Glyptotherium
Glyptotherium
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Zancliano - Pleistoceno Superior ~3,9–0,015 Ma | |||||||||||||||
| Classificação científica | |||||||||||||||
| |||||||||||||||
| Espécie-tipo | |||||||||||||||
| Distribuição geográfica | |||||||||||||||
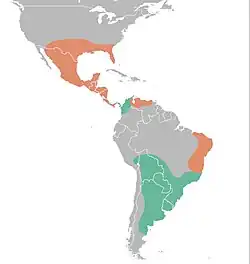 Distribuição de Glyptotherium (laranja) em comparação com Glyptodon (verde) - Américas.
| |||||||||||||||
| Outras espécies | |||||||||||||||
| |||||||||||||||
| Sinónimos | |||||||||||||||
Sinônimos de G. texanum
Sinônimos de G. cylindricum
| |||||||||||||||

Glyptotherium (do grego antigo para "besta entalhada ou esculpida") é um gênero de gliptodonte (um grupo extinto de tatus herbívoros de grande porte) da família Chlamyphoridae que viveu desde o Zancliano, cerca de 3,9 milhões de anos atrás, até o Pleistoceno Superior, aproximadamente 15.000 anos atrás. Foi amplamente distribuído, vivendo nos Estados Unidos, México, Guatemala, Costa Rica, Honduras, El Salvador, Panamá, Venezuela, Colômbia e Brasil.
Fósseis encontrados no "Mount Blanco" do Plioceno em Llano Estacado, Texas, foram nomeados Glyptotherium texanum pelo paleontólogo americano Henry Fairfield Osborn em 1903. Outra espécie, G. cylindricum, foi nomeada em 1912 pelo caçador de fósseis Barnum Brown com base em um esqueleto parcial desenterrado de depósitos do Pleistoceno em Jalisco, México. As duas espécies diferem em vários aspectos, incluindo a idade: G. texanum é do Plioceno Inferior ao Pleistoceno Inferior, enquanto G. cylindricum é exclusivo do Pleistoceno Superior.
Glyptotherium era um tatu grande, quadrúpede e herbívoro, com uma carapaça blindada composta por centenas de osteodermas interconectados (estruturas na derme compostas de osso). Outras peças de armadura cobriam a cauda e o teto craniano, enquanto pequenos pedaços de armadura em forma de seixos estavam na pele. Glyptotherium crescia até 2 metros de comprimento e 400 kg, tornando-se um dos maiores gliptodontes conhecidos. Glyptotherium é morfologicamente mais semelhante ao grupo Glyptodon, embora difiram em vários aspectos. Glyptotherium é, em média, menor, com uma carapaça mais curta, cauda mais longa e tinha uma distribuição diferente.
A diversidade de gliptodontes diminuiu no Pleistoceno, embora eles tenham atingido o pico de tamanho durante esse período. Glyptotherium é considerado um exemplo da megafauna americana, a maioria da qual agora está extinta, possivelmente eliminada por mudanças climáticas ou caça humana. Glyptotherium era principalmente um pastador, vivendo em pastagens abertas, embora também consumisse frutas e outras plantas. A armadura podia proteger o animal de predadores, incluindo o "tigre de dentes de sabre" Smilodon, o canídeo Borophagus [en] e os "ursos de face curta" (Tremarctinae).
História e filogenia
Na década de 1870, fósseis atribuíveis a Glyptotherium foram encontrados pelos engenheiros Juan N. Cuatáparo e Santiago Ramírez, que coletaram um espécime de gliptodonte de um canal de drenagem perto de Tequixquiac, México. Os fósseis vieram do estágio Rancholabreano, que corresponde à época do Pleistoceno do Período Quaternário.[1][2] Este espécime, um crânio, carapaça quase completa e esqueleto pós-craniano associado, foi a primeira descoberta de um gliptodonte na América do Norte.[1][2][3] Cuatáparo e Ramírez nomearam os fósseis Glyptodon mexicanum em 1875, mas os fósseis foram perdidos.[3][4] Outra espécie de Glyptodon do México foi descrita em 1889, G. nathoristi, por paleontólogos alemães com base em restos de carapaça de localidades do Pleistoceno em Ejutla, Oaxaca.[5][6]
Em 1888, o paleontólogo Edward Drinker Cope descreveu um único osteoderma de carapaça coletado em "Equus Beds" do Pleistoceno Inferior em Condado de Nueces, Texas.[7][1] Cope nomeou este osteoderma como Glyptodon peltaliferus,[8] mas não forneceu uma descrição adequada que seguisse as regras do ICZN, tornando-o um nomen nudum e um sinônimo de G. cylindricum.[1][2] No ano seguinte, Joseph Leidy nomeou Glyptodon floridanus com base em osteodermas de carapaça isolados e peças de armadura caudal, que ele inicialmente referiu a G. peltaliferus,[9] coletados de depósitos do Pleistoceno no Condado de DeSoto, Flórida.[10] Esta espécie é um nomen vanum (nome inválido) e considerada um sinônimo de Glyptotherium cylindricum de acordo com uma revisão do gênero pelos paleontólogos americanos David Gillette e Clayton Ray (1981).[11][1]
_(20485352308).jpg)
Glyptotherium foi nomeado em 1903 com base em fósseis coletados por uma expedição do Museu Americano de História Natural (AMNH) liderada por James Gidley das camadas do Plioceno Inferior da formação Blanco de Llano Estacado, Texas. Eles foram descritos por Henry Fairfield Osborn como um novo gênero e espécie de gliptodonte, Glyptotherium texanum.[7] Os fósseis, que foram depositados no AMNH, consistem em uma carapaça e elementos pós-cranianos associados. Este é um dos poucos esqueletos de G. texanum conhecidos.[7][1] O nome genérico Glyptotherium vem das raízes gregas glyph, que significa "entalhado" ou "esculpido", e therion, que significa "besta", um sufixo comumente usado para mamíferos pré-históricos. A espécie é nomeada após a descoberta do espécime holótipo no Texas.[7]
Outra descoberta veio em 1910, quando, enquanto viajava em Jalisco, México, o caçador de fósseis Barnum Brown coletou uma carapaça dorsal completa e vários fósseis adicionais, incluindo dentes, de um único indivíduo das camadas do Pleistoceno da região.[2][12] O espécime foi enviado ao AMNH, onde foi descrito por Brown em 1912 como um novo gênero e espécie de gliptodonte, Brachyostracon cylindricum; o nome da espécie vem das raízes cylindricum, que significa "cilíndrico", devido à anatomia cilindroide dos pré-molares no holótipo de G. cylindricum.[12] Brown também recombinou Glyptodon mexicanum em Brachyostracon mexicanum, embora Brachyostracon seja um sinônimo de Glyptotherium e B. mexicanum seja um sinônimo de G. cylindricum.[12][1][13]
Em 1923, Oliver Hay nomeou uma nova espécie de Glyptodon, G. rivipacis, com base nos fósseis descritos por Leidy do Condado de DeSoto, Flórida.[14] Esta espécie é agora vista como um nomen nudum e sinônima de G. cylindricum.[1][15] Hay também descreveu fósseis bem preservados, incluindo elementos de crânio e dentes, que foram coletados de camadas Rancholabreanas em Wolfe City e Sinton, Texas, e referidos ao Glyptodon peltaliferus de Cope.[13] Esses fósseis foram desde então referidos a G. cylindricum.[1][13] Além de G. peltaliferus, as espécies nomeadas anteriormente G. mexicanum e G. nathoristi foram sinonimizadas com G. cylindricum.[1][2]
Um terceiro gênero de gliptodonte norte-americano, Boreostracon floridanum, foi estabelecido por George Gaylord Simpson em 1929 com base em vários espécimes isolados desenterrados pelo AMNH de localidades de idade Rancholabreana na Flórida, o holótipo consistindo na porção traseira de uma carapaça recuperada da localidade de Seminole Field no Condado de Pinellas, Flórida.[11] Simpson referiu todos os fósseis previamente descritos da Flórida a B. floridanum e acreditava que nenhum dos fósseis de gliptodontes desenterrados na América do Norte era Glyptodon. Simpson optou por não designar um novo gênero ou espécie para Glyptodon peltaliferus, mas ainda acreditava que os fósseis eram de um gênero separado de gliptodonte.[16] B. floridanum foi sinonimizado com Glyptotherium cylindricum.[1] O interesse em gliptodontes norte-americanos diminuiu após as pesquisas de Gidley, Hay, Simpson e outros, mas alguns paleontólogos ainda referiram incorretamente fósseis do continente a Glyptodon.[13]
Em 1927, muitos fósseis de idade Pleistoceno Inferior foram coletados pela Universidade de Oklahoma de uma localidade em Frederick, Oklahoma, incluindo vários fósseis fragmentários de gliptodontes, équidos, gonfotérios e camelídeos.[17] Os fósseis de gliptodonte foram originalmente referidos a Glyptodon em 1928,[18][19] mas não foram devidamente descritos até 1953.[17] Eles foram descritos como um novo gênero e espécie, Xenoglyptodon fredericensis, em 1953 com base em uma mandíbula inferior parcial e vários dentes.[17] A espécie é agora considerada um sinônimo júnior de G. texanum.[1][13] Após todos esses gêneros norte-americanos serem nomeados, a análise de suas relações entre si não foi conduzida até 1981, quando David Gillette e Clayton Ray publicaram uma monografia sobre gliptodontes norte-americanos. Em sua monografia, eles recombinaram todos os gêneros e espécies previamente nomeados em Glyptotherium e pesquisaram a ecologia, anatomia e distribuição do gênero.[13] G. arizonae, G. floridanum e G. mexicanum foram mantidos como espécies válidas ao lado de G. texanum e G. cylindricum.[13] A sinonímia de G. arizonae com G. texanum[1] e G. floridanum e G. mexicanum com G. cylindricum ocorreu após a descoberta de esqueletos mais completos.[1][20]
No século XXI, centenas de fósseis adicionais foram referidos ao gênero da América Central e do Brasil.[21][22] Estes incluem fósseis previamente referidos a Glyptodon e Hoplophorus, pois muitos fósseis foram atribuídos apressadamente a ambos por paleontólogos do século XIX.[21][23] Um dos espécimes reatribuídos a Glyptotherium, um osteoderma isolado, foi coletado de cavernas carbonáticas de idade Pleistocênica em Lagoa Santa, Minas Gerais, Brasil, por Friedrich von Sellow durante o início do século XIX.[21] Em 1845, o paleontólogo dinamarquês Peter Wilhelm Lund descreveu-o como uma nova espécie de Hoplophorus, H. meyeri.[24][23][21] Lund nomeou incorretamente o táxon, tornando-o um nomen nudum.[21] O osteoderma foi referido a Glyptotherium em 2010, tornando-o o primeiro espécime conhecido de Glyptotherium.[21] Em 2022, uma série de fósseis de G. cylindricum, incluindo crânios (alguns preservando patologias causadas por humanos), foram descritos, coletados de vários locais em Falcón, norte da Venezuela, datados do Pleistoceno Superior.[25]

Filogenia
Glyptotherium é um gênero da subfamília Glyptodontinae, um clado extinto de tatus grandes e completamente blindados que evoluiu pela primeira vez no Eoceno Superior (ca. 33,5 milhões de anos atrás) e foi extinto nas extinções do Pleistoceno Superior.[26] A subfamília Glyptodontinae foi classificada como uma família própria ou até superfamília até 2016, quando DNA antigo foi extraído da carapaça de um gliptodonte de 12.000 anos chamado Doedicurus, e um genoma mitocondrial quase completo foi reconstruído (cobertura de 76x).
