Evolução dos sistemas nervosos
A evolução dos sistemas nervosos remonta ao primeiro desenvolvimento de sistemas nervosos em animais (ou metazoários). Os neurônios surgiram como células especializadas em sinalização elétrica em animais multicelulares, adaptando o mecanismo dos potenciais de ação presentes em eucariotos unicelulares e coloniais móveis. Sistemas primitivos, como os encontrados em protistas, utilizam sinalização química para movimento e sensibilidade; evidências sugerem que esses foram precursores dos tipos celulares neurais modernos e suas sinapses.[1] Quando alguns animais começaram a adotar um estilo de vida móvel e a consumir partículas de alimento maiores por digestão extracelular [en], desenvolveram epitélios ciliados, músculos contráteis e neurônios coordenadores e sensíveis para isso em sua camada externa, a ectoderme.[2][3]
Redes nervosas simples, observadas em acelos (bilaterais basais) e cnidários, são consideradas a condição ancestral dos planulozoários (bilaterais, cnidários e, possivelmente, placozoários). Uma rede nervosa mais complexa, com cordões nervosos simples, está presente em animais antigos chamados ctenóforos,[4] mas não há nervos, e portanto nenhum sistema nervoso, em outro grupo de animais antigos, as esponjas (Porifera). Devido à presença comum e à semelhança de alguns genes neurais nesses animais antigos e em seus parentes protistas, à controvérsia sobre se ctenóforos ou esponjas divergiram primeiro, e à recente descoberta de células "neuroides" especializadas na coordenação de coanócitos digestivos em Spongilla [en], a origem dos neurônios na árvore filogenética da vida permanece em debate.[5][6][7] A cefalização adicional e a evolução dos cordões nervosos (ventral [en]e dorsal [en]) ocorreram várias vezes de forma independente nos bilaterais.[5]
Precursores neurais
Os potenciais de ação, essenciais para a atividade neural, evoluíram em eucariotos unicelulares. Estes utilizam potenciais de ação baseados em cálcio, em vez de sódio, mas o mecanismo provavelmente foi adaptado para a sinalização elétrica neural em animais multicelulares. Em alguns eucariotos coloniais, como Obelia [en], os sinais elétricos se propagam não apenas por redes neurais, mas também por células epiteliais no sistema digestivo compartilhado da colônia.[8] Vários filos não metazoários, incluindo coanoflagelados, filasterea e mesomicetozoários, possuem homólogos de proteínas sinápticas, como SNAREs secretórios, Shank e Homer. Em coanoflagelados e mesomicetozoários, essas proteínas são reguladas positivamente durante fases coloniais, sugerindo a importância desses proto-sinápticos para a comunicação célula a célula.[9] A história das ideias sobre como neurônios e os primeiros sistemas nervosos surgiram na evolução foi discutida em um livro de 2015 por Michel Anctil.[10] Em 2022, duas proteínas, SMIM20 e NUCB2, precursoras dos neuropeptídeos phoenixin e nesfatin-1, respectivamente, mostraram homologia profunda em todas as linhagens que precederam criaturas com sistemas nervosos centrais, incluindo bilaterais, cnidários, ctenóforos e esponjas, bem como em coanoflagelados.[11][12]
Esponjas
As esponjas não possuem células conectadas por junções sinápticas, ou seja, não têm neurônios e, portanto, nenhum sistema nervoso. No entanto, elas possuem homólogos de muitos genes que desempenham papéis-chave na função sináptica. Estudos recentes mostraram que células de esponjas expressam um grupo de proteínas que se agrupam para formar uma estrutura semelhante a uma densidade pós-sináptica [en] (a parte receptora de sinal de uma sinapse).[13] Contudo, a função dessa estrutura ainda é incerta. Embora as células das esponjas não exibam transmissão sináptica, elas se comunicam por meio de ondas de cálcio e outros impulsos, que mediam ações simples, como a contração do corpo inteiro.[14] Outras formas de comunicação entre células de esponjas incluem o transporte vesicular através de regiões densas das membranas celulares. Esses vesículos transportam íons e moléculas sinalizadoras, mas não possuem função sináptica verdadeira.[15]
Redes nervosas
