Evidências da evolução
| Parte de uma série sobre |
| Evolução |
|---|
 |
|
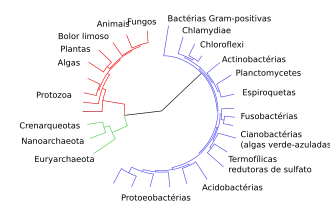
O termo evidências da evolução refere-se ao conjunto de dados empíricos, oriundos de múltiplas áreas das ciências biológicas e geológicas, que corroboram a hipótese de que as espécies se modificam ao longo do tempo por processos naturais e compartilham ancestrais comuns. Essas evidências não constituem provas isoladas, mas um corpo integrado de observações independentes que convergem para o mesmo modelo explicativo denominado evolução biológica. [1][2][3][4]
No contexto científico, uma evidência é um dado verificável que sustenta ou refuta uma hipótese. Assim, as evidências evolutivas são linhas de suporte testáveis, obtidas por métodos distintos (morfologia, genética, geologia, biogeografia, embriologia, observação experimental), que reforçam a robustez da teoria da evolução. A força dessas evidências deriva não apenas do volume acumulado, mas da congruência entre resultados independentes: registros fósseis[5], homologias anatômicas[6], sequências de DNA[7], distribuição geográfica e observações experimentais apontam para um conjunto de explicações coerentes e mutuamente consistentes.
Fósseis são importantes para estimar quando as várias linhagens se desenvolveram. Como a fossilização é de rara ocorrência, normalmente requerendo tecidos duros do corpo dos espécimes e da morte próxima a um local onde sedimentos estão sendo depositados, o registro fóssil somente fornece informações intermitentes sobre a evolução da vida. Evidências de organismos anteriores ao desenvolvimento de partes duras do corpo como conchas, ossos e dentes são especialmente raras, mas existem na forma de antigos microfósseis de alguns organismos de tecido mole.[5]
Comparações da sequência genética de organismos revelam que os organismos que são filogeneticamente mais próximos tem um grau maior de similaridades em sua sequência genética do que organismos que estão mais filogeneticamente distantes. Evidências adicionais da descendência em comum vem de “detritos” genéticos como os pseudogenes, regiões do DNA que são ortólogas a um gene em um organismo aparentado, mas não são mais ativos e aparentam passar por um constante processo de degeneração.[8] Já que processos metabólicos não deixam fósseis, pesquisas evolutivas sobre os básicos processos biológicos são também feitas na sua maior parte com a comparação de organismos existentes. Muitas linhagens divergem em um diferente estágio de desenvolvimento, então é teoricamente possível determinar quando certos processos metabólicos surgiram realizando uma comparação dos traços de um descendente de um ancestral em comum.[9]
Evidências da Paleontologia

Quando um organismo morre, ele normalmente decompõem-se rapidamente ou é consumidos por necrófagos, não deixando nenhuma evidência permanente de sua existência. Entretanto, ocasionalmente, alguns organismos são preservados. Os restos ou traços dos organismos de uma era geológica passada, envoltos em rocha por processos naturais são chamados de fósseis. A análise de fósseis oferece insights e novos conhecimentos que permitem o entendimento da história evolutiva da vida na Terra, já que fornecem evidência direta da evolução e informações detalhadas sobre as linhagens dos organismos.[10] Portanto, a paleontologia, que é o estudo da vida passada baseado no registro fóssil, compõe uma linha verificável de evidências.
Sendo o estudo da biologia dos organismos do passado geológico, por via do estudo dos fósseis, a paleobiologia, surge dentro da paleontologia portando duas disciplinas relevantes para a compreensão da evolução biológica das principais linhagens de plantas e animais: a paleozoologia e a paleobotânica.[11] É justamente no seio da paleobiologia que se insere a paleozoologia, o estudo dos fósseis de animais, e a paleobotânica, o estudo dos fósseis de plantas.[11][12] Ambas se fundamentam em fósseis para compreender a modificação dos organismos vivos ao longo do tempo e das gerações pela evolução.
Fósseis conservados em resina são um bom exemplo de mumificação (ou conservação) que permitem verificar como eram os organismos vivos no passado.[13] Animais menores, como insetos, aranhas e pequenos lagartos, quando presos em resina segregada por certas árvores, ficam praticamente intactos por milhões de anos. A produção de resinas remonta ao Paleozóico, no período Carbonífero, mas a partir do Triássico encontra-se maior abundância de âmbar no registro geológico.[13] Com isso, para uma verificação da história das formas de vida anteriores a essa preservação em âmbar, os paleontólogos recorrem aos registros fósseis também presentes nas rochas.
Registro fóssil
É possível descobrir como um grupo de organismos evoluiu arrumando seu registro fóssil em uma sequência cronológica. Tal sequência pode ser determinada porque fósseis são maioritariamente encontrados em rochas sedimentares. Rochas sedimentares são formadas por camadas de silte ou lama uma sobre o topo da outra; deste modo, a rocha resultante desse processo contém uma série de camadas horizontais, ou estratos. Cada camada contém fósseis que são típicos para o específico período de tempo durante o qual eles se formaram. Os estratos mais baixos contêm as rochas mais velhas e os fósseis mais antigos, enquanto os estratos superiores contêm as rochas mais novas e os fósseis mais recentes.
