements Fossiles de Georges Cuvier em 1812 estabeleceu sua doutrina da correlação de partes, ou seja, que, como um organismo era um sistema completo, todas as suas partes se correspondiam mutuamente, contribuindo para a função do todo. Assim, a partir de um único osso, o zoólogo frequentemente poderia identificar a classe ou até o gênero do animal. Se um animal tinha dentes adaptados para cortar carne, o zoólogo poderia ter certeza, sem mesmo olhar, que seus órgãos sensoriais seriam os de um predador e seus intestinos os de um carnívoro. Uma espécie tinha uma complexidade funcional irredutível, e "nenhuma de suas partes pode mudar sem que as outras também mudem".[11] Os evolucionistas esperavam que uma parte mudasse de cada vez, uma mudança seguindo a outra. Na visão de Cuvier, a evolução era impossível, pois qualquer mudança desequilibraria todo o sistema delicado.[11]
O Ensaio sobre Classificação de Louis Agassiz em 1856 exemplificou o idealismo filosófico alemão. Ele sustentava que cada espécie era complexa em si mesma, tinha relações complexas com outros organismos e se encaixava precisamente em seu ambiente, como uma árvore de pinheiro em uma floresta, e não poderia sobreviver fora desses círculos. O argumento a partir de formas ideais opunha-se à evolução sem oferecer um mecanismo alternativo real. Richard Owen tinha uma visão semelhante na Grã-Bretanha.[12]
O filósofo social lamarckiano e evolucionista Herbert Spencer, ironicamente o autor da frase "sobrevivência do mais apto" adotada por Darwin,[13] usou um argumento semelhante ao de Cuvier para se opor à seleção natural. Em 1893, ele afirmou que uma mudança em qualquer estrutura do corpo exigiria que todas as outras partes se adaptassem para se encaixar no novo arranjo. A partir disso, ele argumentou que era improvável que todas as mudanças aparecessem no momento certo se cada uma dependesse de variação aleatória; enquanto em um mundo lamarckiano, todas as partes se adaptariam naturalmente ao mesmo tempo, por meio de um padrão alterado de uso e desuso.[14]
Explicações alternativas de mudança
Onde a ocorrência de mudanças evolutivas foi aceita pelos biólogos, mas a seleção natural foi negada, incluindo, mas não se limitando ao final do século XIX durante a eclipse do darwinismo, explicações científicas alternativas como lamarckismo, ortogênese, estruturalismo, catastrofismo, vitalismo e evolução teísta[a] foram consideradas, não necessariamente de forma separada. Diferentes fatores motivaram pessoas a propor mecanismos evolutivos não darwinianos. A seleção natural, com sua ênfase em morte e competição, não atraía alguns naturalistas por considerarem-na imoral, deixando pouco espaço para a teleologia ou o conceito de progresso no desenvolvimento da vida.[15][16] Alguns desses cientistas e filósofos, como George Jackson Mivart e Charles Lyell, que vieram a aceitar a evolução, mas rejeitavam a seleção natural, levantaram objeções religiosas.[17] Outros, como o biólogo e filósofo Herbert Spencer, o botânico George Henslow (filho do mentor de Darwin, John Stevens Henslow, também botânico), e o autor Samuel Butler, acreditavam que a evolução era um processo inerentemente progressivo que a seleção natural sozinha não podia explicar. Outros ainda, incluindo os paleontólogos americanos Edward Drinker Cope e Alpheus Hyatt, tinham uma perspectiva idealista e acreditavam que a natureza, incluindo o desenvolvimento da vida, seguia padrões ordenados que a seleção natural não explicava.[18]
Alguns consideravam que a seleção natural seria muito lenta, dados os cálculos da idade da Terra e do Sol (10–100 milhões de anos) feitos na época por físicos como Lorde Kelvin, e alguns acreditavam que a seleção natural não poderia funcionar porque, na época, os modelos de herança envolviam a mistura de características herdadas, uma objeção levantada pelo engenheiro Fleeming Jenkin em uma revisão do A Origem das Espécies escrita logo após sua publicação.[18][19] Outro fator no final do século XIX foi o surgimento de uma nova facção de biólogos, tipificada por geneticistas como Hugo de Vries e Thomas Hunt Morgan, que desejavam reformular a biologia como uma ciência experimental de laboratório. Eles desconfiavam do trabalho de naturalistas como Darwin e Alfred Russel Wallace, dependente de observações de campo de variação, adaptação e biogeografia, por considerá-lo excessivamente anedótico. Em vez disso, focavam em tópicos como fisiologia e genética que poderiam ser investigados com experimentos controlados em laboratório, e desconsideravam fenômenos menos acessíveis como a seleção natural e a adaptação ao ambiente.[20]
| Teoria | Data | Proponente notável |
Espécies podem mudar? |
Mecanismo de mudança |
Mecanismo é físico? |
Extinção possível? |
Notas |
|---|---|---|---|---|---|---|---|
| Scala naturae[6] | c. 350 a.C. | Aristóteles | Não | Nenhum | N/A | Não | Características dos grupos não se encaixam em uma escala linear, como Aristóteles observou.[6] Teleologia e homologia reconhecidas, mas não conectadas como evolução com adaptação; não espiritual |
| Grande cadeia do ser[1] | 1305 | Llull, Ramon; escolásticos |
Não | Nenhum | N/A | Não | Aristotélico, integrado à teologia cristã |
| Vitalismo[21] | 1759 | Wolff, Caspar Friedrich | Sim | Uma força vital no embrião | Não | Não? | Variedades da teoria desde o Antigo Egito, frequentemente espirituais. Abandonada da biologia com a síntese química de moléculas orgânicas, como a da ureia em 1828 |