Águas-vivas, ctenóforos e animais relacionados possuem redes nervosas difusas em vez de um sistema nervoso central. Na maioria das águas-vivas, a rede nervosa é distribuída de forma mais ou menos uniforme pelo corpo; nos ctenóforos, ela se concentra perto da boca. As redes nervosas consistem em neurônios sensoriais que captam sinais químicos, táteis e visuais, neurônios motores que ativam contrações da parede corporal e neurônios intermediários que detectam padrões de atividade nos neurônios sensoriais e enviam sinais a grupos de neurônios motores. Em alguns casos, grupos de neurônios intermediários formam gânglios distintos.[16]
O desenvolvimento do sistema nervoso nos radiados é relativamente desestruturado. Diferentemente dos bilaterais, os radiados possuem apenas duas camadas celulares primordiais, a endoderme e a ectoderme. Os neurônios são gerados a partir de um conjunto especial de células precursoras ectodérmicas, que também servem como precursoras de todos os outros tipos celulares ectodérmicos.[17]
Cordões nervosos

A grande maioria dos animais existentes são bilaterais, ou seja, animais com lados esquerdo e direito que são imagens espelhadas aproximadas um do outro. Todos os bilaterais teriam descendido de um ancestral comum semelhante a um verme, que surgiu no período Criogeniano, há 700–650 milhões de anos.[18] A forma corporal fundamental dos bilaterais é um tubo com uma cavidade intestinal oca que vai da boca ao ânus, e um cordão nervoso com um gânglio especialmente grande na frente, chamado de "cérebro".
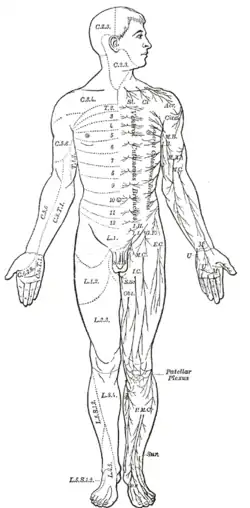
Mesmo mamíferos, incluindo humanos, exibem o plano corporal segmentado dos bilaterais no nível do sistema nervoso. A medula espinhal contém uma série de gânglios segmentares, cada um dando origem a nervos motores e sensoriais que inervam uma porção da superfície corporal e da musculatura subjacente. Nos membros, o padrão de inervação é complexo, mas no tronco ele forma uma série de faixas estreitas. Os três segmentos superiores pertencem ao cérebro, dando origem ao prosencéfalo, mesencéfalo e rombencéfalo.[19]
Os bilaterais podem ser divididos, com base em eventos que ocorrem muito cedo no desenvolvimento embrionário, em dois grupos (superfilos) chamados protóstomos e deuteróstomos.[20] Os deuteróstomos incluem vertebrados, equinodermos e hemicordados (principalmente da classe Enteropneusta). Os protóstomos, o grupo mais diverso, incluem artrópodes, moluscos e vários tipos de vermes. Há uma diferença básica entre os dois grupos na posição do sistema nervoso no corpo: os protóstomos possuem um cordão nervoso no lado ventral (geralmente inferior), enquanto nos deuteróstomos o cordão nervoso está no lado dorsal (geralmente superior). De fato, vários aspectos do corpo são invertidos entre os dois grupos, incluindo os padrões de expressão de vários genes que mostram gradientes dorsoventrais. Alguns anatomistas agora consideram que os corpos de protóstomos e deuteróstomos são "invertidos" um em relação ao outro, uma hipótese proposta inicialmente por Geoffroy Saint-Hilaire para insetos em comparação com vertebrados. Assim, por exemplo, os insetos têm cordões nervosos que correm ao longo da linha média ventral do corpo, enquanto todos os vertebrados têm medulas espinhais que correm ao longo da linha média dorsal.[21] Mas dados moleculares recentes de diferentes protóstomos e deuteróstomos rejeitam esse cenário e sugerem que os cordões nervosos evoluíram independentemente em ambos.[22]
Anelídeos
Minhocas possuem cordões nervosos ventrais duplos que percorrem o comprimento do corpo e se fundem na cauda e na boca. Esses cordões nervosos são conectados por nervos transversais, como os degraus de uma escada. Esses nervos transversais ajudam a coordenar os dois lados do animal. Dois gânglios na extremidade da cabeça funcionam de forma semelhante a um cérebro simples. Fotorreceptores nos olhos do animal fornecem informações sensoriais sobre claro e escuro.[23]
Nematoides