Uma sucessão de animais e plantas também pode ser vista no registro fóssil. Evidências fósseis apoiam a teoria de que organismos tendem a aumentar progressivamente em complexidade. Ao estudar o número e complexidade de diferentes fósseis em diferentes níveis estratígrafos, foi demonstrado que rochas antigas que contêm fósseis apresentam poucos tipos de organismos fossilizados, e todos eles têm uma estrutura simples, enquanto que as rochas mais recentes contêm uma grande variedade de fósseis, frequentemente com um aumento na complexidade de suas estruturas.
No passado, as idades dos vários estratos e dos fósseis encontrados eram irregularmente estimadas por geologistas. Eles faziam, por exemplo, a estimação do tempo para a formação das rochas sedimentares, camada por camada. Hoje, realizando medições das proporções de elementos radioativos e estáveis na rocha, as idades dos fósseis podem ser datadas pelos cientistas com uma grande precisão. Essa técnica é conhecida como datação radiométrica.

Pelo do registro fóssil, muitas espécies que aparecem em um nível estratígrafo antigo, desaparecem em níveis posteriores. Isso é interpretado em termos evolucionários como a indicação do tempo em que uma espécie se originou e tornou-se extinta. Regiões geográficas e condições climáticas variaram consideravelmente através da história da Terra. Já que organismos se adaptam a ambientes em particular, a mudança constante das condições climáticas e geográficas favoreceu espécies que se adaptaram a novos ambientes através do mecanismo de seleção natural.
Limitações do registro
O registro fóssil é uma fonte importante para cientistas na investigação da história evolucionária dos organismos. Entretanto, devido a limitações inerentes ao registro fóssil, não existe uma boa sequência de formas intermediárias entre grupos relacionados de espécies. Essa falta de uma sequência contínua de fósseis (de espécies para espécies) no registro é uma grande limitação na investigação da descendência de grupos biológicos. Além disso existem lacunas entre certas linhagens evolutivas. Os fósseis que “tapariam” as lacunas são frequentemente chamados de “elos perdidos”. Esses elos perdidos são esporadicamente encontrados em escavações paleontológicas gradativamente melhorando o registro fóssil.
Existe uma lacuna de aproximadamente 100 milhões de anos entre o período do Cambriano antigo e o período Ordoviciano posterior, O período do Cambriano antigo foi o período onde numerosos fósseis de esponjas, cnidarias, equinodermos, moluscos e artrópodes são encontrados. No período Ordoviciano posterior, o primeiro animal que realmente possuí as características de um vertebrado(um peixe) apareceu. Portanto não existe uma sequencia impecável de fósseis intermediários entre invertebrados e vertebrados.
Alguma das razões para a imperfeição do registro fóssil são:
- Em geral, a probabilidade de um organismo fossilizar-se depois de morto é bem baixa;
- Algumas espécies ou grupos têm menos chance de tornarem-se fósseis porque apresentam corpos moles;
- Algumas espécies ou grupos têm menos chance de tornarem-se fósseis, porque eles vivem (e morrem) em condições que não favorecem a fossilização;
- Muitos fósseis são destruídos por movimentos de terra e pela erosão;
- Alguns fósseis são completos, mas a maioria é fragmentada;
- Algumas mudanças evolucionárias ocorrem no limite do alcance ecológico de uma espécie, e como essas espécies provavelmente são pequenas, a probabilidade da fossilização é baixa (Equilíbrio pontuado);
- Similarmente, quando condições ambientais mudam, a população de um espécie provavelmente se reduzirão em grande quantidades, logo qualquer adaptação as condições tem chances baixas de ser fossilizada;
- A maioria dos fósseis apresenta informações sobre a forma externa, mas muito pouco sobre como o organismo funcionava;
- Usando a biodiversidade atual com um guia, supõem-se que os fósseis descobertos representam só uma fração do real número de organismos que viveram no passado.
Gradualismo
O gradualismo é a ideia de que as mudanças evolutivas ocorrem de forma lenta, contínua e acumulativa ao longo do tempo. Segundo essa visão, novas espécies surgem por meio da acumulação gradual de pequenas variações genéticas, sem saltos bruscos. Essa teoria foi defendida por Charles Darwin e Charles Lyell, e contrasta com o equilíbrio pontuado, que propõe períodos longos de estabilidade interrompidos por mudanças rápidas.[14]
Evolução gradual do cavalo
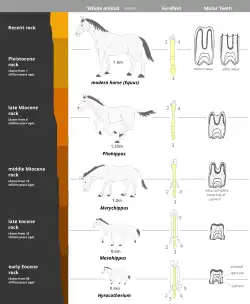
Devido à descoberta de um registro fóssil praticamente completo encontrado nos depósitos sedimentares da América do Norte do período antigo do Eoceno até o presente, o cavalo apresenta um dos melhores exemplos de história evolucionária (filogenia).
