Syssomonas
Syssomonas
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Espécie-tipo | |||||||||||||||||
| Syssomonas multiformis[1] Tikhonenkov et al. 2017 | |||||||||||||||||
| Estirpe-tipo | |||||||||||||||||
| Colp-12 MI-PR205[a] | |||||||||||||||||
| Espécies | |||||||||||||||||
| |||||||||||||||||

Syssomonas é um gênero monotípico de protistas unicelulares flagelados que contém a espécie Syssomonas multiformis.[1] Pertence à Pluriformea [en], dentro da linhagem de Holozoa, um clado que inclui animais e seus parentes protistas mais próximos. Vive em habitats de água doce. Possui um ciclo de vida complexo que inclui as fases unicelulares ameboide e flagelada, além de agregados multicelulares, dependendo do meio de crescimento e do estado nutricional.
Estágios de vida
Syssomonas multiformis é uma espécie de protistas unicelulares com células nuas (sem testas ou escamas) que apresenta uma variedade de formas de vida durante seu ciclo de vida complexo. Essas formas incluem: células flageladas redondas (7–14 μm de diâmetro) com um flagelo posterior, células ameboflageladas (com flagelos e pseudópodes), células ameboides não flageladas e cistos esféricos. Também podem formar agregados de múltiplas células.[1][2]
Estágios unicelulares: flagelar, ameboide e cisto
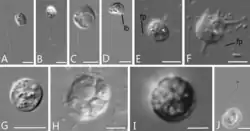
Na fase de natação flagelar, a mais comum em seu ciclo de vida, as células de Syssomonas lembram uma célula típica de Opisthokonta, semelhante a espermatozoides de animais ou zoósporos de quitrídios. Possuem um flagelo liso que emerge do ponto médio-lateral da célula, voltando-se para trás durante a natação. Durante o movimento, o batimento rápido do flagelo pode dar a aparência de dois flagelos. As células nadadoras giram e podem parar subitamente e mudar de direção. Células solitárias podem se fixar temporariamente a um substrato pela parte anterior do corpo celular, produzindo fluxo de água por batimento rápido do flagelo, semelhante a coanoflagelados ou coanócitos de esponjas. Células flutuantes descem para se transformar em ameboflageladas, gerando lobopódios largos e filopódios curtos e finos, e diminuindo o batimento do flagelo. As ameboflageladas podem rastejar ao longo do substrato por meio de seus lobopódios anteriores.[2]
Os estágios ameboflagelados de Syssomonas podem perder o flagelo de três maneiras: descartando-o abruptamente, retraindo-o para dentro da célula quando esticado ou enrolando-o sob a célula e retraindo-o como uma espiral. Como resultado, tornam-se a fase ameboide, que produz filopódios curtos e finos e, às vezes, possui dois vacúolos contráteis. Tanto as amebas quanto as ameboflageladas podem voltar a ser flageladas.[2]
A fase ameboide pode retrair seus filopódios e se tornar um cisto redondo, no qual ocorre a divisão celular palintômica (ou seja, divisões celulares rápidas sem crescimento citoplasmático entre elas, uma característica da clivagem embrionária de animais),[3] gerando 2, 4, 8 ou 16 células flageladas que são liberadas de dentro do cisto.[2]
Estágios multicelulares agregativos