O sistema nervoso de um nematóide muito pequeno, Caenorhabditis elegans, foi mapeado até o nível sináptico. Cada neurônio e sua linhagem celular foram registrados, e a maioria, senão todas, as conexões neurais são conhecidas. Nessa espécie, o sistema nervoso é sexualmente dimórfico; os sistemas nervosos dos dois sexos, machos e hermafroditas, têm diferentes números de neurônios e grupos de neurônios que desempenham funções específicas de cada sexo. Em C. elegans, os machos têm exatamente 383 neurônios, enquanto os hermafroditas têm exatamente 302 neurônios.[24]
Artrópodes
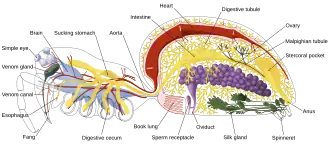
Artrópodes, como insetos e crustáceos, possuem um sistema nervoso composto por uma série de gânglios conectados por um cordão nervoso ventral formado por dois conectivos paralelos que percorrem o comprimento do abdômen.[25] Tipicamente, cada segmento do corpo tem um gânglio de cada lado, embora alguns gânglios sejam fundidos para formar o cérebro e outros gânglios maiores. O segmento da cabeça contém o cérebro, também conhecido como gânglio supraesofágico [en]. No sistema nervoso dos insetos, o cérebro é anatomicamente dividido em protocérebro, deutocérebro e tritocérebro. Imediatamente atrás do cérebro está o gânglio subesofágico [en], composto por três pares de gânglios fundidos. Ele controla as peças bucais [en], as glândulas salivares e certos músculos. Muitos artrópodes possuem órgãos sensoriais bem desenvolvidos, incluindo olhos compostos para visão e antenas para olfato e sensação de feromônios. As informações sensoriais desses órgãos são processadas pelo cérebro.
Em insetos, muitos neurônios têm corpos celulares posicionados na borda do cérebro e são eletricamente passivos — os corpos celulares servem apenas para fornecer suporte metabólico e não participam da sinalização. Uma fibra protoplasmática sai do corpo celular e se ramifica profusamente, com algumas partes transmitindo sinais e outras recebendo sinais. Assim, a maior parte do cérebro dos insetos tem corpos celulares passivos dispostos na periferia, enquanto o processamento de sinais neurais ocorre em um emaranhado de fibras protoplasmáticas chamado neurópilo [en], no interior.[26]
Evolução dos sistemas nervosos centrais
Evolução do cérebro humano
Houve um aumento gradual no volume cerebral à medida que os ancestrais dos humanos modernos progrediram ao longo da linha do tempo da evolução humana (ver Homininae), começando com cerca de 600 cm³ em Homo habilis até 1736 cm³ em Homo neanderthalensis. Assim, em geral, há uma correlação entre o volume cerebral e a inteligência.[27] No entanto, os modernos Homo sapiens têm um volume cerebral menor (tamanho cerebral de 1.250 cm³) que os neandertais; as mulheres têm um volume cerebral ligeiramente menor que os homens, e os homem de flores (Homo floresiensis), apelidados de "hobbits", tinham uma capacidade craniana de cerca de 380 cm³, cerca de um terço da média de Homo erectus e considerada pequena para um chimpanzé. Propõe-se que eles evoluíram de H. erectus como um caso de nanismo insular. Apesar de seu cérebro três vezes menor, há evidências de que H. floresiensis usava fogo e fabricava ferramentas de pedra tão sofisticadas quanto as de seu suposto ancestral, H. erectus.[28] Iain Davidson resume as restrições evolutivas opostas ao tamanho do cérebro humano como "tão grande quanto você precisa e tão pequeno quanto você pode".[29] O cérebro humano evoluiu em torno das necessidades metabólicas, ambientais e sociais que a espécie enfrentou ao longo de sua existência. À medida que as espécies de hominídeos evoluíram com tamanho cerebral e poder de processamento aumentados, a necessidade metabólica geral também cresceu. Comparados aos chimpanzés, os humanos consomem mais calorias de animais do que de plantas. Embora não seja certo, estudos sugerem que essa mudança na dieta se deve à maior necessidade de ácidos graxos mais facilmente encontrados em produtos animais. Esses ácidos graxos são essenciais para a manutenção e o desenvolvimento do cérebro. Outros fatores a considerar são a necessidade de interação social e como os hominídeos interagiram com seus ambientes ao longo do tempo.[30]
A evolução do cérebro pode ser estudada usando moldes endocast, um ramo da neurologia e paleontologia chamado paleoneurologia.