Essa história evolucionária começa com um pequeno animal chamado Hyracotherium que viveu na América do Norte aproximadamente há 54 milhões de anos e espalhou-se pela Europa e Ásia. Restos de fósseis do Hyracotherium mostram que ele divergia do cavalo moderno em três importantes aspectos: era um animal pequeno (do tamanho de uma raposa), de estrutura leve e adaptada para correr; os membros eram pequenos e delgados, e patas alongadas fazendo com que os artelhos (dedos do pé) fossem quase verticais, com quatro artelhos nos membros anteriores e três artelhos nos membros traseiros; os incisivos eram pequenos, os molares tinham coroas com cúspides cobertas de esmalte.
O curso provável do desenvolvimento dos cavalos, do Hyracotherium ao Equus (o cavalo moderno) envolveu no mínimo 12 gêneros a centenas de espécies. As maiores correntes vistas no desenvolvimento dos cavalos a mudanças das condições ambientais podem ser resumidas como se segue:
- Aumento no tamanho (de 0,4 m para 1,5 m);
- Alongamento dos membros e patas;
- Redução dos artelhos laterais;
- Aumento em tamanho e espessura do terceiro artelho;
- Aumento em largura dos incisivos;
- Substituição de pré-molares por molares; e
- Aumento do tamanho dos dentes, altura da coroa dos molares.
Plantas fossilizadas em estratos diferentes mostram que o ambiente pantanoso e arborizado no qual o Hyracotherium viveu tornou-se gradualmente seco. Sua sobrevivência dependia de uma posição elevada da cabeça para ganhar uma boa visão do ambiente ao redor, e de uma grande velocidade de rotação para escapar de predadores. Consequentemente, ocorreram o aumento de tamanho e a substituição das patas abertas pelas patas com cascos. O chão mais seco e duro tornaria as patas originais não muito adaptadas para o suporte do animal. As mudanças nos dentes podem ser explicadas presumindo-se que a dieta deles mudou de vegetação macia para grama. Um genus dominante de cada período geológico foi selecionado para mostrar o desenvolvimento progressivo do cavalo. Entretanto, é importante salientar que não há evidências de que as formas ilustradas são diretamente descendentes umas das outras, embora elas sejam relacionadas.
Evidências da anatomia comparada
Estruturas análogas — evolução convergente
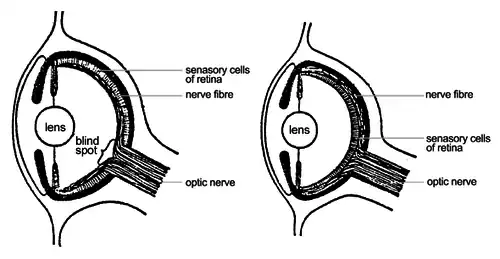
Sobre condições ambientais similares, estruturas fundamentalmente diferentes em diferentes grupos de organismos podem passar por modificações para servirem a funções similares. Esse fenômeno é chamado de evolução convergente. Estruturas similares, processos fisiológicos ou modos de vida em organismos que aparentemente não apresentam ligações filogenéticas próximas, mas que apresentam adaptações para realizar as mesmas funções são descritas como análogas, por exemplo:
- Asas de morcegos, pássaros e insetos;
- as pernas articuladas de insetos e vertebrados;
- nadadeiras do rabo de peixes, baleias e lagostas;
- os olhos dos vertebrados e moluscos cefalópodes (lulas e polvos).
As diferenças entre as retinas invertidas e não invertidas, e as células censoras abaixo do axônio podem ser vistos na figura (em inglês). Isso resulta na ausência de células censoras onde o nervo óptico liga-se ao olho, consequentemente criando um ponto cego. O olho do polvo tem uma retina não invertida logo, as células sensoras ficam sobre o axônio. Portanto não há ponto cego nesse tipo de olho. Fora essa diferença, os dois olhos são incrivelmente similares, um exemplo de evolução convergente.
Estruturas homólogas — evolução divergente
Se grupos bem separados de organismos são originalmente descendentes de um ancestral em comum, é esperado que eles apresentem certas características em comum. O grau de semelhança entre dois grupos de organismos deve indicar o quão próximos eles são no parentesco evolucionário:
- Se assume que grupos com pouco em comum devem ter divergido-se de um ancestral em comum muito cedo na história geológica do que grupos que tem muito em comum;
- Na decisão do grau de parentesco de dois animais, um anatomista comparativo procura por estruturas que, embora sirvam propósitos diferentes no adulto funcional de cada espécie, são fundamentalmente similares, sugerindo um origem em comum. Tais estruturas são descritas como homólogas; e no caso de estruturas similares que diferem em funções nos adultos, é talvez necessário investigar suas origens e desenvolvimento embriológico, para procurar por mais similaridades de um ancestral em comum.