Comparações com o DNA de tatus modernos revelaram que os gliptodontes divergiram dos tatus tolipeutíneos e clamiforinos aproximadamente há 33,5 milhões de anos no Eoceno Superior.[26][27] Isso levou à reclassificação deles de sua própria família, Glyptodontidae, para uma subfamília dentro da família existente Chlamyphoridae, renomeada como Glyptodontinae.[27] A seguinte análise filogenética foi conduzida por Frédéric Delsuc e colegas em 2016 e representa a filogenia de Cingulata usando DNA antigo de Doedicurus para determinar a posição dele e de outros gliptodontes:[26][28] A análise filogenética a seguir foi realizada por Frédéric Delsuc e colegas em 2016 e representa a filogenia dos Cingulata usando DNA antigo do Doedicurus para determinar a posição dele e de outros Glyptodonts:[29]
| Cingulata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
A filogenia interna de Glyptodontinae é complexa e está em fluxo. Muitas espécies e gêneros foram estabelecidos com base em material fragmentário ou não diagnóstico que não foi revisado de forma abrangente.[30][23][1] Geralmente é considerada uma família própria, mas análises de DNA a reduziram a uma subfamília com tribos em vez de subfamílias. Uma tribo, Glyptodontini (geralmente rotulada como Glyptodontinae), é um grupo de gliptodontes mais jovens e maiores que evoluíram no Mioceno Médio (ca. 13 milhões de anos atrás) com Boreostemma,[31] mas se dividiram em dois gêneros: Glyptodon e Glyptotherium.[1][32] Geralmente é considerada sua própria família, mas as análises de DNA a reduziram a uma subfamília com tribos em vez de subfamílias. Uma tribo, Glyptodontini (normalmente rotulada como Glyptodontinae) é um grupo de gliptodontes mais jovens e maiores que evoluíram no Mioceno Médio (ca. 13 ma) com o Boreostemma,[33] mas se dividiram em dois gêneros: Glyptodon e Glyptotherium.[1][34] Glyptodontini é frequentemente recuperado como mais basal em relação à maioria dos outros gliptodontes, como Doedicurus, Hoplophorus e Panochthus. Glyptodontini é distinguível de outros grupos, por exemplo, por ter grandes osteodermas tuberculados cônicos ausentes ou presentes apenas na borda caudal (em direção à cauda) na extremidade posterior da carapaça e ornamentação diferente da armadura na carapaça em relação à cauda.[35]
Quando Glyptotherium foi descrito pela primeira vez por Henry Osborn em 1903, foi colocado no nível de família Glyptodontidae, e Osborn afirmou que era mais próximo em aparência e classificação de Panochthus e Neosclerocalyptus (então Sclerocalyptus).[7] Barnum Brown acreditava que Brachyostracon estava em sua própria família de gliptodontes, mas não cunhou nenhum novo nome para essa família. Em vez disso, ele classificou o gênero dentro de Sclerocalyptidae junto com gliptodontes sul-americanos como Panochthus, Neosclerocalyptus e Plohophorus.[12] George Gaylord Simpson acreditava que Boreostracon era um parente próximo de Glyptodon.[16] Xenoglyptodon foi colocado como um gliptodonte próximo aos outros gêneros norte-americanos por Grayson Meade em 1953, mas ele não afirmou sua relação com gêneros sul-americanos.[17] O consenso atual é que Glyptotherium pertence à tribo Glyptodontini e possui duas espécies em seu gênero.[1][20] Abaixo está a análise filogenética conduzida por Cuadrelli et al., 2020, de Glyptodontinae, com Glyptodontidae como uma família em vez de subfamília, que se concentra em gliptodontes avançados:
| Chlamyphoridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Descrição

Como seu parente vivo, o tatu, o Glyptotherium tinha uma carapaça que cobria todo o tronco, com uma armadura menor cobrindo também o teto do crânio da cabeça. A carapaça (parte superior) do Glyptotherium era composta de 1.800 ou mais pequenos osteodermas hexagonais em cada indivíduo.[36] O esqueleto axial dos gliptodontes mostra uma fusão extensa na coluna vertebral e a pélvis (osso do quadril) é fundida à carapaça, tornando a pélvis totalmente imóvel.[13] Glyptotherium era muito gravitacional e tinha membros curtos que são muito semelhantes aos de outros gliptodontes. As caudas grandes dos gliptodontes provavelmente serviam como contrapeso para o resto do corpo. A armadura caudal dos Glyptotherium terminava em um tubo composto de 2 a 3 tubos fundidos,[1] em contraste com os dos gliptodontes de cauda de macaco da América do Sul.[37] Os dígitos do Glyptotherium são muito robustos e adaptados para suportar peso, com alguns preservando grandes bainhas de garras que tinham uma morfologia intermediária entre garras e cascos.[13]
Os pesos e tamanhos do Glyptotherium variam, mas o G. texanum era menor e mais leve do que a espécie posterior, o G. cylindricum. Um espécime de G. cylindricum foi estimado em 350 a 380 kg, em comparação com 457 kg de seu parente Glyptodon reticulatus.[25] As maiores estimativas da massa do Glyptotherium são baseadas em espécimes adultos do Brasil e do Arizona, com uma estimativa de um espécime de G. cylindricum em 710 kg e um indivíduo de G. texanum em 1.165 kg. Um espécime de G. cylindricum, AMNH 15548, preserva um comprimento de carapaça de 169 cm em comparação com apenas 140 cm do MSM P4464, um espécime de G. texanum.[1]
_and_Taima-Taima_sites_(C%E2%80%93I)_are_shown.webp.png)
G. texanum é distinto de G. cylindricum na configuração dos osteodermas na carapaça dorsal, pois as figuras centrais de G. texanum são planas ou ligeiramente convexas e muito grandes, enquanto as de G. cylindricum são planas ou côncavas e muito menores.[1][38] Em ambas as espécies, as figuras centrais dos osteodermas aumentam de tamanho em direção às margens da carapaça.[1][20]
Crânio e mandíbula
A dentição dos gliptodontes possui todos os dentes hipsodontes (dentes de coroa alta adaptados para pastagem), dentes molares, que estão entre os de coroa proporcionalmente mais alta conhecidos em mamíferos terrestres.[39] Os crânios dos gliptodontes possuem várias características únicas; o maxilar e o palatino são ampliados verticalmente para criar espaço para os molares, enquanto a caixa craniana é braquicefálica, curta e plana.[40] A armadura dérmica não se restringia à carapaça e à cauda, pois o teto craniano [en] era protegido por um "escudo cefálico" feito de osteodermas.[25][41] Apenas um crânio completo é conhecido de G. texanum, oferecendo uma perspectiva limitada sobre sua anatomia.[42][43]
Os zigomas de Glyptotherium são estreitos, delgados, quase paralelos e próximos ao plano sagital na vista anterior. Em Glyptodon, os zigomas são mais largos, mais robustos e mais divergentes em vez de paralelos. Glyptotherium e outros gliptodontes preservam grandes passagens nasais e seios paranasais que podem ter tido narinas adaptadas para respirar nos climas frios e áridos das Américas durante o Pleistoceno.[44] Alguns paleontólogos propuseram que Glyptotherium e alguns gliptodontes também tinham uma probóscide ou focinho grande semelhante aos de elefantes e antas,[45] mas poucos aceitaram essa hipótese.[1][25] Os forames infraorbitais [en] (aberturas no maxilar) são estreitos e não visíveis na vista anterior em Glyptotherium, mas são largos e claramente visíveis em Glyptodon.[1]
Na vista lateral, a altura dorso-ventral entre o teto craniano e o plano palatal em Glyptodon diminui anteriormente, ao contrário de Glyptotherium; a ponta nasal está em um plano mais baixo em relação ao arco zigomático em Glyptodon, mas em Glyptotherium é mais alta que o plano do arco zigomático. Em Glyptotherium, o perfil lateral oclusal é ligeiramente curvo, enquanto em Glyptodon é fortemente curvo. Em Glyptodon, o primeiro molariforme (abreviado como mf1) é distintamente trilobado (com três lóbulos) tanto lingual quanto labialmente, quase tão trilobado quanto o mf2; ao contrário, Glyptotherium mostra uma trilobação muito baixa do mf1, que é elíptico em seção transversal, o mf2 é fracamente trilobado, e o mf3 é trilobado. Em ambos os gêneros, os mf4 a mf8 são totalmente trilobados e serialmente idênticos.[1]
As mandíbulas de Glyptotherium e Glyptodon são muito semelhantes, mas a mandíbula de Glyptotherium é cerca de 10% menor. O ângulo entre o plano oclusal e a margem anterior do ramo ascendente é aproximadamente 60° em Glyptotherium, enquanto é 65° em Glyptodon. A margem ventral do ramo horizontal é mais côncava em Glyptodon do que em Glyptotherium. A área da sínfise é muito esticada antero-posteriormente em Glyptotherium em comparação com Glyptodon. O mf1 é elipsoidal em Glyptotherium e o mf2 é submolariforme, enquanto em Glyptodon ambos os dentes são trilobados.[1]
Carapaça e osteodermas

A carapaça de Glyptotherium era mais curta que a de seu parente Glyptodon, mas muito mais alongada que a de Boreostemma. O ponto mais alto da carapaça estava no centro da linha média, enquanto o de Glyptodon era ligeiramente deslocado. A carapaça de Glyptotherium era fortemente arqueada, com uma região pré-ilíaca (quadril) convexa e pós-ilíaca côncava, dando-lhe uma aparência de sela sobre a cauda.[1][31] Em Glyptodon, a altura da carapaça de cima para baixo representa 60% de seu comprimento total, enquanto em Glyptotherium é mais alta, cerca de 70%. A margem ventral da carapaça em Glyptotherium é mais retangular e menos convexa que em Glyptodon. Em Glyptotherium, os osteodermas nas áreas antero-laterais da carapaça são menos anquilosados que em Glyptodon, sugerindo que as regiões antero-laterais da carapaça do primeiro eram mais flexíveis. Os osteodermas da abertura caudal são mais cônicos em Glyptodon e mais arredondados em Glyptotherium, embora neste último a anatomia dos osteodermas da abertura caudal varie por sexo[20] enquanto em Glyptodon varia por idade.[46] Um exame de espécimes de Glyptotherium do Blancan do Arizona sugere que, ao longo do tempo, a carapaça de Glyptotherium tornou-se mais robusta e com uma concha mais espessa, correspondendo a uma construção geral maior.[47]
Embora frequentemente usados para diferenciar os dois táxons, Glyptotherium e Glyptodon têm morfologias de osteodermas muito semelhantes que mal diferem. Ambos os gêneros têm osteodermas espessos em comparação com os de muitos gliptodontes sul-americanos como Hoplophorus e Neosclerocalyptus, mas Glyptotherium sempre preserva um padrão de "roseta", onde a figura central do osteoderma é cercada por uma fileira de figuras periféricas. Alguns espécimes de Glyptodon preservam essas "rosetas", mas outros não as possuem. Os sulcos centrais e radiais são mais profundos e largos em Glyptodon (ca. 4-6 mm) do que em Glyptotherium (ca. 1-2.4 mm).[1][20] Notavelmente, os osteodermas de Glyptotherium preservam pequenos espaços para folículos pilosos nos sulcos que indicam que Glyptotherium tinha uma carapaça "peluda" com pelos saindo. O número de folículos varia entre idades e a área da carapaça, com juvenis tendo mais folículos que adultos, e menos folículos são conhecidos nas porções laterais, caudais e traseiras da carapaça.[48][20]
Anéis caudais
.jpg)
Glyptotherium é um gliptodonte, o que significa que sua armadura caudal é composta por uma série de anéis caudais terminando em um tubo caudal curto, mas a morfologia difere entre Glyptotherium, Glyptodon e Boreostemma. No geral, Boreostemma preserva uma armadura caudal mais semelhante a Glyptotherium do que a Glyptodon. A armadura caudal é mais longa em Glyptotherium do que em Glyptodon, com um espécime de G. texanum (UMMP 34 826) preservando um conjunto de armadura caudal de 1 m de comprimento. Em Glyptotherium, o comprimento da armadura caudal representa cerca de 50% do comprimento total da carapaça dorsal, enquanto em Glyptodon, esse valor é menor, cerca de 30-40%.