Ver também
Referências
- ↑ «sistema nervoso | Definição, Função, Estrutura e Fatos». Encyclopædia Britannica (em inglês). Consultado em 31 de março de 2025
- ↑ Arendt, D. (2021). Elementary Nervous Systems. Philosophical Transactions of the Royal Society B: Biological Sciences, 376 (1821), 20200347. https://doi.org/10.1098/rstb.2020.0347
- ↑ Arendt, D., Benito-Gutierrez, E., Brunet, T., & Marlow, H. (2015). Gastric pouches and the mucociliary sole: Setting the stage for nervous system evolution. Philosophical Transactions of the Royal Society B: Biological Sciences, 370 (1684), 20150286. https://doi.org/10.1098/rstb.2015.0286
- ↑ Jékely, G., Paps, J. & Nielsen, C. The phylogenetic position of ctenophores and the origin(s) of nervous systems. EvoDevo 6, 1 (2015). https://doi.org/10.1186/2041-9139-6-1
- ↑ a b Moroz, L. L., Romanova, D. Y., & Kohn, A. B. (2021). Neural versus alternative integrative systems: Molecular insights into origins of neurotransmitters. Philosophical Transactions of the Royal Society B: Biological Sciences, 376 (1821), 20190762. https://doi.org/10.1098/rstb.2019.0762
- ↑ Musser, J. M., Schippers, K. J., Nickel, M. et al. (2021). Profiling cellular diversity in sponges informs animal cell type and nervous system evolution. Science, 374 (6568), 717–723. https://doi.org/10.1126/science.abj2949
- ↑ Hayakawa, E., Guzman, C., Horiguchi, O. et al. Mass spectrometry of short peptides reveals common features of metazoan peptidergic neurons. Nat Ecol Evol (2022). https://doi.org/10.1038/s41559-022-01835-7
- ↑ Matthews GG (2001). «Evolution of nervous systems». Neurobiology: molecules, cells, and systems. [S.l.]: Wiley-Blackwell. p. 21. ISBN 978-0-632-04496-2
- ↑ Burkhardt P, Sprecher SG (outubro de 2017). «Evolutionary origin of synapses and neurons - Bridging the gap». BioEssays. 39 (10). 1700024 páginas. PMID 28863228. doi:10.1002/bies.201700024

- ↑ Anctil M (2015). Dawn of the Neuron: The Early Struggles to Trace the Origin of Nervous Systems. Montreal & Kingston, London, Chicago: McGill-Queen's University Press. ISBN 978-0-7735-4571-7
- ↑ Yañez-Guerra LA, Thiel D, Jékely G (abril de 2022). O'Connell M, ed. «Premetazoan Origin of Neuropeptide Signaling». Molecular Biology and Evolution. 39 (4): msac051. PMC 9004410. PMID 35277960. doi:10.1093/molbev/msac051
- ↑ Callier V (3 de junho de 2022). «Brain-Signal Proteins Evolved Before Animals Did». Quanta Magazine (em inglês). Consultado em 31 de março de 2025
- ↑ Sakarya O, Armstrong KA, Adamska M, Adamski M, Wang IF, Tidor B, et al. (junho de 2007). Vosshall L, ed. «A post-synaptic scaffold at the origin of the animal kingdom». PLOS ONE. 2 (6): e506. Bibcode:2007PLoSO...2..506S. PMC 1876816. PMID 17551586. doi:10.1371/journal.pone.0000506
- ↑ Jacobs DK, Nakanishi N, Yuan D, Camara A, Nichols SA, Hartenstein V (novembro de 2007). «Evolution of sensory structures in basal metazoa». Integrative and Comparative Biology. 47 (5): 712–723. PMID 21669752. doi:10.1093/icb/icm094