Quando um grupo de organismos compartilha uma estrutura homóloga que é especializada para realizar uma variedade de funções e adaptar-se a diferentes condições ambientais e modos de vida, é chamado de irradiação adaptativa. A difusão gradual de organismos com irradiação adaptativa é conhecida como evolução divergente.
Membros pentadáctilos
A disposição dos ossos dos membros chamados de pentadáctilos é um exemplo de estruturas homologas. É encontrado em todas as classes de tetrápodes (de anfíbios a mamíferos). Pode até ser ligado as nadadeiras de certos fósseis de peixes dos quais os anfíbios teriam evoluído. O membro tem um único osso proximal (úmero), dois ossos distais (rádio e a ulna), uma série de carpos (ossos do pulso), seguido por uma série de metacarpos (ossos da palma) e falanges (dedos). Através dos tetrápodes, as estruturas fundamentais dos membros pentadáctilos são os mesmos, indicando que eles originaram-se de um ancestral em comum. Mas através do curso da evolução, estas estruturas fundamentais foram se modificando. Eles se tornaram estruturas superficialmente diferentes e aparentemente não relacionadas, que serviam a diferentes funções devido a adaptação a diferentes ambientes e modos de vida. Tal fenômeno é claramente evidenciado nos membros anteriores dos mamíferos. Por exemplo:
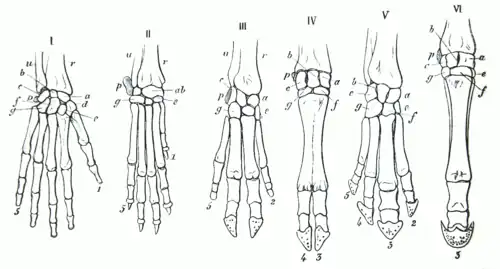
- Nos macacos, os membros anteriores são muito mais alongados o que permite uma melhor adaptação para escalar e balançar entre árvores;
- Nos porcos, o primeiro dedo foi perdido, o segundo e quinto dedo reduziram-se. Os dois dedos remanescentes são mais longos e robustos que o resto e têm um casco para dar suporte ao corpo;
- No cavalo, os membros anteriores são adaptados para dar suporte e favorecer corridas devido a uma grande elongação do terceiro dedo que apresenta um casco;
- A toupeira tem um par de membros adaptados para cavar e fazer tocas;
- O tamanduá usa seu terceiro dedo alongado para destruir formigueiros e cupinzeiros;
- Na baleia, os membros anteriores tornaram-se nadadeiras usadas na navegação e equilíbrio durante o nado;
- No morcego, os membros anteriores se tornaram asas para o voo devido a grande elongação dos quatro dedos, e um dedo em formato de gancho permaneceu livre para se segurar em árvores.
Outro aspecto da anatomia comparada é a presença de estruturas vestigiais que podem ser órgãos. Órgãos que são menores e mais simples em sua estrutura do que as partes correspondentes nas espécies ancestrais. Eles são normalmente degenerados ou subdesenvolvidos. A existência de órgãos vestigiais pode ser explicada em termos de mudanças no ambiente ou modos de vida da espécie. Supõe-se que esses órgãos eram funcionais em espécies ancestrais, mas tornaram-se não funcionais ou desnecessários. Exemplos são os membros traseiros de baleias, as asas traseiras vestigiais de moscas e mosquitos, as asas vestigiais de aves que não voam, como avestruzes, o osso cóccix no ser humano, e folhas vestigiais de algumas xeromorfas (cactus) e de plantas parasitas (cuscuta).
Estruturas Rudimentares
Embora os termos estrutura vestigial e estrutura rudimentar sejam frequentemente usados como sinônimos, há uma clara distinção entre eles.[15] Estruturas rudimentares são parcialmente transformações formadas ou incompletas de uma característica de desenvolvimento e são encontradas apenas em embriões, enquanto estruturas vestigiais são remanescentes evolutivos de uma característica ancestral e são encontradas em adultos[15]. De forma grosseira, o rudimento surge e depois é retido pela ação de um gene, enquanto o vestígio dura por toda a vida do indivíduo. Acontece que uma mutação casual pode desencadear uma extensão do desenvolvimento desse rudimento embrionário. A presença de um rudimento embrionário é em parte a consequência do compartilhamento de genes reguladores entre diferentes tecidos e órgãos e a dificuldade de remover inteiramente um primórdio. No último caso, se o desenvolvimento do rudimento não for interrompido pela ação de um gene, o indivíduo nasce com a estrutura atávica.[16]
Estruturas Atávicas (atavismo)
Atavismo ("ancestral") é o reaparecimento de uma certa característica no organismo depois de várias gerações de ausência. Em biologia, atavismo é uma reminiscência evolutiva, como reaparecimento de traços que tiveram ausentes em várias gerações. Pode ocorrer de várias maneiras. Uma maneira é quando genes para características previamente fenotípicas existentes são preservadas no DNA, e estes tornam-se expressar através de uma mutação que quer nocautear os genes primordiais para os novos traços ou fazer os traços antigos substituírem os atuais.