| Evolução teísta | 1871–6 | Gray, Asa Mivart, St George J. |
Sim | Divindade fornece mutações benéficas (Gray 1876), ou define direção (ortogenética) (Mivart 1871) | Não | Sim | "Falhou no teste do naturalismo metodológico que passou a definir a ciência".[22] Descartada por biólogos até 1900[23] |
| Ortogênese[24] | 1859 | Baer, Karl von | Sim | "Criação proposital" | Não | Sim? | Muitas variantes nos séculos XIX e XX |
| Ortogênese[25] incl. evolução emergente |
1959 | Teilhard de Chardin, Pierre | Sim | "Tendência progressiva inerente" (teleológica, vitalista) | Não | Sim | Teoria espiritual, emergência da mente, Ponto Ômega |
| Lamarckismo[26] | 1809 | Lamarck, Jean-Baptiste | Sim | Uso e desuso; herança de características adquiridas | Pensava-se que sim, mas nenhum foi encontrado | Não | Parte de sua visão de ortogênese. Abandonado da biologia, pois a barreira de Weismann impede que mudanças nas células somáticas afetem a linha germinativa nas gônadas |
| Catastrofismo[27] | 1812 | Cuvier, Georges | Não | Extinções causadas por eventos naturais como vulcanismo, inundações | Sim, para reduzir o número de espécies | Sim | Para explicar extinções e sucessão faunística de tetrápodes no registro fóssil; repovoamento por novas espécies após tais eventos notado, mas não explicado |
| Estruturalismo[28] | 1917 | Thompson, D'Arcy | Sim | Auto-organização, forças físicas | Sim | Sim? | Muitas variantes, algumas influenciadas por vitalismo |
| Saltacionismo[29][30] ou Mutacionismo |
1831 | Geoffroy Saint-Hilaire, Étienne | Sim | Grandes mutações | Sim | Sim? | Produção súbita de novas espécies sob pressão ambiental |
| Teoria neutralista da evolução molecular[31] | 1968 | Kimura, Motoo | Sim | Deriva genética | Sim | Sim | Apenas no nível molecular; compatível com a seleção natural em níveis superiores. O 'relógio molecular' observado apoia a deriva neutra; não é rival da seleção natural, pois não causa evolução do fenótipo |
| Evolução darwiniana[32] |
1859 | Darwin, Charles | Sim | Seleção natural | Sim | Sim | Faltavam mecanismos de mutação e hereditariedade até o nascimento da genética, 1900; Darwin propôs em vez disso pangênese e certo grau de herança de características adquiridas |
Vitalismo

O vitalismo sustenta que os organismos vivos diferem de outras coisas por conterem algo não físico, como um fluido ou espírito vital, que os faz viver.[33] A teoria remonta ao Antigo Egito.[34][21] Desde os tempos modernos iniciais, o vitalismo contrastava com a explicação mecanicista de sistemas biológicos iniciada por Descartes. Químicos do século XIX propuseram-se a refutar a alegação de que a formação de compostos orgânicos exigia influência vitalista.[33] Em 1828, Friedrich Wöhler demonstrou que a ureia poderia ser produzida inteiramente a partir de compostos inorgânicos.[35] Louis Pasteur acreditava que a fermentação exigia organismos inteiros, que ele supunha realizarem reações químicas exclusivas de seres vivos. O embriologista Hans Driesch, experimentando com ovos de ouriço-do-mar, mostrou que separar as duas primeiras células resultava em duas blástulas completas, mas menores, aparentemente indicando que a divisão celular não dividia o ovo em sub-mecanismos, mas criava mais células, cada uma com a capacidade vital de formar um novo organismo. O vitalismo foi gradualmente abandonado com a demonstração de explicações mecanicistas mais satisfatórias para cada uma das funções de uma célula ou organismo vivo.[33][36] Até 1931, os biólogos haviam "quase unanimemente abandonado o vitalismo como uma crença reconhecida."[37]
Evolução teísta
O botânico americano Asa Gray usou o termo "evolução teísta"[b] para seu ponto de vista, apresentado em seu livro de 1876 Essays and Reviews Pertaining to Darwinism.[38] Ele argumentou que a divindade fornece mutações benéficas para guiar a evolução. St George Jackson Mivart argumentou, em seu livro de 1871 On the Genesis of Species, que a divindade, equipada com presciência, define a direção da evolução ao especificar as leis (ortogenéticas) que a governam, deixando as espécies evoluírem de acordo com as condições que experimentam com o passar do tempo. O Duque de Argyll apresentou visões semelhantes em seu livro de 1867 The Reign of Law.[23][39] Segundo o historiador Edward Larson, a teoria falhou como explicação aos olhos dos biólogos do final do século XIX, pois violava as regras do naturalismo metodológico que eles passaram a esperar.[22] Assim, por volta de 1900, os biólogos não viam mais a evolução teísta como uma teoria válida. Na visão de Larson, ela "nem mesmo merecia um aceno entre os cientistas" naquela época.[23] No século XX, a evolução teísta poderia assumir outras formas, como a ortogênese de Teilhard de Chardin.[40]
Ortogênese
A ortogênese ou progressionismo é a hipótese de que a vida tem uma tendência inata de mudar, desenvolvendo-se de maneira unilinear em uma direção particular ou simplesmente progredindo de forma definida. Muitas versões diferentes foram propostas, algumas, como a de Teilhard de Chardin, abertamente espirituais, outras, como a de Theodor Eimer, aparentemente apenas biológicas. Essas teorias frequentemente combinavam ortogênese com outros supostos mecanismos. Por exemplo, Eimer acreditava na evolução lamarckiana, mas sentia que leis internas de crescimento determinavam quais características seriam adquiridas e guiariam a direção de longo prazo da evolução.[41][42]