- ↑ Leys SP (fevereiro de 2015). «Elements of a 'nervous system' in sponges». The Journal of Experimental Biology. 218 (Pt 4): 581–591. Bibcode:2015JExpB.218..581L. PMID 25696821. doi:10.1242/jeb.110817
- ↑ Ruppert EE, Fox RS, Barnes RD (2004). Invertebrate Zoology 7th ed. [S.l.]: Brooks / Cole. pp. 111–124. ISBN 978-0-03-025982-1
- ↑ Sanes DH, Reh TA, Harris WA (2006). Development of the nervous system. [S.l.]: Academic Press. pp. 3–4. ISBN 978-0-12-618621-5
- ↑ Balavoine G, Adoutte A (fevereiro de 2003). «The segmented urbilateria: a testable scenario». Integrative and Comparative Biology. 43 (1): 137–147. PMID 21680418. doi:10.1093/icb/43.1.137
- ↑ Ghysen A (2003). «The origin and evolution of the nervous system». The International Journal of Developmental Biology. 47 (7–8): 555–562. PMID 14756331
- ↑ Erwin DH, Davidson EH (julho de 2002). «The last common bilaterian ancestor». Development. 129 (13): 3021–3032. PMID 12070079. doi:10.1242/dev.129.13.3021
- ↑ Lichtneckert R, Reichert H (maio de 2005). «Insights into the urbilaterian brain: conserved genetic patterning mechanisms in insect and vertebrate brain development». Heredity. 94 (5): 465–477. PMID 15770230. doi:10.1038/sj.hdy.6800664
- ↑ Martín-Durán JM, Pang K, Børve A, Lê HS, Furu A, Cannon JT, et al. (janeiro de 2018). «Convergent evolution of bilaterian nerve cords». Nature. 553 (7686): 45–50. Bibcode:2018Natur.553...45M. PMC 5756474. PMID 29236686. doi:10.1038/nature25030
- ↑ Adey WR (fevereiro de 1951). «The nervous system of the earthworm Megascolex». The Journal of Comparative Neurology. 94 (1): 57–103. PMID 14814220. doi:10.1002/cne.900940104
- ↑ «Specification of the nervous system». Wormbook
- ↑ Chapman RF (1998). «Ch. 20: Nervous system». The insects: structure and function. [S.l.]: Cambridge University Press. pp. 533–568. ISBN 978-0-521-57890-5
- ↑ Chapman, p. 546
- ↑ Ko KH (2016). «Origins of human intelligence: The chain of tool-making and brain evolution» (PDF). Anthropological Notebooks. 22 (1): 5–22
- ↑ Brown P, Sutikna T, Morwood MJ, Soejono RP, Saptomo EW, Due RA (outubro de 2004). «A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia» (PDF). Nature. 431 (7012): 1055–1061. Bibcode:2004Natur.431.1055B. PMID 15514638. doi:10.1038/nature02999
- ↑ Davidson I (2007). «As large as you need and as small as you can": Implications of the brain size of Homo floresiensis». In: Schalley AC, Khlentzos D. Mental States Volume 1: Evolution, function, nature. Amsterdam: J. Benjamins Pub. Co. pp. 35–42. ISBN 978-90-272-9121-9
- ↑ Barrickman NL (1 de janeiro de 2017). «4.04 – Energetics, Life History, and Human Brain Evolution». In: Kaas JH. Evolution of Nervous Systems (em inglês) Second ed. Oxford: Academic Press. pp. 51–62. ISBN 978-0-12-804096-6