Glyptodon tem oito a nove anéis caudais completos mais um tubo caudal, mas Glyptotherium preserva um anel caudal incompleto além dos oito a nove anéis caudais completos e tubo caudal. Em ambos os gêneros, cada anel caudal é composto por duas ou três fileiras transversais de osteodermas anquilosados, onde a fileira mais distal de osteodermas mostra uma morfologia cônica mais ou menos desenvolvida. Em Glyptotherium, em alguns espécimes (por exemplo, AMNH 95,737), há um baixo número de osteodermas cônicos (geralmente dois). Isso é diferente de Glyptodon, no qual a maioria dos osteodermas da fileira distal (até 12) apresenta uma morfologia cônica clara. O tubo caudal terminal é mais curto em Glyptodon. Em Glyptotherium, o tubo caudal é composto por dois a três anéis anquilosados, enquanto em Glyptodon ele tem apenas dois anéis anquilosados. Em Glyptotherium, este tubo caudal representa cerca de 20% do comprimento total da armadura caudal, enquanto em Glyptodon, essa estrutura representa 13% do comprimento total.[1]
Paleobiologia
Postura
Diversas interpretações da postura dos gliptodontes foram feitas,[49] inicialmente pelo paleontólogo britânico Richard Owen em 1841, utilizando anatomia comparativa.[50] Owen teorizou que as falanges eram adaptadas para suportar peso devido à sua fisiologia curta e larga, além das evidências fornecidas pelo esqueleto pós-craniano.[50] Também foi proposto que uma postura ereta era possível para os gliptodontes, primeiramente por Sénéchal (1865), que afirmou que a cauda poderia servir como equilíbrio para a metade frontal do corpo, além de ser um método de suporte para as pernas.[51][49] Medições lineares posteriores forneceram insights sobre essa hipótese, constatando que o bipedismo seria possível.[52][53] A articulação patelar com o fêmur sugere rotação do crus durante a extensão do joelho e possivelmente até mesmo o travamento do joelho.[54]

Alimentação e dieta
Tradicionalmente, acredita-se que Glyptotherium consumia ervas úmidas de zonas ripárias.[55] O gênero era principalmente um pastador, mas também tinha uma dieta mista de plantas C3 e C4, com base em análises isotópicas de espécimes dentários recuperados da localidade de Cedral, no Pleistoceno Superior, em San Luis Potosí, México.[56][57] A localidade preserva plantas C4 das famílias Poacea, Amaranthacea e Quenopodiacea,[58] o que significa que elas eram possíveis fontes de alimento para Glyptotherium.[56] Cedral, especificamente, era uma área com fontes termais e pastagens abertas próximas, sugerindo que Glyptotherium se alimentava em pastagens próximas a fontes de água, semelhante aos hábitos alimentares das modernas capivaras.[45][56] Análises isotópicas adicionais de Glyptotherium e da preguiça gigante Eremotherium encontraram níveis isotópicos semelhantes aos do hipopótamo atual, indicando que eram herbívoros semi-aquáticos que se alimentavam de plantas aquáticas. Estudos adicionais de espécimes dentários do leste do Brasil sugerem que Glyptotherium eram pastadores em habitats tropicais a subtropicais úmidos e baixos ao longo de rios ou fontes de água, apoiando a hipótese de Glyptotherium semi-aquático.[22][59] Evidências isotópicas adicionais do Brasil sugerem que frutas também faziam parte da dieta de Glyptotherium, embora representassem apenas cerca de 20% do total.[60] Glyptotherium e todos os outros gliptodontes tinham dentes hipsofontes, dentes de coroa alta com superfícies ásperas e planas adaptadas para triturar e esmagar, que eram adaptados para quebrar materiais fibrosos e ásperos como gramíneas.[61][45] Essa dieta de Glyptotherium contrasta com as supostas para seus parentes Pampateriídeos, que foram considerados insetívoros ou pastadores.[56][62] Como a maioria dos outros parentes, os gliptodontes tinham requisitos energéticos mais baixos do que a maioria dos outros mamíferos. Eles podiam sobreviver com taxas de ingestão menores do que outros herbívoros com massa semelhante.[63]
Habilidades de escavação
Muitas espécies atuais de tatus têm capacidades de escavação, com grandes garras adaptadas para raspar terra para fazer tocas ou forragear por comida no subsolo.[64][65] Grande parte de sua dieta consiste em insetos e outros invertebrados que vivem no solo,[66] em contraste com as dietas herbívoras de Glyptotherium.[67] Sendo da família dos tatus, as capacidades fossoriais dos gliptodontes foram pesquisadas em várias ocasiões.[49] Owen (1841) opôs-se a essa ideia, embora tenha recebido críticas de Nodot (1856) e Sénéchal (1865), que acreditavam que a escavação era possível para o gênero.[49][50] No entanto, a evolução de uma carapaça rígida em oposição a uma flexível nos tatus atuais, bem como uma crista deltoide pouco desenvolvida no úmero (osso do braço superior), forneceu evidências contra hipóteses fossoriais. O cotovelo tinha uma grande amplitude de movimento, como nos cingulados escavadores, mas isso é mais provável devido a adaptações de tamanho.[68][49]
Ontogenia
Espécimes juvenis e adultos de G. texanum encontrados em localidades do Blancan no Arizona preservam uma série de crescimento quase completa, uma das poucas conhecidas em gliptodontes.[20] Os dentes de Glyptotherium preservam hipodontia, em que os dentes não param de crescer.[20] Nos juvenis, o contato osteoderma-a-osteoderma totalmente articulado já é conhecido, mesmo em sua idade.[20] O crescimento dos osteodermas continuou de juvenis a subadultos e cessou quando os osteodermas se tornaram anquilosados (fundidos).[16][20] O perfil lateral das carapaças juvenis é suavemente convexo, transformando-se nas carapaças altamente arqueadas dos adultos. Outra mudança ontogenética está nos sulcos e figuras periféricas dos osteodermas; as figuras centrais são as maiores em relação aos sulcos em recém-nascidos e juvenis, mas essa proporção diminui em adultos. Em G. cylindricum, no entanto, os osteodermas crescem muito mais rápido e os sulcos são muito menores. Os osteodermas também são relativamente mais espessos em indivíduos juvenis de Glyptotherium em comparação com adultos.[20]
Dimorfismo sexual
Indivíduos de G. texanum encontrados em localidades do Blancan no Arizona preservam dimorfismo sexual entre machos e fêmeas. A abertura caudal, grandes osteodermas cônicos que protegem a base da cauda, de machos e fêmeas difere, pois os osteodermas marginais dos machos são muito mais cônicos e convexos do que os das fêmeas. Mesmo nas carapaças de recém-nascidos de Glyptotherium, os osteodermas marginais são cônicos ou planos, permitindo determinar seu sexo.[20]
Osteodermas e proteção

Os osteodermas de Glyptotherium são feitos de um núcleo trabecular esponjoso entre duas camadas compactas. Cada osteoderma hexagonal é unido aos osteodermas adjacentes com suturas, criando uma carapaça grande e robusta semelhante às de animais modernos. No entanto, as carapaças dos gliptodontes como Glyptotherium eram muito menos flexíveis do que as dos tatus modernos.[69] O núcleo trabecular era composto por suportes usados para suporte com uma espessura média de 0,25 mm, esses suportes formando o suporte central do osteoderma. Análises mecânicas revelaram que áreas de carga menores, representando objetos mais afiados, causam tensões mais altas do que aquelas causadas por objetos grandes e rombudos. Isso pode ser entendido como a estrutura natural evoluída para resistir a impactos rombudos de objetos grandes, como clavas caudais, e não como proteção contra objetos afiados, como dentes.[38] Isso apresenta mais evidências para a teoria de que os gliptodontes usavam suas carapaças para combate intraespecífico usando clavas caudais, além de defesa.[38]
Doenças e patologias
Um esqueleto pós-craniano parcial de G. cylindricum encontrado no nordeste do Brasil, que incluía membros parciais, preservou três tipos diferentes de artrite: um tipo de artrite inflamatória (espondiloartropatia), um tipo de artrite cristalina (doença por deposição de pirofosfato de cálcio) e uma artrite proliferativa (osteoartrite). A espondiloartropatia foi a mais prevalente, com erosão óssea na ulna direita, rádios direito e esquerdo, um fêmur esquerdo e ambas as tíbias-fíbulas direitas e esquerdas, e está associada à esclerose óssea na ulna e rádio direitos. A doença por deposição de pirofosfato de cálcio está presente na superfície articular da patela esquerda, e a osteoartrite é representada por osteófitos em alguns dos membros posteriores esquerdos.[70] Em vários osteodermas também encontrados no Brasil, foram encontrados casos de ectoparasitismo, lesões e crescimentos estranhos, algumas dessas infecções provavelmente causadas por pulgas.[71]
Paleoecologia
Glyptotherium era principalmente um pastador em áreas campestres florestadas e savanas arbóreas, embora possam ter preferido pastagens próximas a fontes de água com base em localidades fósseis do México.[56] Devido à sua ampla distribuição, a paleoecologia de Glyptotherium pode ter variado entre regiões e espécies.[22][56]

Na América do Norte, Glyptotherium é conhecido por duas espécies diferentes que viveram em três intervalos diferentes, o Blancan [en], Irvingtoniano e Rancholabreano, nos Estados Unidos e México.[1] No entanto, Glyptotherium não foi encontrado em drenagens do Pacífico nos Estados Unidos ou a oeste do rio Colorado,[1] estando presente principalmente na Planície Atlântica [en] e nas Planícies Interiores.[72] Durante o Blancan, G. texanum coexistiu com muitos gêneros endêmicos da América do Norte, pois Beríngia ainda não havia se formado. Por causa disso, a fauna do Blancan contrastava fortemente com a fauna do Pleistoceno subsequente. Os estratos da idade Blancana do oeste do Texas, Novo México e Arizona preservam proboscídeos gomfotéridos Stegomastodon [en] e Cuvieronius, equídeos representados por pastadores como Nannipus e Equus. Outros xenartros também são conhecidos, como a preguiça terrestre megaloniquídea Megalonyx e o milodontídeo Paramylodon [en].