A ortogênese foi popular entre paleontólogos como Henry Fairfield Osborn. Eles acreditavam que o registro fóssil mostrava uma mudança unidirecional, mas não aceitavam necessariamente que o mecanismo que impulsionava a ortogênese era teleológico (direcionado a um objetivo). Osborn argumentou em seu livro de 1918 Origin and Evolution of Life que as tendências nos chifres dos Titanothere eram tanto ortogenéticas quanto não adaptativas, e poderiam ser prejudiciais ao organismo. Por exemplo, supunham que os grandes chifres do alce irlandês causaram sua extinção.[41][42]
O apoio à ortogênese diminuiu durante a síntese moderna na década de 1940, quando ficou evidente que ela não podia explicar os complexos padrões ramificados da evolução revelados por análises estatísticas do registro fóssil.[18][19] Trabalhos no século XXI apoiaram o mecanismo e a existência de adaptação enviesada por mutação (uma forma de mutacionismo), significando que a ortogênese restrita é agora vista como possível.[43][44][45] Além disso, os processos de auto-organização envolvidos em certos aspectos do desenvolvimento embrionário frequentemente exibem resultados morfológicos estereotipados, sugerindo que a evolução procederá em direções preferenciais uma vez que componentes moleculares-chave estejam presentes.[46]
Lamarckismo

A teoria evolutiva de Jean-Baptiste Lamarck de 1809, transmutação de espécies, baseava-se em um impulso progressivo (ortogenético) para maior complexidade. Lamarck também compartilhava a crença, comum na época, de que características adquiridas durante a vida de um organismo poderiam ser herdadas pela próxima geração, produzindo adaptação ao ambiente. Essas características eram causadas pelo uso ou desuso da parte do corpo afetada. Esse componente menor da teoria de Lamarck tornou-se conhecido, muito mais tarde, como lamarckismo.[26] Darwin incluiu Efeitos do Uso e Desuso Aumentado de Partes, como Controlado pela Seleção Natural em A Origem das Espécies, dando exemplos como aves terrestres grandes que desenvolvem pernas mais fortes pelo exercício e asas mais fracas por não voarem, até que, como o avestruz, não conseguissem voar de todo.[47] No final do século XIX, o neo-lamarckismo foi apoiado pelo biólogo alemão Ernst Haeckel, os paleontólogos americanos Edward Drinker Cope e Alpheus Hyatt, e o entomologista americano Alpheus Packard. Butler e Cope acreditavam que isso permitia que os organismos conduzissem efetivamente sua própria evolução.[48] Packard argumentou que a perda de visão em insetos cegos de cavernas que ele estudou era melhor explicada por um processo lamarckiano de atrofia por desuso combinado com a herança de características adquiridas.[48][49][50] Enquanto isso, o botânico inglês George Henslow estudou como o estresse ambiental afetava o desenvolvimento das plantas, e escreveu que as variações induzidas por tais fatores ambientais poderiam explicar amplamente a evolução; ele não via necessidade de demonstrar que tais variações poderiam ser herdadas.[51] Críticos apontaram que não havia evidências sólidas para a herança de características adquiridas. Em vez disso, o trabalho experimental do biólogo alemão August Weismann resultou na teoria do plasma germinativo da herança, que Weismann dizia tornar impossível a herança de características adquiridas, uma vez que a barreira de Weismann impediria que mudanças ocorridas no corpo após o nascimento fossem herdadas pela próxima geração.[49][52]
Na epigenética moderna, os biólogos observam que os fenótipos dependem de mudanças herdáveis na expressão gênica que não envolvem alterações na sequência de DNA. Essas mudanças podem atravessar gerações em plantas, animais e procariotos. Isso não é idêntico ao lamarckismo tradicional, pois as mudanças não duram indefinidamente e não afetam a linha germinativa e, portanto, a evolução dos genes.[53]
.jpg)
Catastrofismo
O catastrofismo é a hipótese, defendida pelo anatomista e paleontólogo francês Georges Cuvier em seu Recherches sur les ossements fossiles de quadrupèdes de 1812, de que as várias extinções e os padrões de sucessão faunística vistos no fóssil foram causados por catástrofes naturais em grande escala, como erupções vulcânicas e, para as extinções mais recentes na Eurásia, a inundação de áreas de baixa altitude pelo mar. Isso foi explicado puramente por eventos naturais: ele não mencionou o Dilúvio de Noé,[54] nem se referiu à criação divina como o mecanismo para o repovoamento após um evento de extinção, embora também não apoiasse teorias evolutivas como as de seus contemporâneos Lamarck e Geoffroy Saint-Hilaire.[55][56] Cuvier acreditava que o registro estratigráfico indicava que houve várias dessas catástrofes, eventos naturais recorrentes, separados por longos períodos de estabilidade durante a história da vida na Terra. Isso o levou a acreditar que a Terra tinha vários milhões de anos.[57]
O catastrofismo encontrou um lugar na biologia moderna com o evento de extinção do Cretáceo-Paleógeno no final do período Cretáceo, conforme proposto em um artigo de Walter e Luis Alvarez em 1980. Ele argumentou que um asteroide de 10 km atingiu a Terra há 66 milhões de anos no final do período Cretáceo. O evento, qualquer que tenha sido, causou a extinção de cerca de 70% de todas as espécies, incluindo os dinossauros, deixando para trás a fronteira Cretáceo-Paleógeno.[58] Em 1990, uma cratera candidata de 180 km marcando o impacto foi identificada em Chicxulub na península de Yucatán do México.[59]