Relações Anatômicas
As estruturas rudimentares, vestigiais e atávicas estão intimamente relacionadas do ponto de vista anatômico e evolutivo, pois todas se referem a características corporais que revelam traços da história evolutiva dos organismos. Veja como se relacionam:
| Estrutura Rudimentar (embrionária) | Estrutura Vestigial (permanente) | Estrutura Atávica (atavismo) |
|---|---|---|
| Cauda no embrião Homo sapiens por um período de 4 semanas [17] | Cauda óssea atrofiada denominada cóccix [18] | Cauda atávica com vértebras (truetail). [19][20][21][22][23] |
| Membro inferior em embrião de cobra Python próximo à cloaca [24] | Perna vestigial denominada esporão pévico [25] | Perna atávica em cobras sem esporão pélvico. [26] |
| Membro traseiro em embriões de golfinhos (Stenella attenuata) [27] | Pelve vestigial em golfinhos e baleias [28] | Barbatanas traseiras em baleias e golfinhos. [29][30][31] |
| Múltiplos dígitos em embriões de Equus caballus de 29-35 dias [32] | Ossos denominados como ossos da tala [33] | Dedos laterais atávicos em cavalos. [29][30][34][35][36] |
| Cauda longa em embriões de aves como frangos (Gallus gallus) [37] | Cauda fusionada denominada pigóstilo [38] | — ainda não documentado na literatura — |
| Dentição em fetos de baleias modernas desdentadas Mysticeti [39] | Epitélio com alvéolos dentários vestigiais [40][41] | — ainda não documentado na literatura — |
| — ainda não documentado na literatura — | Genes vestigiais para a odontogênese [42][43] | Dentição em embriões de frangos (Gallus gallus). [42][43] |
| Arcos e fendas branquiais iniciais em embriões Homo sapiens [44] | Seio Pré-Auricular próximo das orelhas [45][46] | Fenda branquial em pescoço de recém-nascido. [47] |
| Esboço rudimentar da membrana nictitante em Homo sapiens [48] | Membranas chamadas pregas semilunares [49] | Membranas nictitantes em olhos de duas crianças. [50][51] |
| Botões pela linha mamária enquanto embrião Homo sapiens [52][53] | Mama masculina; [54] mama axilar feminina [55] | Mamilos supranumerários em ambos. [30][56][57][58][59] |
Evidência da distribuição geográfica
Distribuição continental
Todos os organismos são adaptados ao seu ambiente a um maior ou menor grau. Se os fatores abióticos e bióticos dentro de um habitat são capazes de sustentar uma espécie em particular em uma área geográfica, então se supõem que a mesma espécie seria encontrada em um habitat similar em uma área geográfica também similar, por exemplo, África e América do Sul. Mas não é isso que ocorre. Plantas e animais são descontinuamente distribuídos através do mundo:
- África tem macacos com cauda mais curta (Velho Mundo), elefantes, leões e girafas;
- América do Sul tem macacos de cauda longa, pumas, jaguares e lhamas.
Diferenças ainda maiores podem ser evidenciadas se a Austrália é tomada em consideração embora ocupe a mesma latitude da América do Sul e da África. Marsupiais como o canguru podem ser encontrados na Austrália, mas são totalmente ausentes na África e só são representados pelo gambá na América do Sul e o gambá da Virginia na América do Norte:
- O equidna e o ornitorrinco, um dos únicos representantes dos primitivos mamíferos que colocavam ovos (monotremata), são encontrados somente na Austrália, e são ausentes no resto do mundo;
- Entretanto, a Austrália tem poucos mamíferos placentários, exceto aqueles que foram introduzidos pelo homem.
Explanação
Os grandes grupos de mamíferos modernos originaram-se no Hemisfério Norte e subsequentemente migraram para três direções principais:
- Para a América do Sul através de pontes de terra (Estreito de Bering e Istmo do Panamá); um grande número de famílias sul americanas de marsupiais tornaram-se extintas como resultado direto da competição com suas contrapartes norte americanas;
- Para a África através do Estreito de Gibraltar e para Austrália pelo Sudeste da Ásia que já foi no passado ligado por terra.
A superficialidade do Estreito de Bergin teria feita a passagem de animais entre os continentes do norte um processo relativamente raso, e explica a similaridade atual entre as duas faunas. Mas uma vez que houve migrações para os continentes ao sul, as espécies presumivelmente ficaram isoladas uma das outras por vários tipo de barreiras.
- A submersão do Istmo do Panamá: Isolou a fauna da América do Sul;
- O Mar Mediterrâneo e o Deserto Norte africano: isolam parcialmente a fauna africana; e
- A submersão da conexão original entre a Austrália e o Sudeste da Ásia: isolou a fauna australiana.