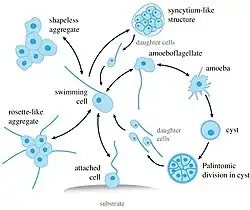
As células de Syssomonas podem se fundir parcialmente e formar agregações temporárias de cerca de 3 a 10 células, geralmente sem forma definida e observadas próximas ao fundo da coluna de água. Também podem se agregar unindo apenas células flageladas, com seus flagelos voltados para fora, semelhantes às colônias em forma de roseta de coanoflagelados. Ambas as agregações se desfazem facilmente, e suas membranas celulares não estão fundidas.[2]
Em culturas microbiológicas sólidas, células solitárias podem, às vezes, se fundir completamente no fundo da placa de Petri em uma estrutura semelhante a um sincício ou pseudoplasmódio, na qual os núcleos não se fundem. A partir desses sincícios, ocorre a gemulação de células-filhas. Esse fenômeno de gemulação a partir de sincícios não foi observado em outros eucariontes, embora a formação de células multinucleadas como resultado da agregação de múltiplas células seja conhecida em outras linhagens de protistas (membros da ordem Dictyosteliida em Eumycetozoa [en], Copromyxa [en] em Lobosa,[4] acrasídeos em Excavata,[5] Sorogena em Alveolata,[6] Sorodiplophrys em Stramenopiles, Guttulinopsis em Rhizaria,[7] e Fonticula alba dentro de Opisthokonta).[8] A transição de uma fase filopodial ameboide para uma fase agregativa também é observada em outro membro de Holozoa, Capsaspora owczarzaki [en]. A formação de sincícios também ocorre em animais; o citoplasma de esponjas-de-vidro, tegumentos de platelmintos e os músculos esqueléticos e a placenta de mamíferos são todas estruturas sinciciais.[2]
A fusão de células em Syssomonas atrai, provavelmente por quimiotaxia, outras células próximas que nadam ativamente e tentam se aderir aos agregados. Esse parece ser o único método pelo qual os agregados crescem, em oposição à divisão celular.[2]
Ecologia
Syssomonas multiformis foi isolada de uma poça de água doce no Vietnã. O organismo pode sobreviver a temperaturas variando de 5 a 36 °C. Alimenta-se do conteúdo citoplasmático de outros eucariontes de tamanho semelhante, uma característica incomum entre membros unicelulares de Holozoa. Em particular, é um predador de Chrysophyceae e Bodonida [en] heterotróficos (por exemplo, Parabodo caudatus e espécies de Spumella).[2] Também pode engolir bactérias e pequenos detritos, de maneira semelhante aos coanoflagelados.[1]
Diferentemente de muitos outros protistas eucariontes, as células de Syssomonas não possuem organelas extrusivas para caça. Em vez disso, elas se fixam à célula presa e sugam seu citoplasma sem ingerir a membrana celular. Alimentam-se melhor de células inativas, lentas ou mortas, ou de cistos. Provavelmente por quimiotaxia, após uma célula se fixar à presa, outras células de Syssomonas são atraídas para a mesma célula presa e tentam se fixar a ela. Várias células podem sugar o citoplasma da mesma célula presa em conjunto.[2]
Elas usam pseudópodes curtos para se alimentar de aglomerados de bactérias. Após isso, formam um grande vacúolo alimentar na extremidade posterior da célula. No entanto, as bactérias sozinhas não são nutrição suficiente para Syssomonas: sem presas eucariontes, suas células morrem ou formam cistos de repouso.[2]
Evolução
Como uma linhagem de Holozoa, Syssomonas é um dos muitos grupos de protistas intimamente relacionados a animais e, portanto, é objeto de pesquisa na busca pela origem da multicelularidade animal. As primeiras análises filogenômicas incluindo Syssomonas recuperaram o gênero como o táxon-irmão de Corallochytrium [en]. Juntos, eles compõem o clado Pluriformea, que foi recuperado como o táxon-irmão de Filozoa.[1] Uma hipótese alternativa coloca Pluriformea como o táxon-irmão de Ichthyosporea em um clado conhecido como Teretosporea [en].[2] O seguinte cladograma exibe a posição de Syssomonas entre os membros de Opisthokonta, de acordo com a primeira hipótese:
| Opisthokonta |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Notas
- ↑ Depositado na Coleção de Invertebrados Marinhos, Museu Beaty de Biodiversidade [en], Universidade da Colúmbia Britânica
Referências
- ↑ a b c d e Hehenberger E, Tikhonenkov DV, Kolisko M, del Campo J, Esaulov AS, Mylnikov AP, Keeling PJ (2017). «Novel Predators Reshape Holozoan Phylogeny and Reveal the Presence of a Two-Component Signaling System in the Ancestor of Animals». Current Biology. 27 (13): 2043–2050.e6. Bibcode:2017CBio...27E2043H. PMID 28648822. doi:10.1016/j.cub.2017.06.006

- ↑ a b c d e f g h i j k Tikhonenkov DV, Hehenberger E, Esaulov AS, Belyakova OI, Mazei YA, Mylnikov AP, Keeling PJ (2020). «Insights into the origin of metazoan multicellularity from predatory unicellular relatives of animals». BMC Biology. 18 (1). 39 páginas. PMC 7147346. PMID 32272915. doi:10.1186/s12915-020-0762-1
- ↑ Chen L, Xiao S, Pang K, Zhou C, Yuan X (setembro de 2014). «Cell differentiation and germ–soma separation in Ediacaran animal embryo-like fossils». Nature. 516 (7530): 238–241. Bibcode:2014Natur.516..238C. PMID 25252979. doi:10.1038/nature13766
- ↑ Brown MW, Silberman JD, Spiegel FW (abril de 2011). «"Slime molds" among the Tubulinea (Amoebozoa): molecular systematics and taxonomy of Copromyxa». Protist. 162 (2): 277–287. PMID 21112814. doi:10.1016/j.protis.2010.09.003
- ↑ Brown MW, Silberman JD, Spiegel FW (maio de 2012). «A contemporary evaluation of the acrasids (Acrasidae, Heterolobosea, Excavata)». European Journal of Protistology. 48 (2): 103–123. PMID 22154141. doi:10.1016/j.ejop.2011.10.001
- ↑ Lasek-Nesselquist E, Katz LA (setembro–outubro de 2001). «Phylogenetic position of Sorogena stoianovitchae and relationships within the class Colpodea (Ciliophora) based on SSU rDNA sequences». Journal of Eukaryotic Microbiology. 48 (5): 604–607. PMID 11596926. doi:10.1111/j.1550-7408.2001.tb00197.x
- ↑ Brown MW, Kolisko M, Silberman JD, Roger AJ (2012). «Aggregative Multicellularity Evolved Independently in the Eukaryotic Supergroup Rhizaria». Current Biology. 22 (12): 1123–1127. Bibcode:2012CBio...22.1123B. PMID 22608512. doi:10.1016/j.cub.2012.04.021
- ↑ Schaap P, Winckler T, Nelson M, Alvarez-Curto E, Elgie B, Hagiwara H, et al. (outubro de 2006). «Molecular phylogeny and evolution of morphology in the social amoebas». Science. 314 (5799): 661–663. Bibcode:2006Sci...314..661S. PMC 2173941. PMID 17068267. doi:10.1126/science.1130670