Estruturalismo
O estruturalismo biológico se opõe a uma explicação exclusivamente darwiniana da seleção natural, argumentando que outros mecanismos também guiam a evolução, e às vezes implicando que esses superam completamente a seleção.[28] Os estruturalistas propuseram diferentes mecanismos que poderiam ter guiado a formação de planos corporais. Antes de Darwin, Étienne Geoffroy Saint-Hilaire argumentou que os animais compartilhavam partes homólogas, e que se uma fosse aumentada, as outras seriam reduzidas em compensação.[60] Após Darwin, D'Arcy Thompson sugeriu vitalismo e ofereceu explicações geométricas em seu clássico livro de 1917 On Growth and Form.[28] Adolf Seilacher sugeriu inflação mecânica para estruturas "pneu" em fósseis da biota ediacarana como Dickinsonia.[61][62] Günter P. Wagner defendeu o viés de desenvolvimento, restrições estruturais no desenvolvimento embrionário.[63][64] Stuart Kauffman favoreceu a auto-organização, a ideia de que uma estrutura complexa emerge holisticamente e espontaneamente da interação dinâmica de todas as partes de um organismo.[65] Michael Denton defendeu leis da forma pelas quais universais platônicos ou "Tipos" são auto-organizados.[66] Em 1979, Stephen J. Gould e Richard Lewontin propuseram enjuntas biológicas, características criadas como subproduto da adaptação de estruturas próximas.[63] Gerd Müller e Stuart Newman argumentaram que o aparecimento no registro fóssil da maioria dos filos atuais na explosão cambriana foi uma evolução "pré-mendeliana" causada por respostas plásticas de sistemas morfogenéticos que eram parcialmente organizados por mecanismos físicos.[67][68] Brian Goodwin, descrito por Wagner como parte de "um movimento fringe em biologia evolutiva",[63] negou que a complexidade biológica pudesse ser reduzida à seleção natural e argumentou que a formação de padrões é impulsionada por campos morfogenéticos.[69] Biólogos darwinistas criticaram o estruturalismo, enfatizando que há evidências abundantes de homologia profunda de que genes estiveram envolvidos na formação dos organismos ao longo da história evolutiva. Eles aceitam que algumas estruturas, como a membrana celular, se auto-organizam, mas questionam a capacidade da auto-organização de impulsionar a evolução em grande escala.[70][71]
Saltacionismo e Mutacionismo
%252C_by_Th%C3%A9r%C3%A8se_Schwartze_(1851-1918).jpg)
O saltacionismo[72][73] sustentava que novas espécies surgem como resultado de grandes mutações. Era visto como uma alternativa muito mais rápida ao conceito darwiniano de um processo gradual de pequenas variações aleatórias sendo influenciadas pela seleção natural. Era popular entre os primeiros geneticistas, como Hugo de Vries, que, junto com Carl Correns, ajudou a redescobrir as leis de herança de Gregor Mendel em 1900, William Bateson, um zoólogo britânico que mudou para a genética, e, no início de sua carreira, Thomas Hunt Morgan. Essas ideias desenvolveram-se no mutacionismo, a teoria da mutação da evolução.[29][30] Isso sustentava que as espécies passavam por períodos de mutação rápida, possivelmente como resultado de estresse ambiental, que poderiam produzir múltiplas mutações e, em alguns casos, espécies completamente novas em uma única geração, com base nos experimentos de Hugo com a primavera-noturna, Oenothera, a partir de 1886. As primaveras pareciam estar constantemente produzindo novas variedades com variações marcantes em forma e cor, algumas das quais pareciam ser novas espécies, pois as plantas da nova geração só podiam ser cruzadas entre si, não com seus pais.[74] No entanto, Hermann Joseph Muller mostrou em 1918 que as novas variedades observadas por Hugo eram o resultado de híbridos poliploides em vez de mutações genéticas rápidas.[75]
Inicialmente, de Vries e Morgan acreditavam que as mutações eram tão grandes a ponto de criar novas formas, como subespécies ou até espécies, instantaneamente. Os experimentos de Morgan com moscas-das-frutas em 1910, nos quais ele isolou mutações para características como olhos brancos, mudaram sua opinião. Ele viu que as mutações representavam pequenas características mendelianas que só se espalhariam por uma população quando fossem benéficas, auxiliadas pela seleção natural. Isso representou o germe da síntese moderna e o início do fim do mutacionismo como uma força evolutiva.[76]
Os biólogos contemporâneos aceitam que mutação e seleção desempenham papéis na evolução; a visão dominante é que, enquanto a mutação fornece material para a seleção na forma de variação, todos os resultados não aleatórios são causados pela seleção natural.[77] Masatoshi Nei argumenta, em vez disso, que a produção de genótipos mais eficientes por mutação é fundamental para a evolução, e que a evolução é frequentemente limitada por mutações.[78] A teoria endossimbiótica implica eventos raros, mas significativos, de evolução saltacional por simbiogênese.[79] Carl Woese e colegas sugeriram que a ausência de um continuum de assinaturas de RNA entre domínios de bactérias, arqueias e eucariotas mostra que essas linhagens principais se materializaram por grandes saltos na organização celular.[80] Saltos em várias escalas são considerados possíveis por mecanismos incluindo poliploidia, que certamente pode criar novas espécies de plantas,[81][82] duplicação gênica, transferência horizontal de genes,[83] e elementos de transposição (genes saltadores).[84]