Fósseis de pequenos mamíferos também foram desenterrados, como os insetívoros Hesperoscalops e Sorex. Um esquilo terrestre gigante, Paenemarmota, também é conhecido do Blancan.[73] Os carnívoros incluem o incomum cão "triturador de ossos" Borophagus [en][74] e a "hiena caçadora" Chasmaporthetes [en],[75][76] além do tigre "dente-de-sabre" Smilodon gracilis.[77] Alguns fósseis isolados de aves também foram encontrados, consistindo de abutres, falcões e possivelmente corvídeos.[78][79][80] Fósseis da Guatemala foram encontrados em grandes altitudes, mostrando que Glyptotherium era altamente adaptável e podia viver em uma variedade de ambientes.[81] Isso é apoiado pela dieta generalista de Glyptotherium e descobertas de fósseis em ambientes tropicais, subtropicais, florestais e até semi-aquáticos. No estado mexicano de Yucatán, fósseis de Glyptotherium e da preguiça terrestre Paramylodon são conhecidos de áreas ricas em água, como florestas ripárias e pântanos, em oposição a pastagens abertas.[82]
Na região intertropical brasileira (RIB) no leste do Brasil, Glyptotherium era um pastador misto em savanas arbóreas, áreas campestres tropicais e outras áreas abertas próximas a fontes de água. Grandes mamíferos mesoherbívoros na RIB eram amplamente distribuídos e diversos, incluindo os toxodontídeos semelhantes a rinocerontes, como Toxodon platensis e Piauhytherium [en], o macraucheniídeo litopterno Xenorhinotherium e equídeos como Hippidion principale [en] e Equus neogeus [en]. Toxodontídeos também eram grandes alimentadores mistos e viviam em áreas florestadas, enquanto os equídeos eram quase inteiramente pastadores.
Outros fósseis de xenartros também estão presentes na área, de várias famílias diferentes, como a preguiça terrestre gigante megateriídea Eremotherium, os escelidoteriídeos Catonyx [en] e Valgipes [en], os milodontídeos Glossotherium, Ocnotherium [en] e Mylodonopsis [en]. Preguiças terrestres menores, como os megaloniquídeos Ahytherium [en] e Australonyx [en] e o notroteriídeo Nothrotherium também foram encontrados na área. Eremotherium era um generalista, enquanto Nothrotherium era um especialista em árvores em florestas de baixa densidade, e Valgipes era um intermediário dos dois que vivia em savanas arbóreas. Outros gliptodontes e cingulados como o gliptodonte pastador Panochthus e os pampateriídeos onívoros Pampatherium e Holmesina estavam presentes nos campos abertos. Uma espécie de proboscídeo também foi encontrada na RIB, Notiomastodon platensis, também era um pastador misto nas áreas campestres. Carnívoros incluíam alguns dos maiores mamíferos carnívoros conhecidos, como o felídeo gigante Smilodon populator e o urso Arctotherium wingei.[83][59]
Vários táxons atuais também são conhecidos da RIB, como guanacos, tamanduá-bandeira, queixadas e jaritatacas.[84] Dois tipos de mamíferos comedores de caranguejos também são conhecidos da região, o guaxinim mão-pelada e o cachorro-do-mato, indicando que caranguejos também estavam presentes na região.[84] O ambiente da RIB é incerto, pois havia várias espécies que eram pastadoras, mas a presença dos fósseis de macacos arbóreos Protopithecus [en] e Caipora [en] na área causa confusão sobre o paleoambiente da região. A maior parte do Brasil era considerada coberta por vegetação de cerrado tropical aberto durante o Pleistoceno Superior, mas se Protopithecus e Caipora eram arbóreos, sua presença sugere que a região pode ter sustentado uma floresta fechada densa durante o mesmo período.[84][85] É possível que a região tenha alternado entre savana aberta seca e floresta úmida fechada ao longo das mudanças climáticas do Pleistoceno Superior.[86]
Grande Intercâmbio Americano
A América do Sul, o continente onde os gliptodontes se originaram, ficou isolada após a separação do continente Gondwana no final da era Mesozoica.[87] Esse período de separação do resto dos continentes da Terra levou a uma era de evolução mamífera única, com o domínio de grupos como marsupiais, xenartros e notoungulados em contraste com a fauna mamífera da América do Norte. Os marsupiais provavelmente chegaram à América do Sul antes de sua separação do resto do Gondwana no Cretáceo Superior ou Paleogeno, embora a origem de ordens mamíferas como Xenarthra e Notoungulata que acabaram no continente permaneça um mistério.[88] Houve vários movimentos de mamíferos externos para a América do Sul antes da formação do istmo do Panamá, como com primatas e roedores que podem ter chegado ao continente a partir da África e o movimento de morcegos por voo.[89][90]
Quanto à fauna da América do Norte, grupos contemporâneos como canídeos, felídeos, ursídeos, tapirídeos, antilocaprídeos e equídeos povoavam a região, além de famílias extintas como gomfotéridos, anficionídeos e mamutídeos [en].[91][87] O grande intercâmbio americano não entrou em sua maior fase até que o istmo do Panamá se formasse há 2,7 milhões de anos durante o estágio Blancan do Plioceno.[92][93][94] Isso intensificou o movimento de gliptodontes, preguiças terrestres, capivaras, pampateres, aves do terror e marsupiais para a América do Norte via a rota da América Central e uma migração inversa de ungulados, proboscídeos, felídeos, canídeos e muitos outros grupos de megafauna para a América do Sul.[95] O período após a formação do istmo testemunhou a extinção ou extirpação de muitos grupos, incluindo as aves do terror sul-americanas, toxodontes, macraucheniídeos, pampateres, preguiças terrestres e gliptodontes.[96][97]
O próprio Glyptotherium fez parte desse intercâmbio, evoluindo no Blancan do México e dos Estados Unidos após a formação do istmo e sua imigração.[1][95] O isolamento geográfico de Glyptotherium em combinação com adaptações aos diferentes ambientes da América do Norte levou Glyptotherium a se separar de outros gliptodontíneos.[1] Embora G. texanum tenha evoluído na América do Norte para G. cylindricum no Rancholabreano, ele emigrou para o sul para a América Central e partes do norte da América do Sul cerca de 20.000-15.000 anos antes do presente (AP).[25][21][98] Isso também está conectado à segregação ecológica, com barreiras montanhosas na Colômbia possivelmente separando Glyptodon e Glyptotherium.[32] Subsequentemente, Glyptodon viveu principalmente em sítios andinos e costeiros, com Glyptotherium sendo conhecido de depósitos de pastagens e áreas florestais leves de terras baixas próximas a áreas aquáticas, motivando sua dispersão para as regiões tropicais da Venezuela e leste do Brasil.[94][22][99]
A reentrada de um grupo na América do Sul a partir da América do Norte também foi observada na família cingulada relacionada Pampatheriidae,[93] possivelmente auxiliada por amplas terras baixas costeiras descobertas durante períodos glaciais nas costas do Caribe e Atlântico, permitindo a migração entre Flórida, México, América Central e/ou norte da América do Sul.[94][100] Um estudo de 2025 analisando os possíveis padrões de migração de Glyptotherium através do istmo do Panamá para a América do Norte concluiu que, com base nos modelos de nicho desenvolvidos pelo estudo, Glyptotherium provavelmente tomou um "caminho alto" para a América do Norte. Esse "caminho alto" estava localizado no noroeste da América do Sul através dos Andes e Colômbia, em vez de uma rota de terras baixas amazônicas. Isso é consistente com a ecologia de pastagem e as preferências climáticas de Glyptotherium.[101]


Predação
Smilodon pode ter ocasionalmente predado Glyptotherium, com base em um crânio de um indivíduo de G. texanum recuperado de depósitos do Pleistoceno no Arizona, que apresenta marcas de perfuração elípticas distintas que melhor correspondem às de um gato machairodonte, indicando que o predador conseguiu morder o crânio através do escudo cefálico blindado.[102] O Glyptotherium em questão era um juvenil, com um escudo cefálico ainda em desenvolvimento, tornando-o muito mais vulnerável ao ataque do gato.[103]
Brandes hipotetizou que a evolução da armadura espessa dos gliptodontes e dos caninos longos dos machairodontes foi um exemplo de coevolução,[104] mas Birger Bohlin argumentou em 1940 que os caninos eram frágeis demais para causar danos contra a armadura dos gliptodontes.[105][46] No entanto, a evolução de estruturas de proteção acessórias pode ter sido uma resposta à chegada de Smilodon e Arctotherium.[46]
Em 2017, uma ulna direita de um Glyptotherium adulto, coletada de estratos do Pleistoceno Superior no estado do Rio Grande do Norte, Brasil, foi descrita, apresentando várias marcas de roedura de uma nova icnoespécie de Machichnus, M. fatimae, que podem ter sido causadas por um juvenil da espécie canídea Protocyon troglodytes ou um indivíduo adulto de Cerdocyon thous.[106]
Relação com humanos
_with_human-caused_pathologies.webp.png)
O primeiro relato de possível consumo humano ou interação com Glyptotherium ou seus fósseis veio em 1958, quando vários osteodermas que possivelmente foram consumidos por humanos foram descritos do sítio de Clovis em Lewisville, Texas.[107][108] No entanto, essa ideia de consumo humano tem pouca evidência para apoiá-la.[25] Em 2022, uma série de fósseis de G. cylindricum, incluindo crânios, foi descrita, coletada de vários sítios em Falcón, norte da Venezuela, datados do Pleistoceno Superior.[25] Essas descobertas foram notáveis não apenas por preservarem crânios, mas quatro deles exibiam fraturas na região fronto-parietal, um padrão em todos os crânios. Evidências visuais e de tomografia computadorizada (TC) indicam que essas foram provavelmente causadas por um efeito mecânico por percussão direta, muito provavelmente um golpe com um machado de pedra ou clava, que causou a fragmentação dos ossos na região para os tecidos moles internos do crânio. Apesar do fato de que os crânios estavam completos e não apresentavam sinais de distorção tafonômica ou transporte, eles frequentemente não tinham suas mandíbulas. As mandíbulas podem ter sido removidas para que "caçadores" acessassem e consumissem músculos mastigatórios e a língua.[109][25]