Uma vez isolados, os animais em cada continente apresentaram radiação adaptativa para evoluir ao longo de suas linhagens.
Evidência para migração e isolamento
O registro fóssil dos camelos indica que a evolução deles começou na América do Norte, de onde eles migraram pelo Estreito de Bering para a Ásia e consequentemente África, e através do istmo do Panamá para a América do Sul. Uma vez isolados, eles evoluíram nas suas linhagens, apresentando o camelo moderno na Ásia e na África e os lamas na América do Sul.
Deriva continental
Os mesmos grupos de fósseis são encontrados em áreas que foram adjacentes umas as outras no passado mas que, através do processo de deriva continental, são agora locais geográficos amplamente diferentes. Por exemplo, fósseis do mesmo grupo de anfíbios antigos, artrópodes e pteridófitas são encontradas na América do Sul, África, Índia, Austrália e Antártica, os quais podem ser datados ara Era Paleozóica, e que nessa época essas regiões eram unidas em uma única massa de terra chamada Gondwana [4]. Algumas vezes os descendentes desses organismos podem ser identificados com similaridades irrefutáveis um com o outro, mesmo que eles habitem regiões e climas bem diferentes.
Distribuição de ilhas oceânicas
Muitos dos animais em pequenas ilhas isoladas só têm espécies nativas que só poderiam chegar nelas pelo mar ou ar; como pássaros, insetos e tartarugas. Os poucos mamíferos grandes presentes hoje nessas ilhas foram trazidos a elas por colonizadores em barcos. Plantas em ilhas vulcânicas remotas como o Havaí poderiam ter chegado por esporos ou sementes trazidas por pássaros.
Evidências da embriologia comparativa
A Embriologia comparativa mostra como embriões começam parecendo o mesmo. Durante seus respectivos desenvolvimentos, suas similaridades decrescem vagarosamente até que eles tomem as formas de suas classes particulares.
Por exemplo, vertebrados adultos são diversos, entretanto seus embriões são bem similares em estágios iniciais. Estruturas similares a estruturas encontradas em peixes ainda se formam em estágios inicias de desenvolvimento de embriões de répteis, pássaros e mamíferos. Em embriões de peixes, um coração de duas câmaras, algumas veias, e partes de artérias desenvolvem-se e persistem em peixes adultos. As mesmas estruturas formaram-se em estágios inicias do desenvolvimento de embriões humanos, mas não persistem nos adultos.
Evidências da filogenética comparada e bioquímica
Veja também: Evidência da Macroevolução, Parte 4: Evidencia da Sequência Molecular
Evolução de moléculas e proteínas amplamente distribuídas
Todos os organismos existentes fazem uso do DNA e ou RNA. O ATP é usado como a “moeda” metabólica por toda a vida existente. O código genético é o mesmo para quase todos os organismos, significando que um pedaço de RNA em uma bactéria codifica para uma mesma proteína em uma célula humana.
Um exemplo clássico de evidência bioquímica para a evolução é a variação da proteína citocroma c em células vivas. A diferença da citocroma c em diferentes organismo é medida na diferença de aminoácidos, cada amino diferente sendo resultado de uma substituição de um par de bases, uma mutação. Se cada amino diferente é considerado como o resultado de uma substituição de um par de base, pode-se então calcular a quanto tempo atrás duas espécies divergiram realizando a multiplicação dos números de substituição de pares de base pelo tempo estimado que leva para uma substituição de pares de base comum do citocroma c ser passada adiante.
Por exemplo, o tempo normal que um par de bases do citocroma c leva para mutar é n anos, o número de aminoácidos que fazem a proteína citocroma c nos macacos, difere por um do citocroma c dos humanos, isso leva a conclusão de que as duas espécies divergiram n anos atrás.
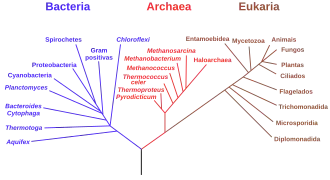
Comparações de sequências de DNA permitem o agrupamento de organismos pelo critério de similaridades entre as sequências, resultando em árvores filogenéticas tipicamente congruentes com a taxonomia tradicional, e são frequentemente usadas para fortalecer ou corrigir classificações taxonômicas. A comparação de sequências é considerada uma medida robusta o suficiente para corrigir suposições errôneas sobre árvores filogenéticas em casos quando outras evidências são raras. Por exemplo, as sequências de DNA humano neutro divergem aproximadamente 1,2% (baseado na substituição) daqueles de seus parentes mais próximos, os chimpanzés, 1,6% dos gorilas, e 6,6% dos babuínos[60]. A evidência das sequências genéticas permite inferir a quantificação do parentesco entre humanos e outros primatas.[61][62] A sequência do gene 16S rRNA, um componente vital do ribossomo, foi usado para encontrar um parentesco filogenético geral entre toda a vida existente. A analise, originalmente feita por Carl Woese, que resultou no sistema de três domínios, argumentando por duas grandes separações no início da história evolutiva da vida. A primeira divisão para a Bacteria moderna e a divisão subsequente para Archaea e Eukaryota modernos.