Deriva genética
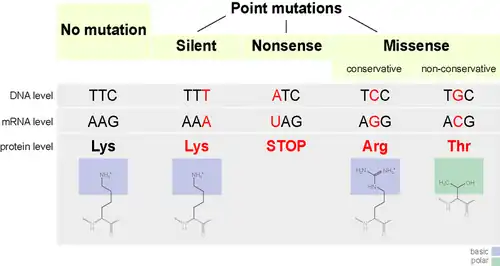
A teoria neutralista da evolução molecular, proposta por Motoo Kimura em 1968, sustenta que, no nível da molecular, a maioria das mudanças evolutivas e a maior parte da variação dentro e entre espécies não é causada pela seleção natural, mas pela deriva genética de alelos mutantes que são neutros. Uma mutação neutra é aquela que não afeta a capacidade de um organismo de sobreviver e se reproduzir. A teoria neutra permite a possibilidade de que a maioria das mutações seja deletéria, mas sustenta que, como essas são rapidamente eliminadas pela seleção natural, elas não contribuem significativamente para a variação dentro e entre espécies no nível molecular. Mutações que não são deletérias são assumidas como majoritariamente neutras, em vez de benéficas.[31]
A teoria foi controversa, pois soava como um desafio à evolução darwiniana; a controvérsia foi intensificada por um artigo de 1969 de Jack Lester King e Thomas H. Jukes, provocativamente, mas enganosamente, intitulado Non-Darwinian Evolution. Ele forneceu uma ampla variedade de evidências, incluindo comparações de sequência de proteínas, estudos do gene mutador de Treffers em E. coli, análise do código genético e imunologia comparativa, para argumentar que a maior parte da evolução de proteínas é devida a mutações neutras e deriva genética.[85][86]
Segundo Kimura, a teoria aplica-se apenas à evolução no nível molecular, enquanto a evolução fenotípica é controlada pela seleção natural, de modo que a teoria neutra não constitui uma verdadeira alternativa.[31][87]
Teorias combinadas
As várias alternativas à evolução darwiniana por seleção natural não eram necessariamente mutuamente exclusivas. A filosofia evolutiva do paleontólogo americano Edward Drinker Cope é um exemplo disso. Cope, um homem religioso, começou sua carreira negando a possibilidade de evolução. Na década de 1860, ele aceitou que a evolução poderia ocorrer, mas, influenciado por Agassiz, rejeitou a seleção natural. Cope aceitou, em vez disso, a teoria da recapitulação da história evolutiva durante o crescimento do embrião – que ontogenia recapitula filogenia, que Agassiz acreditava mostrar um plano divino levando diretamente ao homem, em um padrão revelado tanto na embriologia quanto na paleontologia. Cope não foi tão longe, vendo que a evolução criou uma árvore ramificada de formas, como Darwin havia sugerido. Cada passo evolutivo, no entanto, não era aleatório: a direção era determinada de antemão e tinha um padrão regular (ortogênese), e os passos não eram adaptativos, mas parte de um plano divino (evolução teísta). Isso deixou sem resposta a questão de por que cada passo deveria ocorrer, e Cope mudou sua teoria para acomodar a adaptação funcional para cada mudança. Ainda rejeitando a seleção natural como a causa da adaptação, Cope voltou-se para o lamarckismo para fornecer a força que guia a evolução. Finalmente, Cope supôs que o uso e desuso lamarckiano operavam causando a concentração de uma substância de força de crescimento vitalista, "batmismo", nas áreas do corpo mais intensamente usadas; por sua vez, isso fazia essas áreas se desenvolverem às custas do resto. O conjunto complexo de crenças de Cope reuniu cinco filosofias evolutivas: recapitulação, ortogênese, evolução teísta, lamarckismo e vitalismo.[88] Outros paleontólogos e naturalistas de campo continuaram a manter crenças combinando ortogênese e lamarckismo até a síntese moderna na década de 1930.[89]
Renascimento da seleção natural
No início do século XX, durante a eclipse do darwinismo, os biólogos duvidavam da seleção natural, mas igualmente descartavam rapidamente teorias como ortogênese, vitalismo e lamarckismo, que não ofereciam mecanismos para a evolução. O mutacionismo propôs um mecanismo, mas não foi geralmente aceito.[90] A síntese moderna, aproximadamente entre 1918 e 1932, amplamente eliminou todas as alternativas ao darwinismo, embora algumas, incluindo formas de ortogênese,[45] mecanismos epigenéticos que se assemelham ao lamarckismo,[53] catastrofismo,[58] estruturalismo,[69] e mutacionismo[78] tenham sido revividas, como através da descoberta de mecanismos moleculares.[91]
A biologia tornou-se darwiniana, mas a crença em alguma forma de progresso (ortogênese) permanece tanto na mente do público quanto entre os biólogos. Ruse argumenta que os biólogos evolutivos provavelmente continuarão a acreditar no progresso por três razões. Primeiramente, o princípio antrópico exige pessoas capazes de perguntar sobre o processo que levou à sua própria existência, como se fossem o ápice de tal progresso. Em segundo lugar, cientistas em geral e evolucionistas em particular acreditam que seu trabalho os está levando progressivamente mais perto de uma verdadeira compreensão da realidade, à medida que o conhecimento aumenta, e, portanto (segue o argumento), há progresso na natureza também. Ruse observa a esse respeito que Richard Dawkins compara explicitamente o progresso cultural com memes ao progresso biológico com genes. Em terceiro lugar, os evolucionistas são autosselecionados; são pessoas, como o entomologista e sociobiólogo E. O. Wilson, que estão interessadas no progresso para fornecer um significado à vida.[92]
Ver também
Notas
- ↑ Não deve ser confundido com o uso mais recente do termo evolução teísta, que se refere a uma crença teológica na compatibilidade entre ciência e religião.