A coexistência de humanos caçadores-coletores primitivos e gliptodontes na América do Sul foi primeiramente hipotetizada em 1881 com base em descobertas fósseis dos pampas argentinos,[110] e muitas descobertas fósseis do Pleistoceno Superior foram desenterradas desde então, exibindo registros de predação humana em gliptodontes. Durante esse período, uma ampla gama de xenartros que habitavam os pampas foi caçada por humanos, com evidências demonstrando que o pequeno gliptodonte Neosclerocalyptus de 300-400 kg,[111] o tatu Eutatus e o gigantesco gliptodonte Doedicurus de 2 toneladas, o maior gliptodonte conhecido, foram caçados.[112] O único outro registro de predação humana fora dos pampas foi uma carapaça parcial, também encontrada na Venezuela, que foi eviscerada por humanos. As descobertas nas localidades de Falcón mostraram os primeiros sinais de caça humana nos crânios de gliptodontes, mas Glyptotherium também era mais indefeso do que gliptodontes como Doedicurus.[25]
Distribuição
Glyptotherium é o único gliptodonte conhecido da América do Norte e é conhecido de várias regiões do continente de diferentes períodos. Durante o estágio Blancan do Plioceno Inferior, G. texanum habitava apenas o centro do México, com base na descoberta de um único osteoderma de G. texanum dos estratos do início do Plioceno de Guanajuato, centro do México, datando de aproximadamente 3,9 milhões de anos atrás.[1] Nos estágios Blancan-Irvingtoniano do Pleistoceno Inferior, fósseis de G. texanum são conhecidos da maior parte do México, bem como dos estados americanos de Arizona, Texas, Oklahoma, Flórida e possivelmente Carolina do Sul.[1][113] No Rancholabreano do Pleistoceno Superior, G. cylindricum evoluiu de G. texanum e seus fósseis foram desenterrados do norte da Venezuela, leste do Brasil, América Central, México e os estados americanos de Texas, Louisiana, Flórida e Carolina do Sul.[1][113][114][55] Pegadas também foram recuperadas de Virgínia.[115] Fósseis de Glyptotherium do Plioceno Inferior não foram encontrados na América Central, mas é provável que G. texanum tenha habitado a área durante o Grande Intercâmbio Biótico Americano. Fósseis de gliptodontes do Irvingtoniano médio-tardio não são conhecidos dos Estados Unidos, criando uma "lacuna de gliptodontes" no registro fóssil dos Estados Unidos.[1] No entanto, Glyptotherium é registrado durante essa "pausa" na América Central, sugerindo uma possível retração de Glyptotherium para áreas ao sul durante tempos glaciais.[98]

Fósseis de Glyptotherium foram coletados na América Central na Guatemala,[81] Costa Rica,[116] Honduras,[117] El Salvador,[118] e Panamá.[119] Os fósseis da América Central são geralmente isolados e fragmentários, sendo a maioria osteodermas ou molares isolados.[1][120] Em 2023, um esqueleto associado de G. cylindricum, incluindo crânio e elementos dos membros, da Guatemala foi descrito, o espécime mais completo da região.[121] É mais provável que as primeiras populações de Glyptotherium tenham começado na América Central durante o Grande Intercâmbio Americano no Plioceno Superior, com base em sua paleobiogeografia.[1][20] Fósseis de Glyptotherium da América Central são às vezes classificados como uma espécie indeterminada,[22][21] mas a maioria é classificada em G. cylindricum ou seus sinônimos.[81][116] Essa classificação também é baseada na idade dos fósseis, pois a idade dos fósseis de G. texanum varia do Plioceno ao Pleistoceno Inferior, enquanto a idade dos fósseis de G. cylindricum é restrita ao Pleistoceno Superior.[1][122] Embora comumente considerado um gênero exclusivamente norte-americano,[122][21][1] fósseis de Glyptotherium do norte da América do Sul em áreas como Brasil e Venezuela foram descobertos.[21][25] Os fósseis da América do Sul são geralmente apenas osteodermas ou anéis caudais e às vezes são indeterminados em nível de espécie, mas são mais provavelmente de G. cylindricum.[1][21][25]
Extinção
A cronologia das extinções da megafauna (como Glyptotherium) do Pleistoceno Superior tem sido debatida.[72] Nos Estados Unidos, a última datação direta confiável por radiocarbono para Glyptotherium é 23.230 ± 490 AP, da caverna Laubach, Texas.[123][72] Glyptotherium agrupa-se com Eremotherium, Holmesina e Paramylodon como tendo datas finais confiáveis antes do fim do último máximo glacial da América do Norte.[124] No entanto, análises estatísticas sugerem que uma sobrevivência posterior até o Pleistoceno terminal dos Estados Unidos é possível, com base em vieses de amostragem associados a fauna incomum e a falta de datas confiáveis da planície Atlântica úmida devido à má preservação.[72]
Na América do Sul, restos queimados de Glyptotherium foram datados imprecisamente entre 16.375 ± 400 AP e 14.300 ± 500 AP por radiocarbono em Muaco, Venezuela,[25] com técnicas semelhantes datando um espécime de Glyptotherium de Taima-Taima para 12.580 ± 60 AP por radiocarbono,[125][126] embora uma data mínima de todo o conjunto (~15.780 cal. AP, 12.980 ± 85 AP por radiocarbono) seja mais recente.[25] Como com outras megafaunas extintas do Pleistoceno, as causas potenciais de extinção incluem caça humana e mudanças climáticas associadas ao intervalo frio de Dryas recente.[72]
Veja também
Referências
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap Zurita, Alfredo Eduardo; Gillette, David D.; Cuadrelli, Francisco; Carlini, Alfredo Armando (1 de junho de 2018). «A tale of two clades: Comparative study of Glyptodon Owen and Glyptotherium Osborn (Xenarthra, Cingulata, Glyptodontidae)». Geobios. 51 (3): 247–258. Bibcode:2018Geobi..51..247Z. ISSN 0016-6995. doi:10.1016/j.geobios.2018.04.004. hdl:11336/83593

- ↑ a b c d e Gillette & Ray 1981, p. 3.
- ↑ a b Cuatáparo, J. N., & Ramírez, S. (1875). Descripción de un mamífero fósil de especie desconocida perteneciente al género" Glyptodon": encontrado entre las capas post-terciarias de Tequisquiac, en el Distrito de Zumpango. F. Diaz de Leon.
- ↑ Gillette & Ray 1981, p. 16.
- ↑ Felix, J. P. (1899). Beiträge zur geologie und paläontologie der republik Mexico (Vol. 1). A. Felix.
- ↑ Maldonado, M. (1948). Los vertebrados fósiles del Cuaternario en México. Revista de la Sociedad Mexicana de Historia Natural, 9 (1-2): 1-35
- ↑ a b c d e Osborn, Henry Fairfield (1903). Glyptotherium Texanum: A New Glyptodont from the Lower Pleistocene of Texas (em inglês). [S.l.]: order of the Trustees, American Museum of Natural History
- ↑ Cope, E. D. (1889). the edentata of North America. The American Naturalist, 23(272), 657-664.
- ↑ Leidy, J. (1889). Description of vertebrate remains from Peace Creek, Florida. Transactions of the Wagner Free Institute of Science of Philadelphia, 2, 19-31.
- ↑ Leidy, D. (1889). Fossil vertebrates from Florida. Proceedings of the Academy of Natural Sciences of Philadelphia, 96-97.
- ↑ a b Gillette & Ray 1981, p. 14.
- ↑ a b c d Brown, Barnum (1912). «Brachyostracon, a new genus of glyptodonts from Mexico.». Bulletin of the AMNH (em inglês). 31 (17): 167–190. hdl:2246/1402
- ↑ a b c d e f g h i Gillette, D. D.; Ray, C. E. (1981), Glyptodonts of North America (PDF)
- ↑ Hay, O. P. (1923). The Pleistocene of North America and Its Vertebrated Animals from the States East of the Mississippi River and from the Canadian Provinces East of Longitude 95> O (No. 22). Carnegie institution of Washington.
- ↑ Gillette & Ray 1981, p. 6 & 14.
- ↑ a b c Simpson, G. G., & Holmes, W. W. (1929). Pleistocene mammalian fauna of the Seminole Field, Pinellas County, Florida. Bulletin of the AMNH; v. 56, article 8.
- ↑ a b c d Meade, G. E. (1953). An early Pleistocene vertebrate fauna from Frederick, Oklahoma. The Journal of Geology, 61(5), 452-460.
- ↑ Gould, C. N. (1928). The fossil Glyptodon in the Frederick gravel beds. In Proceedings of the Oklahoma Academy of Science (pp. 148-150).
- ↑ Spier, Leslie (10 de fevereiro de 1928). «Concerning Man's Antiquity at Frederick, Oklahoma». Science (em inglês). 67 (1728): 160–161. Bibcode:1928Sci....67..160S. ISSN 0036-8075. PMID 17752885. doi:10.1126/science.67.1728.160
- ↑ a b c d e f g h i j k l m Gillette, David D.; Carranza-Castañeda, Óscar; White, Richard S.; Morgan, Gary S.; Thrasher, Larry C.; McCord, Robert; McCullough, Gavin (1 de junho de 2016). «Ontogeny and Sexual Dimorphism of Glyptotherium texanum (Xenarthra, Cingulata) from the Pliocene and Pleistocene (Blancan and Irvingtonian NALMA) of Arizona, New Mexico, and Mexico». Journal of Mammalian Evolution (em inglês). 23 (2): 133–154. ISSN 1573-7055. doi:10.1007/s10914-015-9309-6
- ↑ a b c d e f g h i j k Oliveira, É. V., Porpino, K. D., & Baretto, A. (2010). On the presence of Glyptotherium in the Late Pleistocene of Northeastern Brazil, and the status of "Glyptodon" and "Chlamydotherium". Paleobiogeographic implications.[ligação inativa] Neues Jahrbuch fur Geologie und Palaontologie-Abhandlungen, 258(3), 353.
- ↑ a b c d e Lessa, Carlos Micael Bonfim; Gomes, Verônica Santos; Cherkinsky, Alexander; Dantas, Mário André Trindade (2021). «Isotopic paleoecology (δ13C, δ18O) of two megamammals assemblages from the late pleistocene of Brazilian intertropical region». Journal of South American Earth Sciences (em inglês). 112. 103576 páginas. Bibcode:2021JSAES.11203576L. ISSN 0895-9811. doi:10.1016/j.jsames.2021.103576
- ↑ a b c Porpino, Kleberson de O.; Fernicola, Juan C.; Bergqvist, Lílian P. (18 de maio de 2010). «Revisiting the intertropical Brazilian species Hoplophorus euphractus (Cingulata, Glyptodontoidea) and the phylogenetic affinities of Hoplophorus». Journal of Vertebrate Paleontology. 30 (3): 911–927. Bibcode:2010JVPal..30..911P. ISSN 0272-4634. doi:10.1080/02724631003765735. hdl:11336/69074
- ↑ Lund, P. W. (1845): Conspectum-dasypodum. – Det Kongelige Dans- ke Videnskabernes Selskbas Naturvidenskabelige og Mathematiske Afhandlinger, 11: lxxxii-lxxxvi.