A evidência proteômica também apóia uma ancestralidade universal da vida. Proteínas vitais, como os ribossomos, DNA polimerase, e o RNA polimerase são encontrados desde as (quase todas) as bactérias primitivas até os complexos mamíferos. A região do núcleo da proteína é conservada através de todas as linhagens da vida, servindo funções similares. Organismos mais complexos evoluíram subunidades de proteínas adicionais, afetando largamente a regulação da interação de proteína a proteína no núcleo. Outras similaridades entre todas as linhagens da vida, como DNA, RNA, e a bicamada de lipídios, apoiam a teoria da descendência em comum. A quiralidade do DNA, RNA e aminoácidos é conservado em todas as linhagens da vida. Como não há nenhuma vantagem funcional para quiralidade tanto para a direita quanto para a esquerda nas moléculas, a hipótese mais simples é que a escolha foi feita aleatoriamente no início do desenvolvimento da vida e foi um traço que passou para toda a vida existente através da descendência em comum.
Evidência moleculares também oferecem mecanismos para grandes saltos evolucionários e para macroevolução. A transferência gênica horizontal, o processo pelo qual um organismo transfere ou mistura material genético (DNA, por exemplo) para outra célula que não seja seu progênito, permite a ocorrência de saltos evolucionários repentinos realizando incorporações benéficas de genes que evoluíram em outra espécie. A Teoria da Endossimbiose explica a origem da mitocôndria e do plastídeo (por exemplo, cloroplastos), que são organelas de células eucarióticas, como a incorporação de uma célula procariótica antiga dentro de uma célula eucariótica. Ao invés de uma lenta evolução das organelas, essa teoria oferece um mecanismo de um repentino salto evolucionário já que ocorre a incorporação de material genético e composição bioquímica de uma espécie separada. Evidências suportando esse mecanismo foram recentemente encontradas no protista Hatena arenicola já que como um predador ele engolfa uma célula de alga verde, que subsequentemente comporta-se como um endossimbionte, alimentando a Hatena, que em troca perdeu seu aparatos de alimentação e comporta-se como um autótrofo.[63]
Mais evidências para a reconstrução de linhagens ancestrais vem do DNA lixo como os pseudogenes, i.e., genes “mortos”, que acumulam mutações com regularidade.[64]
Já que processos metabólicos não deixam fósseis, a pesquisa da evolução dos processos celulares básicos é feita comparando-se organismos existentes. Muitas linhagens divergem quando um novo processo metabólico surge, e é teoricamente possível determinar quando certos processos metabólicos surgem realizando a comparação de traços dos descendentes de um ancestral em comum ou na detecção da manifestação física desse processo. Como um exemplo, o aparecimento de oxigênio na atmosfera da Terra está ligado à evolução da fotossíntese.
Evidências da especiação
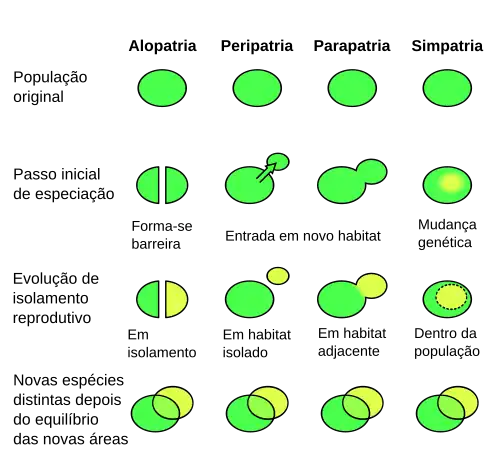
Um caso interessante de evolução ocorrendo é o caso da mosca da fruta, Rhagoletis pomonella, que parece estar passando pelo processo de especiação simpátrica.[65] Populações diferentes da mosca da fruta alimentam-se de diferentes frutas. Uma população distinta surgiu na América do Norte no século XIX algum tempo depois que as maçãs, uma espécie não nativa, que foi introduzida no continente. Essa população que se alimenta de maçãs, alimenta-se somente delas e não da fruta de espinheiros. A população “normal” de moscas da fruta não se alimenta normalmente com maçãs. Cientistas ainda estão pesquisando se a subespécie que se alimenta de maçãs vai ou não evoluir em uma nova espécie.