- ↑ Gray, e historiadores da ciência posteriores, não se referiram a um uso do século XX de evolução teísta (descrito naquele artigo), que se pode aceitar a evolução darwiniana sem ser ateu.
Referências
- ↑ a b c d Ruse 1996, pp. 21–23.
- ↑ Bowler 1989, pp. 246–281.
- ↑ Larson 2004, pp. 12–13.
- ↑ Leroi 2015, p. 89.
- ↑ Lloyd 1968, pp. 166–169.
- ↑ a b c Leroi 2015, pp. 276–278.
- ↑ Mayr 1985, pp. 201–202.
- ↑ a b Ruse 1996, p. 43.
- ↑ Lovejoy 2011, p. 228 and passim.
- ↑ Ruse 1996, p. 95.
- ↑ a b Larson 2004, pp. 18–21.
- ↑ Larson 2004, pp. 42–43, 111.
- ↑ Bowler 1989, pp. 239, 342.
- ↑ Bowler 1989, pp. 149, 253, 259.
- ↑ Bowler 2003, p. 197.
- ↑ Larson 2004, pp. 119–120.
- ↑ Quammen 2006, pp. 209–210.
- ↑ a b c Bowler 2003, pp. 196–253.
- ↑ a b Larson 2004, pp. 105–129.
- ↑ Endersby 2007, pp. 143–147,182.
- ↑ a b BirchCobb 1985, pp. 76–78.
- ↑ a b Larson 2004, p. 126.
- ↑ a b c Larson 2004, pp. 125–128.
- ↑ Brown, Keven; Von Kitzing, Eberhard (2001). Evolution and Bahá'í Belief: ʻAbduʼl-Bahá's Response to Nineteenth-century Darwinism. [S.l.]: Kalimat Press. p. 159. ISBN 978-1-890688-08-0
- ↑ Chardin, Pierre Teilhard de (2003) [1959]. The Human Phenomenon. [S.l.]: Sussex Academic Press. p. 65. ISBN 1-902210-30-1
- ↑ a b Bowler 2003, pp. 86–95.
- ↑ Rudwick 1972, pp. 131–134.
- ↑ a b c d Ruse 2013, pp. 419.
- ↑ a b Bowler 2003, pp. 265–270.
- ↑ a b Larson 2004, pp. 127–129, 157–167.
- ↑ a b c Kimura, Motoo (1983). The neutral theory of molecular evolution. [S.l.]: Cambridge University Press. ISBN 978-0-521-31793-1
- ↑ Lewontin, R. C. (novembro de 1970). «The Units of Selection» (PDF). Annual Review of Ecology, Evolution, and Systematics. 1 (1): 1–18. Bibcode:1970AnRES...1....1L. JSTOR 2096764. doi:10.1146/annurev.es.01.110170.000245
- ↑ a b c Bechtel, William; Richardson, Robert C. (1998). «Vitalism». In: Craig, E. Routledge Encyclopedia of Philosophy. Routledge
- ↑ Jidenu, Paulin (1996). African Philosophy. [S.l.]: Indiana University Press. p. 16. ISBN 978-0-253-21096-8
- ↑ Kinne-Saffran, E.; Kinne, R. K. H. (1999). «Vitalism and Synthesis of Urea». American Journal of Nephrology. 19 (2): 290–292. PMID 10213830. doi:10.1159/000013463
- ↑ Driesch, Hans (1914). The History and Theory of Vitalism. [S.l.]: Macmillan
- ↑ Bedau, Mark A.; Cleland, Carol E. (2010). The Nature of Life: Classical and Contemporary Perspectives from Philosophy and Science. [S.l.]: Cambridge University Press. p. 95. ISBN 978-0-521-51775-1
- ↑ Gray, Asa (1876). Essays and Reviews Pertaining to Darwinism. [S.l.]: Appleton. doi:10.5962/bhl.title.19483. hdl:2027/miun.agj4537.0001.001
- ↑ Campbell, George (1867). The Reign of Law. [S.l.]: Strahan
- ↑ Morris, John D.; Morris, Henry M. (1998). The Modern Creation Trilogy. [S.l.]: New Leaf. p. 36
- ↑ a b Bowler 2003, p. 249.
- ↑ a b Quammen 2006, p. 221.
- ↑ Yampolsky, L. Y.; Stoltzfus, A. (2001). «Bias in the introduction of variation as an orienting factor in evolution». Evolution & Development. 3 (2): 73–83. PMID 11341676. doi:10.1046/j.1525-142x.2001.003002073.x
- ↑ Stoltzfus, A. (2006). «Mutation-Biased Adaptation in a Protein NK Model». Molecular Biology and Evolution. 23 (10): 1852–1862. PMID 16857856. doi:10.1093/molbev/msl064

- ↑ a b Stoltzfus, A.; Yampolsky, L. Y. (2009). «Climbing Mount Probable: Mutation as a Cause of Nonrandomness in Evolution». J Hered. 100 (5): 637–647. PMID 19625453. doi:10.1093/jhered/esp048
- ↑ Newman, S. A. (2014). «Form and function remixed: developmental physiology in the evolution of vertebrate body plans». Journal of Physiology. 592 (11): 2403–2412. PMC 4048098. PMID 24817211. doi:10.1113/jphysiol.2014.271437
- ↑ Darwin 1872, p. 108.