- ↑ a b c d e f g h i j k l m Carlini, Alfredo A.; Carrillo-Briceño, Jorge D.; Jaimes, Arturo; Aguilera, Orangel; Zurita, Alfredo E.; Iriarte, José; Sánchez-Villagra, Marcelo R. (16 de junho de 2022). «Damaged glyptodontid skulls from Late Pleistocene sites of northwestern Venezuela: evidence of hunting by humans?». Swiss Journal of Palaeontology. 141 (1). 11 páginas. Bibcode:2022SwJP..141...11C. ISSN 1664-2384. doi:10.1186/s13358-022-00253-3
- ↑ a b c Mitchell, K.J.; Scanferla, A.; Soibelzon, E.; Bonini, R.; Ochoa, J.; Cooper, A. (2016). «Ancient DNA from the extinct South American giant glyptodont Doedicurus sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos». Molecular Ecology. 25 (14): 3499–3508. Bibcode:2016MolEc..25.3499M. PMID 27158910. doi:10.1111/mec.13695. hdl:11336/48521
- ↑ a b Delsuc, F.; Gibb, G.C.; Kuch, M.; Billet, G.; Hautier, L.; Southon, J.; Rouillard, J.-M.; Fernicola, J.C.; Vizcaíno, S.F.; MacPhee, R.D.E.; Poinar, H.N. (2016). «The phylogenetic affinities of the extinct glyptodonts». Current Biology. 26 (4): R155–R156. Bibcode:2016CBio...26.R155D. PMID 26906483. doi:10.1016/j.cub.2016.01.039. hdl:11336/49579
- ↑ Delsuc, F.; Gibb, G.C.; Kuch, M.; Billet, G.; Hautier, L.; Southon, J.; Rouillard, J.-M.; Fernicola, J. C.; Vizcaíno, S. F.; MacPhee, R. D.E.; Poinar, H. N. (22 de fevereiro de 2016). «The phylogenetic affinities of the extinct glyptodonts» (PDF). Current Biology. 26 (4): R155–R156. Bibcode:2016CBio...26.R155D. PMID 26906483. doi:10.1016/j.cub.2016.01.039
- ↑ Delsuc, Frédéric; Gibb, Gillian C.; Kuch, Melanie; Billet, Guillaume; Hautier, Lionel; Southon, John; Rouillard, Jean-Marie; Fernicola, Juan Carlos; Vizcaíno, Sergio F. (fevereiro de 2016). «The phylogenetic affinities of the extinct glyptodonts». Current Biology (em inglês) (4): R155–R156. doi:10.1016/j.cub.2016.01.039. Consultado em 22 de agosto de 2025
- ↑ Cuadrelli, Francisco; Zurita, Alfredo E.; Toriño, Pablo; Miño-Boilini, Ángel R.; Rodríguez-Bualó, Santiago; Perea, Daniel; Acuña Suárez, Gabriel E. (3 de setembro de 2018). «Late Pleistocene Glyptodontinae (Mammalia, Xenarthra, Glyptodontidae) from southern South America: a comprehensive review». Journal of Vertebrate Paleontology. 38 (5): e1525390. Bibcode:2018JVPal..38E5390C. ISSN 0272-4634. doi:10.1080/02724634.2018.1525390
- ↑ a b Zurita, Alfredo E.; González Ruiz, Laureano R.; Gómez-Cruz, Arley J.; Arenas-Mosquera, Jose E. (1 de maio de 2013). «The most complete known Neogene Glyptodontidae (Mammalia, Xenarthra, Cingulata) from northern South America: taxonomic, paleobiogeographic, and phylogenetic implications». Journal of Vertebrate Paleontology. 33 (3): 696–708. Bibcode:2013JVPal..33..696Z. ISSN 0272-4634. doi:10.1080/02724634.2013.726677. hdl:11336/2971
- ↑ a b Zurita, Alfredo E.; Miño-Boilini, Ángel R.; Francia, Analía; Arenas-Mosquera, José E. (31 de dezembro de 2012). «The Pleistocene Glyptodontidae Gray, 1869 (Xenarthra: Cingulata) of Colombia and some considerations about the South American Glyptodontinae» (PDF). Revista Brasileira de Paleontologia. 15 (3): 273–280. doi:10.4072/rbp.2012.3.04
- ↑ Zurita, Alfredo E.; González Ruiz, Laureano R.; Gómez-Cruz, Arley J.; Arenas-Mosquera, Jose E. (maio de 2013). «The most complete known Neogene Glyptodontidae (Mammalia, Xenarthra, Cingulata) from northern South America: taxonomic, paleobiogeographic, and phylogenetic implications». Journal of Vertebrate Paleontology (em inglês) (3): 696–708. ISSN 0272-4634. doi:10.1080/02724634.2013.726677. Consultado em 22 de agosto de 2025
- ↑ Zurita, Alfredo E.; Miño-Boilini, Ángel R.; Francia, Analía; Arenas-Mosquera, José E. (31 de dezembro de 2012). «The Pleistocene Glyptodontidae Gray, 1869 (Xenarthra: Cingulata) of Colombia and some considerations about the South American Glyptodontinae» (PDF). REVISTA BRASILEIRA DE PALEONTOLOGIA (03): 273–280. doi:10.4072/rbp.2012.3.04. Consultado em 22 de agosto de 2025
- ↑ Cuadrelli, Francisco; Zurita, Alfredo E.; Toriño, Pablo; Miño-Boilini, Ángel R.; Perea, Daniel; Luna, Carlos A.; Gillette, David D.; Medina, Omar (16 de setembro de 2020). «A new species of glyptodontine (Mammalia, Xenarthra, Glyptodontidae) from the Quaternary of the Eastern Cordillera, Bolivia: phylogeny and palaeobiogeography». Journal of Systematic Palaeontology. 18 (18): 1543–1566. Bibcode:2020JSPal..18.1543C. ISSN 1477-2019. doi:10.1080/14772019.2020.1784300
- ↑ Gillette & Ray 1981, p. 2.
- ↑ Gillette & Ray 1981, p. 1.
- ↑ a b c du Plessis, Anton; Broeckhoven, Chris; Yadroitsev, Igor; Yadroitsava, Ina; le Roux, Stephan Gerhard (1 de junho de 2018). «Analyzing nature's protective design: The glyptodont body armor». Journal of the Mechanical Behavior of Biomedical Materials (em inglês). 82: 218–223. ISSN 1751-6161. PMID 29621689. doi:10.1016/j.jmbbm.2018.03.037
- ↑ Gillette & Ray 1981, p. 200.
- ↑ Gillette & Ray 1981, p. 58.
- ↑ Gillette & Ray 1981, p. 39.
- ↑ Zurita, A. E., Miño-Boilini, Á. R., Soibelzon, E., Carlini, A. A., & Paredes Rios, F. (2009). The diversity of Glyptodontidae (Xenarthra, Cingulata) in the Tarjia Valley (Bolivia): Systematic, biostratigraphic and paleobiogeographic aspects of a particular assemblage.(With 3 figures and 1 table). Neues Jahrbuch fur Geologie und Palaontologie-Abhandlungen, 251(2), 225.
- ↑ Zurita, A.E.; Scarano, A.C.; Carlini, A.A.; Scillato-Yané, G.J.; Soibelzon, E. (4 de abril de 2011). «Neosclerocalyptus spp. (Cingulata: Glyptodontidae: Hoplophorini): cranial morphology and palaeoenvironments along the changing Quaternary». Journal of Natural History. 45 (15–16): 893–914. Bibcode:2011JNatH..45..893Z. ISSN 0022-2933. doi:10.1080/00222933.2010.536917
- ↑ Fernicola, Juan Carlos; Toledo, Néstor; Bargo, M. Susana; Vizcaíno, Sergio F. (22 de setembro de 2012). «A neomorphic ossification of the nasal cartilages and the structure of paranasal sinus system of the glyptodont Neosclerocalyptus Paula Couto 1957 (Mammalia, Xenarthra)». Palaeontologia Electronica (em inglês). 15 (3): 1–22. ISSN 1094-8074. doi:10.26879/333. hdl:11336/68093
- ↑ a b c Gillette & Ray 1981, p. 202.
- ↑ a b c Zurita, Alfredo Eduardo; Soibelzon, Leopoldo Hector; Soibelzon, Esteban; Gasparini, Germán Mariano; Cenizo, Marcos Martín; Arzani, Héctor (1 de janeiro de 2010). «Accessory protection structures in Glyptodon Owen (Xenarthra, Cingulata, Glyptodontidae)». Annales de Paléontologie (em inglês). 96 (1): 1–11. Bibcode:2010AnPal..96....1Z. ISSN 0753-3969. doi:10.1016/j.annpal.2010.01.001
- ↑ Thrasher, Larry (julho de 2022). «Fossils and Age Relationships of the Late Pliocene and Early Pleistocene (Blancan) 111 Ranch Beds and Bear Springs Wash Beds, Graham County, Arizona». Late Cenozoic Vertebrate Paleontology: Tribute to Arthur H. Harris. New Mexico Museum of Natural History and Science Bulletin. 88 – via ResearchGate
- ↑ Gillette & Ray 1981, p. 169, 172, & 173.
- ↑ a b c d e Amson, Eli; Nyakatura, John A. (1 de dezembro de 2018). «The Postcranial Musculoskeletal System of Xenarthrans: Insights from over Two Centuries of Research and Future Directions». Journal of Mammalian Evolution (em inglês). 25 (4): 459–484. ISSN 1573-7055. doi:10.1007/s10914-017-9408-7
- ↑ a b c Owen, Richard (1841). «VI.— Description of a Tooth and Part of the Skeleton of the Glyptodon clavipes, a large Quadruped of the Edentate Order, to which belongs the Tesselated Bony Armour described and figured by Mr. Clift in the former Volume of the Transactions of the Geological Society; with a consideration of the question whether the Megatherium possessed an analogous Dermal Armour.». Transactions of the Geological Society of London (em inglês). 6 (1): 81–106. ISSN 2042-5295. doi:10.1144/transgslb.6.1.81
- ↑ Sénéchal, D. L. (1865). Notice sur l'armure ou le dermato-squelette et le système dentaire du Glyptodon clavipes, et particularités biologiques de cet animal, déduites d'après l'étude de ses restes fossiles. Balitout, Questroy et Cie.
- ↑ Fariña, R. A., & Vizcaíno, S. F. (1997). Allometry of the bones of living and extinct armadillos (Xenarthra, Dasypoda). Zeitschrift fur Saugetierkunde, 62, 65-70.
- ↑ Fariña, Richard A.; Vizcaíno, Sergio F.; Blanco, R. Ernesto (21 de abril de 1997). «Scaling of the Indicator of Athletic Capability in Fossil and Extant Land Tetrapods». Journal of Theoretical Biology (em inglês). 185 (4): 441–446. Bibcode:1997JThBi.185..441F. ISSN 0022-5193. doi:10.1006/jtbi.1996.0323
- ↑ Shockey, B. J. (2001). Specialized knee joints in some extinct, endemic, South American herbivores. Acta Palaeontologica Polonica, 46(2).
- ↑ a b Russell, Dale A.; Rich, Fredrick J.; Schneider, Vincent; Lynch-Stieglitz, Jean (maio de 2009). «A warm thermal enclave in the Late Pleistocene of the South-eastern United States». Biological Reviews (em inglês). 84 (2): 173–202. ISSN 1464-7931. PMID 19391200. doi:10.1111/j.1469-185X.2008.00069.x
- ↑ a b c d e f Pérez-Crespo, Victor Adrián; Arroyo-Cabrales, Joaquín; Alva-Valdivia, Luis M.; Morales-Puente, Pedro; Cienfuegos-Alvarado, Edith (2012). «Diet and habitat definitions for Mexican glyptodonts from Cedral (San Luis Potosí, México) based on stable isotope analysis». Geological Magazine (em inglês). 149 (1): 153–157. ISSN 1469-5081. doi:10.1017/S0016756811000951
- ↑ Fariña, Richard A.; Vizcaíno, Sergio F. (2024). «Giants beasts updated: A review of new knowledge about the South American megafauna». Journal of Quaternary Science (em inglês). 39 (8): 1139–1153. ISSN 1099-1417. doi:10.1002/jqs.3663
- ↑ Keeley, J. E., & Rundel, P. W. (2003). Evolution of CAM and C4 carbon-concentrating mechanisms. International journal of plant sciences, 164(S3), S55-S77.