- ↑ a b Bowler 2003, p. 236.
- ↑ a b Larson 2004, pp. 125–129.
- ↑ Quammen 2006, p. 217.
- ↑ Bowler 2003, pp. 239–240.
- ↑ Bowler 2003, pp. 253–255.
- ↑ a b Heard, E.; Martienssen, R. A. (2014). «Transgenerational epigenetic inheritance: myths and mechanisms». Cell. 157 (1): 95–109. PMC 4020004. PMID 24679529. doi:10.1016/j.cell.2014.02.045
- ↑ McGowan 2001, pp. 3–6.
- ↑ Rudwick 1972, pp. 133–134.
- ↑ Waggoner, Ben (1996). «Georges Cuvier (1769–1832)». University of California, Berkeley
- ↑ Rudwick 1972, p. 131.
- ↑ a b Alvarez, Luis W.; Alvarez, Walter; Asaro, F.; Michel, H. V. (1980). «Extraterrestrial cause for the Cretaceous–Tertiary extinction». Science. 208 (4448): 1095–1108. Bibcode:1980Sci...208.1095A. CiteSeerX 10.1.1.126.8496. PMID 17783054. doi:10.1126/science.208.4448.1095
- ↑ Hildebrand, A. R.; Penfield, G. T.; et al. (1991). «Chicxulub crater: a possible Cretaceous/Tertiary boundary impact crater on the Yucatán peninsula, Mexico». Geology. 19 (9): 867–871. Bibcode:1991Geo....19..867H. doi:10.1130/0091-7613(1991)019<0867:ccapct>2.3.co;2
- ↑ Racine, Valerie (7 de outubro de 2013). «Essay: The Cuvier-Geoffroy Debate». The Embryo Project Encyclopedia, Arizona State University. Consultado em 10 de dezembro de 2016
- ↑ Seilacher 1991, pp. 251–271.
- ↑ Seilacher, Adolf (julho de 1989). «Vendozoa: Organismic construction in the Proterozoic biosphere». Lethaia. 22 (3): 229–239. Bibcode:1989Letha..22..229S. doi:10.1111/j.1502-3931.1989.tb01332.x
- ↑ a b c Wagner, Günter P., Homology, Genes, and Evolutionary Innovation. Princeton University Press. 2014. Capítulo 1: The Intellectual Challenge of Morphological Evolution: A Case for Variational Structuralism. Páginas 7–38
- ↑ Simpson, Carl; Erwin, Douglas H. (quoted) (13 de abril de 2014). Homology, Genes, and Evolutionary Innovation Günter P. Wagner. [S.l.]: Princeton University Press. ISBN 9780691156460
- ↑ Fox, Ronald F. (dezembro de 1993). «Review of Stuart Kauffman, The Origins of Order: Self-Organization and Selection in Evolution». Biophys. J. 65 (6): 2698–2699. Bibcode:1993BpJ....65.2698F. PMC 1226010. doi:10.1016/S0006-3495(93)81321-3
- ↑ Denton, Michael J. (agosto de 2013). «The Types: A Persistent Structuralist Challenge to Darwinian Pan-Selectionism». BIO-Complexity. 2013 (3). doi:10.5048/BIO-C.2013.3
- ↑ Erwin, Douglas H. (setembro de 2011). «Evolutionary uniformitarianism». Developmental Biology. 357 (1): 27–34. Bibcode:2011DevBi.357...27E. PMID 21276788. doi:10.1016/j.ydbio.2011.01.020
- ↑ Muller, G.B.; Newman, S.A. The innovation triad: an EvoDevo agenda. J. Exp. Zool. (Mol. Dev Evol), 304B (2005), pp. 487–503
- ↑ a b Goodwin, Brian (2009). «Beyond the Darwinian Paradigm: Understanding Biological Forms». In: Ruse, Michael; Travis, Joseph. Evolution: The First Four Billion Years. [S.l.]: Harvard University Press
- ↑ Moran, Laurence A. (2 de fevereiro de 2016). «What is "Structuralism"?». Sandwalk (blog of a recognised expert). Consultado em 9 de dezembro de 2016
- ↑ Held, Lewis Irving (2014). How the snake lost its legs: curious tales from the frontier of evo-devo. Cambridge, England: Cambridge University Press. p. 121. ISBN 978-1-107-62139-8
- ↑ Stoltzfus, A.; Cable, K. (2014). «Mendelian-Mutationism: The Forgotten Evolutionary Synthesis». Journal of the History of Biology. 47 (4): 501–546. PMID 24811736. doi:10.1007/s10739-014-9383-2
- ↑ Bowler 1989, pp. 211–212, 276–277.
- ↑ Endersby 2007, pp. 148–162.
- ↑ Endersby 2007, pp. 202–205.
- ↑ Bowler 1989, pp. 310–311.
- ↑ Wagner, G. P. (1 de janeiro de 2013). «The Changing Face of Evolutionary Thinking». Genome Biology and Evolution. 5 (10): 2006–2007. PMC 3814208. doi:10.1093/gbe/evt150
- ↑ a b Stoltzfus, A. (2014). «In search of mutation-driven evolution». Evolution & Development. 16: 57–59. doi:10.1111/ede.12062
- ↑ Syvanen, Michael; Kado, Clarence I. (2002). Horizontal Gene Transfer. [S.l.]: Academic Press. p. 405. ISBN 978-0-12-680126-2
- ↑ Roberts, E.; Sethi, A.; Montoya, J.; Woese, C .R.; Luthey-Schulten, Z. (2008). «Molecular signatures of ribosomal evolution». Proc. Natl. Acad. Sci. USA. 105 (37): 13953–13958. Bibcode:2008PNAS..10513953R. PMC 2528867. PMID 18768810. doi:10.1073/pnas.0804861105
- ↑ Dufresne, France; Herbert, Paul D. N. (1994). «Hybridization and origins of polyploidy». Proceedings: Biological Sciences. 258 (1352): 141–146. Bibcode:1994RSPSB.258..141D. JSTOR 49988. doi:10.1098/rspb.1994.0154
- ↑ Oladele Ogunseitan. (2004). Microbial Diversity: Form and Function in Prokaryotes. Wiley-Blackwell. p. 13. ISBN 978-0-632-04708-6
- ↑ Serres, M. H.; Kerr, A. R.; McCormack, T. J.; Riley, M. (2009). «Evolution by leaps: gene duplication in bacteria». Biology Direct. 4: 46. PMC 2787491. PMID 19930658. doi:10.1186/1745-6150-4-46
- ↑ McClintock, B. (1984). The significance of responses of the genome to challenge. Science Vol. 226, pp. 792–801.
- ↑ King, Jack Lester; Thomas H. Jukes (1969). «Non-Darwinian Evolution». Science. 164 (3881): 788–798. Bibcode:1969Sci...164..788L. PMID 5767777. doi:10.1126/science.164.3881.788
- ↑ Dietrich, Michael R. (1994). «The origins of the neutral theory of molecular evolution». Journal of the History of Biology. 27 (1): 21–59. PMID 11639258. doi:10.1007/BF01058626
- ↑ Hughes, Austin L. (2007). «Looking for Darwin in all the wrong places: the misguided quest for positive selection at the nucleotide sequence level». Heredity. 99 (4): 364–373. PMID 17622265. doi:10.1038/sj.hdy.6801031
- ↑ a b Bowler 1989, pp. 261–262.
- ↑ Bowler 1989, p. 264.
- ↑ Larson 2004, pp. 128–129.
- ↑ Larson 2004, pp. 222–243, 284–286.
- ↑ Ruse 1996, pp. 536–539.
Fontes
- Birch, Charles; Cobb, John B. (1985). The Liberation of Life: From the Cell to the Community. [S.l.]: University of North Texas. ISBN 978-0-9626807-0-0
- Bowler, Peter J. (1989) [1983]. The Eclipse of Darwinism: anti-Darwinian evolutionary theories in the decades around 1900. [S.l.]: Johns Hopkins University Press. ISBN 978-0-8018-4391-4
- Bowler, Peter J. (2003). Evolution:The History of an Idea. [S.l.]: University of California Press. ISBN 978-0-520-23693-6
- Darwin, Charles (1872). The Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life 6th ed. London: John Murray. ISBN 978-1-904633-78-5
- Endersby, Jim (2007). A Guinea Pig's History of Biology. [S.l.]: Harvard University Press. ISBN 978-0-674-02713-8
- Larson, Edward J. (2004). Evolution: The Remarkable History of Scientific Theory. [S.l.]: Modern Library. ISBN 978-0-679-64288-6
- Leroi, Armand Marie (2015) [2014]. The Lagoon: How Aristotle Invented Science. [S.l.]: Bloomsbury. ISBN 978-1-4088-3622-4
- Lloyd, G. E. R. (1968). Aristotle: The Growth and Structure of His Thought. [S.l.]: Cambridge University Press. ISBN 978-0-521-09456-6
- Lovejoy, Arthur O. (2011) [1936]. The Great Chain of Being: A Study of the History of an Idea. [S.l.]: Transaction Publishers. ISBN 978-0-674-36153-9
- Mayr, Ernst (1985). The Growth of Biological Thought: Diversity, Evolution, and Inheritance. [S.l.]: Harvard University Press. ISBN 978-0-674-36446-2
- McGowan, Christopher (2001). The Dragon Seekers. Cambridge, Massachusetts: Perseus Publishing. ISBN 978-0-7382-0282-2
- Quammen, David (2006). The Reluctant Mr. Darwin. [S.l.]: Atlas Books. ISBN 978-0-393-05981-6
- Rudwick, Martin J. S. (1972). The Meaning of Fossils. Chicago, Illinois: University of Chicago Press. ISBN 978-0-226-73103-2
- Ruse, Michael (1996). Monad to man: the Concept of Progress in Evolutionary Biology. [S.l.]: Harvard University Press. ISBN 978-0-674-03248-4
- Ruse, Michael (2013). «17. From Organicism to Mechanism-and Halfway Back?». In: Henning, Brian G.; Scarfe, Adam. Beyond Mechanism: Putting Life Back Into Biology. [S.l.]: Lexington Books. p. 419. ISBN 9780739174371
- Seilacher, Adolf (1991). «Self-Organizing Mechanisms in Morphogenesis and Evolution». In: Schmidt-Kittler, Norbert; Vogel, Klaus. Constructional Morphology and Evolution. [S.l.]: Springer. pp. 251–271. ISBN 978-3-642-76158-4. doi:10.1007/978-3-642-76156-0_17