- ↑ a b Omena, Érica Cavalcante; Silva, Jorge Luiz Lopes da; Sial, Alcides Nóbrega; Cherkinsky, Alexander; Dantas, Mário André Trindade (3 de outubro de 2021). «Late Pleistocene meso-megaherbivores from Brazilian Intertropical Region: isotopic diet (δ13C), niche differentiation, guilds and paleoenvironmental reconstruction (δ13C, δ18O)». Historical Biology. 33 (10): 2299–2304. Bibcode:2021HBio...33.2299O. ISSN 0891-2963. doi:10.1080/08912963.2020.1789977
- ↑ Dantas, M. A. T., Cherkinsky, A., Lessa, C. M. B., Santos, L. V., Cozzuol, M. A., Omena, É. C., ... & Bocherens, H. (2020). Isotopic paleoecology (δ13C, δ18O) of a late Pleistocene vertebrate community from the Brazilian Intertropical Region. Revista Brasileira de Paleontologia, 23(2), 138-152.
- ↑ Phillip E. Jardine, Christine M. Janis, Sarda Sahney, Michael J. Benton. "Grit not grass: Concordant patterns of early origin of hypsodonty in Great Plains ungulates and Glires." Palaeogeography, Palaeoclimatology, Palaeoecology. December 2012:365–366, 1–10
- ↑ De Iuliis, Gerardo; Bargo, María S.; Vizcaíno, Sergio F. (19 de janeiro de 2001). «Variation in skull morphology and mastication in the fossil giant armadillos Pampatherium spp. and allied genera (Mammalia: Xenarthra: Pampatheriidae), with comments on their systematics and distribution». Journal of Vertebrate Paleontology. 20 (4): 743–754. ISSN 0272-4634. doi:10.1671/0272-4634(2000)020[0743:VISMAM]2.0.CO;2
- ↑ Vizcaíno, Sergio F.; Cassini, Guillermo H.; Fernicola, Juan C.; Bargo, M. Susana (2011). "Evaluating Habitats and Feeding Habits Through Ecomorphological Features in Glyptodonts (Mammalia, Xenarthra)". Ameghiniana: 305–319. doi:10.5710/AMGH.v48i3(364).. Retrieved 2015-10-29.
- ↑ Vizcaíno, S. F., Fariña, R. A., & Mazzetta, G. V. (1999). Ulnar dimensions and fossoriality in armadillos. Acta Theriologica, 44.
- ↑ Carter, T. S., & Encarnaçao, C. D. (1983). Characteristics and use of burrows by four species of armadillos in Brazil. Journal of Mammalogy, 64(1), 103-108.
- ↑ Vizcaíno, Sergio F.; Fariña, Richard A. (29 de março de 2007). «Diet and locomotion of the armadillo Peltephilus: a new view». Lethaia (em inglês). 30 (1): 79–86. doi:10.1111/j.1502-3931.1997.tb00446.x
- ↑ Saarinen, Juha; Karme, Aleksis (15 de junho de 2017). «Tooth wear and diets of extant and fossil xenarthrans (Mammalia, Xenarthra) – Applying a new mesowear approach». Palaeogeography, Palaeoclimatology, Palaeoecology (em inglês). 476: 42–54. Bibcode:2017PPP...476...42S. ISSN 0031-0182. doi:10.1016/j.palaeo.2017.03.027
- ↑ Vizcaíno; Blanco, R. Ernesto; Bender, J. Benjamín; Milne, Nick (2011). «Proportions and function of the limbs of glyptodonts: Glyptodont limbs». Lethaia (em inglês). 44 (1): 93–101. Bibcode:2011Letha..44...93V. doi:10.1111/j.1502-3931.2010.00228.x
- ↑ Chen, Irene H.; Kiang, James H.; Correa, Victor; Lopez, Maria I.; Chen, Po-Yu; McKittrick, Joanna; Meyers, Marc A. (1 de julho de 2011). «Armadillo armor: Mechanical testing and micro-structural evaluation». Journal of the Mechanical Behavior of Biomedical Materials. Special Issue on Natural Materials / Papers from the Third International Conference on the Mechanics of Biomaterials and Tissues (em inglês). 4 (5): 713–722. ISSN 1751-6161. PMID 21565719. doi:10.1016/j.jmbbm.2010.12.013
- ↑ Barbosa, Fernando Henrique de Souza; Porpino, Kleberson de Oliveira; Fragoso, Ana Bernadete Lima; Oliveira, Edison Vicente (13 de fevereiro de 2014). «Arthritis in a Glyptodont (Mammalia, Xenarthra, Cingulata)». PLOS ONE (em inglês). 9 (2): e88646. Bibcode:2014PLoSO...988646B. ISSN 1932-6203. PMC 3923812. PMID 24551126. doi:10.1371/journal.pone.0088646
- ↑ Lima, Fábio Cunha Guimarães de; Porpino, Kleberson de Oliveira (18 de outubro de 2018). «Ectoparasitism and infections in the exoskeletons of large fossil cingulates». PLOS ONE (em inglês). 13 (10): e0205656. Bibcode:2018PLoSO..1305656D. ISSN 1932-6203. PMC 6193641. PMID 30335796. doi:10.1371/journal.pone.0205656
- ↑ a b c d e Faith, J. Tyler; Surovell, Todd A. (8 de dezembro de 2009). «Synchronous extinction of North America's Pleistocene mammals». Proceedings of the National Academy of Sciences (em inglês). 106 (49): 20641–20645. Bibcode:2009PNAS..10620641F. ISSN 0027-8424. PMC 2791611. PMID 19934040. doi:10.1073/pnas.0908153106
- ↑ Repenning, Charles A. (1962). "The Giant Ground Squirrel Paenemarmota". Journal of Paleontology. 36 (93): 540–556. JSTOR 1301086.
- ↑ Dalquest, Walter W. (1968). «The Bone-Eating Dog, Borophagus diversidens Cope». Quarterly Journal of the Florida Academy of Sciences. 31 (2): 115–129. ISSN 0015-3850
- ↑ Kurtén, Björn; and Werdelin, Lars (31 de março de 1988). «A review of the genus Chasmaporthetes Hay, 1921 (Carnivora, Hyaenidae)». Journal of Vertebrate Paleontology. 8 (1): 46–66. ISSN 0272-4634. doi:10.1080/02724634.1988.10011683
- ↑ Berta, Annalisa (1 de dezembro de 1981). «The Plio-Pleistocene hyaena Chasmaporthetes ossifragus from Florida». Journal of Vertebrate Paleontology. 1 (3-4): 341–356. ISSN 0272-4634. doi:10.1080/02724634.1981.10011905
- ↑ Martin, Larry; Schultz, C. B.; Schultz, M. R. (1988). «Saber-Toothed Cats from the Plio-Pleistocene of Nebraska». Transactions of the Nebraska Academy of Sciences and Affiliated Societies. 186: 153-163
- ↑ Dalquest, Walter Woelber; Dalquest, Walter Woelber (1975). Vertebrate fossils from the Blanco local fauna of Texas. [Lubbock]: Museum, Texas Tech University. doi:10.5962/bhl.title.142927
- ↑ Feduccia, J. Alan; Ford, Norman L. (1 de outubro de 1970). «Some birds of prey from the Upper Pliocene of Kansas». The Auk. 87 (4): 795–797. ISSN 1938-4254. JSTOR 4083714. doi:10.2307/4083714
- ↑ Marshall, William H. (1 de janeiro de 1937). «Double-crested Cormorant Nesting on the Bear River Refuge in Utah». The Condor. 39 (1). 36 páginas. ISSN 1938-5129. JSTOR 1363487. doi:10.2307/1363487
- ↑ a b c Dávila, S. Lorena; Stinnesbeck, Sarah R.; Gonzalez, Silvia; Lindauer, Susanne; Escamilla, Juan; Stinnesbeck, Wolfgang (1 de setembro de 2019). «Guatemala's Late Pleistocene (Rancholabrean) fauna: Revision and interpretation». Quaternary Science Reviews (em inglês). 219: 277–296. Bibcode:2019QSRv..219..277D. ISSN 0277-3791. doi:10.1016/j.quascirev.2019.07.011
- ↑ Stinnesbeck, Sarah (2020). «Mexican fossil ground sloths - A case study for Late Pleistocene megafaunal turnover in the Mexican Corridor». Hochschulschrift. 14. 29 páginas
- ↑ Keeley, Jon; Rundel, Philip (2003). «Evolution of CAM and C4 Carbon‐Concentrating Mechanisms». International Journal of Plant Sciences. 164 (S3): S55–S77. ISSN 1058-5893. doi:10.1086/374192
- ↑ a b c Cartelle, Castor; Hartwig, W. C. (1996). "A new extinct primate among the Pleistocene megafauna of Bahia, Brazil". Proceedings of the National Academy of Sciences. 93 (13): 6405–6409.
- ↑ Eisenberg, John F.; Redford, Kent H. (1989). Mammals of the Neotropics, Volume 3: Ecuador, Bolivia, Brazil. University of Chicago Press. p. 247. ISBN 978-0-226-19542-1.
- ↑ Halenar, Lauren B. (2011). «Reconstructing the Locomotor Repertoire of Protopithecus brasiliensis. I. Body Size». The Anatomical Record (em inglês). 294 (12): 2024–2047. ISSN 1932-8494. doi:10.1002/ar.21501
- ↑ a b David Webb, S. (23 de agosto de 2006). «The Great American Biotic Interchange: Patterns and Processes1». Annals of the Missouri Botanical Garden (em inglês). 93 (2): 245–257. ISSN 0026-6493. doi:10.3417/0026-6493(2006)93[245:TGABIP]2.0.CO;2
- ↑ Charrier, John J. Flynn, André R. Wyss and Reynaldo (2007). «South America's Missing Mammals». Scientific American (em inglês). 296 (5): 68–75. Bibcode:2007SciAm.296e..68F. PMID 17500416. doi:10.1038/scientificamerican0507-68. Consultado em 9 de maio de 2023
- ↑ Bloch, Jonathan I.; Woodruff, Emily D.; Wood, Aaron R.; Rincon, Aldo F.; Harrington, Arianna R.; Morgan, Gary S.; Foster, David A.; Montes, Camilo; Jaramillo, Carlos A.; Jud, Nathan A.; Jones, Douglas S.; MacFadden, Bruce J. (2016). «First North American fossil monkey and early Miocene tropical biotic interchange». Nature (em inglês). 533 (7602): 243–246. ISSN 1476-4687. doi:10.1038/nature17415
- ↑ Croft, D. A. (2016). Horned armadillos and rafting monkeys: the fascinating fossil mammals of South America. Indiana University Press.
- ↑ Morgan, G. S. (2005). The great American biotic interchange in Florida. Bulletin of the Florida Museum of Natural History, 45(4), 271-311.
- ↑ McDonald, H. G. (2005). Paleoecology of extinct xenarthrans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History, 45(4), 319-340.
- ↑ a b Scillato-Yané, G. J.; Carlini, A. A.; Tonni, E. P.; Noriega, J. I. (1 de outubro de 2005). «Paleobiogeography of the late Pleistocene pampatheres of South America». Journal of South American Earth Sciences. Quaternary Paleontology and biostratigraphy of southern South Africa (em inglês). 20 (1): 131–138. Bibcode:2005JSAES..20..131S. ISSN 0895-9811. doi:10.1016/j.jsames.2005.06.012. hdl:11336/80762
- ↑ a b c Carlini, Alfredo A.; Zurita, Alfredo E.; Aguilera, Orangel A. (2008). «North American Glyptodontines (Xenarthra, Mammalia) in the Upper Pleistocene of northern South America». Paläontologische Zeitschrift (em inglês). 82 (2): 125–138. Bibcode:2008PalZ...82..125C. ISSN 0031-0220. doi:10.1007/BF02988404. eISSN 1867-6812. hdl:11336/36844
- ↑ a b Cione, A. L., Gasparini, G. M., Soibelzon, E., Soibelzon, L. H., & Tonni, E. P. (2015). The great American biotic interchange: a South American perspective (p. 97). Dordrecht: Springer.
- ↑ Woodburne, Michael O. (1 de dezembro de 2010). «The Great American Biotic Interchange: Dispersals, Tectonics, Climate, Sea Level and Holding Pens». Journal of Mammalian Evolution (em inglês). 17 (4): 245–264. ISSN 1573-7055. PMC 2987556. PMID 21125025. doi:10.1007/s10914-010-9144-8
- ↑ Lundelius, Ernest L.; Bryant, Vaughn M.; Mandel, Rolfe; Thies, Kenneth J.; Thoms, Alston (1 de janeiro de 2013). «The first occurrence of a toxodont (Mammalia, Notoungulata) in the United States». Journal of Vertebrate Paleontology. 33 (1): 229–232. Bibcode:2013JVPal..33..229L. ISSN 0272-4634. doi:10.1080/02724634.2012.711405. hdl:1808/13587
- ↑ a b Zurita, Alfredo E.; Carlini, Alfredo A.; Gillette, David; Sánchez, Rodolfo (1 de março de 2011). «Late Pliocene Glyptodontinae (Xenarthra, Cingulata, Glyptodontidae) of South and North America: Morphology and paleobiogeographical implications in the GABI». Journal of South American Earth Sciences. 31 (2): 178–185. Bibcode:2011JSAES..31..178Z. ISSN 0895-9811. doi:10.1016/j.jsames.2011.02.001. hdl:11336/41714
- ↑ Pujos, F., & Salas, R. (2004). A systematic reassessment and paleogeographic review of fossil Xenarthra from Peru. Bulletin de l'Institut français d'études andines, (33 (2)), 331-377.
- ↑ Rabassa, Jorge; Coronato, Andrea M.; Salemme, Mónica (1 de outubro de 2005). «Chronology of the Late Cenozoic Patagonian glaciations and their correlation with biostratigraphic units of the Pampean region (Argentina)». Journal of South American Earth Sciences. Quaternary Paleontology and biostratigraphy of southern South Africa (em inglês). 20 (1): 81–103. Bibcode:2005JSAES..20...81R. ISSN 0895-9811. doi:10.1016/j.jsames.2005.07.004. hdl:11336/150967
- ↑ Magoulick, Katherine M.; Saupe, Erin E.; Farnsworth, Alexander; Valdes, Paul J.; Marshall, Charles R. (2025). «Evaluating migration hypotheses for the extinct Glyptotherium using ecological niche modeling». Ecography (em inglês). n/a (n/a): e07499. ISSN 1600-0587. doi:10.1111/ecog.07499
- ↑ Antón, Mauricio (2013). Sabertooth. Bloomington, Indiana: University of Indiana Press. pp. 203–204. ISBN 978-0-253-01042-1
- ↑ Gillette, D. D. (primavera de 2010). «Glyptodonts in Arizona». Arizona Geological Survey. Arizona Geology. Consultado em 17 de agosto de 2018
- ↑ Brandes, G. (1900) : Ueber eine Ursache des Aussterbens Diluvialer Säugethiere. Corrblatt d. Deutsch . Ges. f. Anthropol. Jahrg. 31. Munichen 1901.
- ↑ Bohlin, B. (1940). 8. Food habit of the machairodonts, with special regard to Smilodon. Bull. Geol. Inst. Upsala.28:156-174
- ↑ Araújo-Júnior, Hermínio Ismael de; Barbosa, Fernando Henrique de Souza; Silva, Lucas Henrique Medeiros da (15 de fevereiro de 2017). «Overlapping paleoichnology, paleoecology and taphonomy: Analysis of tooth traces in a Late Pleistocene-early Holocene megafaunal assemblage of Brazil and description of a new ichnotaxon in hard substrate». Palaeogeography, Palaeoclimatology, Palaeoecology (em inglês). 468: 122–128. Bibcode:2017PPP...468..122A. ISSN 0031-0182. doi:10.1016/j.palaeo.2016.12.007
- ↑ Crook, Wilson W.; Harris, R. K. (1958). «A Pleistocene Campsite near Lewisville, Texas». American Antiquity (em inglês). 23 (3): 233–246. ISSN 0002-7316. JSTOR 276304. doi:10.2307/276304
- ↑ Waguespack, Nicole M.; Surovell, Todd A. (2003). «Clovis Hunting Strategies, or How to Make out on Plentiful Resources». American Antiquity (em inglês). 68 (2): 333–352. ISSN 0002-7316. JSTOR 3557083. doi:10.2307/3557083
- ↑ Prates, Luciano; Perez, S. Ivan (12 de abril de 2021). «Late Pleistocene South American megafaunal extinctions associated with rise of Fishtail points and human population». Nature Communications (em inglês). 12 (1). 2175 páginas. Bibcode:2021NatCo..12.2175P. ISSN 2041-1723. PMC 8041891. PMID 33846353. doi:10.1038/s41467-021-22506-4
- ↑ Vogt, C. (1881). Squelette humain associe aux glyptodontidae. Bulletin de la Société d'Antropologie de Paris, 3(4), 693–699
- ↑ Quiñones, Sofía I.; De los Reyes, Martin; Zurita, Alfredo E.; Cuadrelli, Francisco; Miño-Boilini, Ángel R.; Poiré, Daniel G. (1 de novembro de 2020). «Neosclerocalyptus Paula Couto (Xenarthra, Glyptodontidae) in the late Pliocene-earliest Pleistocene of the Pampean region (Argentina): Its contribution to the understanding of evolutionary history of Pleistocene glyptodonts». Journal of South American Earth Sciences (em inglês). 103. 102701 páginas. Bibcode:2020JSAES.10302701Q. ISSN 0895-9811. doi:10.1016/j.jsames.2020.102701
- ↑ Politis, Gustavo G.; Messineo, Pablo G.; Stafford, Thomas W.; Lindsey, Emily L. (2019). «Campo Laborde: A Late Pleistocene giant ground sloth kill and butchering site in the Pampas». Science Advances (em inglês). 5 (3): eaau4546. Bibcode:2019SciA....5.4546P. ISSN 2375-2548. PMC 6402857. PMID 30854426. doi:10.1126/sciadv.aau4546
- ↑ a b Albright, L.; Sanders, Albert; Weems, Robert; Cicimurri, David; Knight, James (31 de outubro de 2019). «Cenozoic vertebrate biostratigraphy of South Carolina, U.S.A., and additions to the fauna». Bulletin of the Florida Museum of Natural History (em inglês). 57 (2): 77–236. ISSN 2373-9991. doi:10.58782/flmnh.qqgg4577
- ↑ Sanders, Albert E. (1 de janeiro de 2002). Additions to the Pleistocene Mammal Faunas of South Carolina, North Carolina, and Georgia: Transactions, American Philosophical Society (vol. 92, part 5) (em inglês). [S.l.]: The American Philosophical Society Press. ISBN 979-8-89398-336-4. doi:10.70249/9798893983364
- ↑ Weems, Robert E.; Bachman, Jon M.; Rayburn, William T.; McCloud Jr., James R. (2021). «A Vertebrate Ichnofauna from the Late Pleistocene (Sangamon Interglacial) Tabb Formation at Stratford Hall Plantation, Westmoreland County, Virginia USA)». New Mexico Museum of Natural History and Science. Bulletin 82: 505-512 – via ResearchDirect
- ↑ a b Valerio, A. L., & Laurito, C. A. (2011). El registro fósil de Glyptotherium floridanum (Xenarthra, Cingulata, Glyptodontidae) en el Cuaternario de Costa Rica, América Central. Revista Geológica de América Central, (45), 141-145.
- ↑ Jackson, D. R., & Fernandez, E. (2005). A small Pleistocene mammalian megafauna from southern Honduras. Bulletin of the Florida Museum of Natural History, 45(4), 261-269.
- ↑ Cisneros, J. C. (2005). New pleistocene vertebrate fauna from El Salvador. Revista Brasileira de Paleontologia, 8(3), 239-255.
- ↑ Lucas, Spencer G. (2014). «Late pleistocene mammals from El Hatillo, Panama». Revista Geológica de América Central (em inglês) (50): 139–151. ISSN 0256-7024
- ↑ Valerio, Ana L.; Laurito, César A. (2011). «El registro fósil de Glyptotherium Floridanum (Xenarthra, Cingulata, Glyptodontidae) en el Cuaternario de Costa Rica, América Central». Revista Geológica de América Central (em espanhol) (45): 141–145. ISSN 0256-7024
- ↑ Cuadrelli, Francisco; Escamilla, Juan; Zurita, Alfredo; Gillette, David D.; Dávila, Lorena S. (22 de agosto de 2023). «Glyptotherium cylindricum (Cingulata, Glyptodontidae) from the Late Pleistocene of Guatemala: the most complete record of Glyptodontinae from Central America». Alcheringa: An Australasian Journal of Palaeontology (em inglês). 47 (3): 336–347. Bibcode:2023Alch...47..336C. ISSN 0311-5518. doi:10.1080/03115518.2023.2242440
- ↑ a b Ramírez-Cruz, Gonzalo A.; Montellano-Ballesteros, Marisol (2014). «Two new glyptodont records (Mammalia: Cingulata) from the late Pleistocene of Tamaulipas and Tlaxcala, Mexico: Implications for the taxonomy of the genus Glyptotherium». The Southwestern Naturalist. 59 (4): 522–530. ISSN 0038-4909. doi:10.1894/JKF-45.1
- ↑ Haynes, Gary (8 de fevereiro de 2013). «Extinctions in North America's Late Glacial landscapes». Quaternary International. Peopling the last new worlds: the first colonisation of Sahul and the Americas. 285: 89–98. Bibcode:2013QuInt.285...89H. ISSN 1040-6182. doi:10.1016/j.quaint.2010.07.026
- ↑ Stuart, Anthony John (maio de 2015). «Late Quaternary megafaunal extinctions on the continents: a short review». Geological Journal (em inglês). 50 (3): 338–363. Bibcode:2015GeolJ..50..338S. ISSN 0072-1050. doi:10.1002/gj.2633
- ↑ Carlini, A.A.; Zurita, A.E. (2006). «Glyptotherium Osborn (Mammalia, Xenarthra, Cingulata) en el Pleistoceno Tardío de Venezuela». 9° Congreso Argentino de Paleontología y Bioestratigrafía, Córdoba. 98 páginas
- ↑ Faria, Fabio Henrique Cortes; Kinoshita, Angela; Carvalho, Ismar de Souza; Araújo-Júnior, Hermínio Ismael de; Pegorin, Priscila; Maria G Figueiredo, Ana; Baffa, Oswaldo (1 de agosto de 2020). «ESR dating of late Quaternary megafauna fossils from João Dourado, Bahia, Brazil». Journal of South American Earth Sciences. 101. 102586 páginas. Bibcode:2020JSAES.10102586F. ISSN 0895-9811. doi:10.1016/j.jsames.2020.102586