Algumas evidências, como o fato de que as moscas desenvolvem-se mais tarde na estação e levam mais tempo para desenvolver do que moscas da maçã; e existem poucas evidências sugerindo que inter-reprodução (pesquisas tem documentado uma taxa de 4-6% de hibridação) esteja ocorrendo. A ocorrência da mosca da fruta é um exemplo de evolução em progresso.[66]
Referências
- ↑ Doebley, John (1 de julho de 1990). «Molecular Evidence and the Evolution of Maize». Economic Botany (em inglês) (3): 6–27. ISSN 1874-9364. doi:10.1007/BF02860472. Consultado em 14 de novembro de 2025
- ↑ Sober, Elliott (27 de março de 2008). Evidence and Evolution: The Logic Behind the Science (em inglês). [S.l.]: Cambridge University Press. Consultado em 14 de novembro de 2025
- ↑ Land, Judy; Lang, Mark H. (1 de março de 2002). «Empirical Evidence on the Evolution of International Earnings». The Accounting Review (em inglês) (s-1): 115–133. ISSN 0001-4826. doi:10.2308/accr.2002.77.s-1.115. Consultado em 14 de novembro de 2025
- ↑ Adams, Greg D. (1997). «Abortion: Evidence of an Issue Evolution». American Journal of Political Science (3): 718–737. ISSN 0092-5853. doi:10.2307/2111673. Consultado em 14 de novembro de 2025
- ↑ a b Lister, Adrian M. (junho de 2014). «Behavioural leads in evolution: evidence from the fossil record: Behavioural Leads in the Fossil Record». Biological Journal of the Linnean Society (em inglês) (2): 315–331. doi:10.1111/bij.12173. Consultado em 14 de novembro de 2025
- ↑ Muller, Gerd B.; Newman, Stuart A. (3 de janeiro de 2003). Origination of Organismal Form: Beyond the Gene in Developmental and Evolutionary Biology (em inglês). [S.l.]: MIT Press. Consultado em 14 de novembro de 2025
- ↑ Osawa, S; Jukes, T H; Watanabe, K; Muto, A (março de 1992). «Recent evidence for evolution of the genetic code». Microbiological Reviews (1): 229–264. PMC 372862
 . PMID 1579111. doi:10.1128/mr.56.1.229-264.1992. Consultado em 14 de novembro de 2025
. PMID 1579111. doi:10.1128/mr.56.1.229-264.1992. Consultado em 14 de novembro de 2025
- ↑ Petrov, D. A. (1 de maio de 2000). «Pseudogene evolution and natural selection for a compact genome». Journal of Heredity (3): 221–227. doi:10.1093/jhered/91.3.221. Consultado em 14 de novembro de 2025
- ↑ Woese, Carl (9 de junho de 1998). «The universal ancestor». Proceedings of the National Academy of Sciences (12): 6854–6859. PMC 22660. PMID 9618502. doi:10.1073/pnas.95.12.6854. Consultado em 14 de novembro de 2025
- ↑ Schopf, J. William (29 de junho de 2006). «Fossil evidence of Archaean life». Philosophical Transactions of the Royal Society B: Biological Sciences (em inglês) (1470): 869–885. ISSN 0962-8436. PMC 1578735. PMID 16754604. doi:10.1098/rstb.2006.1834. Consultado em 14 de novembro de 2025
- ↑ a b Silva, C.M da (2005), p.45
- ↑ Erro de citação: Etiqueta
<ref>inválida; não foi fornecido texto para as "refs" nomeadaspaleoevo - ↑ a b Erro de citação: Etiqueta
<ref>inválida; não foi fornecido texto para as "refs" nomeadaslivropaleogeral2 - ↑ Torre Sáinz, I. de la, & Domínguez-Rodrigo, M. (2009). Gradualismo y equilibrio puntuado en el origen del comportamiento humano. Zephyrvs, 51. 2009.
- ↑ a b Hall, Brian K. (agosto de 2003). «Descent with modification: the unity underlying homology and homoplasy as seen through an analysis of development and evolution». Biological Reviews (3): 409–433. doi:10.1017/S1464793102006097. Consultado em 24 de novembro de 2021
- ↑ Hall, B. K. (fevereiro de 1984). «Development mechanisms underlying the formation of atavisms». Biological Reviews of the Cambridge Philosophical Society (1): 89–124. ISSN 1464-7931. PMID 6367843. doi:10.1111/j.1469-185x.1984.tb00402.x. Consultado em 24 de novembro de 2021
- ↑ Fallon, John F.; Simandl, B. Kay (1978). «Evidence of a role for cell death in the disappearance of the embryonic human tail». American Journal of Anatomy (em inglês) (1): 111–129. ISSN 1553-0795. doi:10.1002/aja.1001520108. Consultado em 24 de novembro de 2021
- ↑ Mukhopadhyay, Biswanath; Shukla, RamM; Mukhopadhyay, Madhumita; Mandal, KartikC; Haldar, Pankaj; Benare, Abhijit (2012). «Spectrum of human tails: A report of six cases». Journal of Indian Association of Pediatric Surgeons (em inglês) (1). 23 páginas. ISSN 0971-9261. doi:10.4103/0971-9261.91082. Consultado em 24 de novembro de 2021
- ↑ Shad, Jimmy; Biswas, Rakesh (18 de abril de 2012). «An infant with caudal appendage». BMJ Case Reports (em inglês): bcr1120115160. ISSN 1757-790X. PMC 3339178. PMID 22604513. doi:10.1136/bcr.11.2011.5160. Consultado em 10 de novembro de 2025
- ↑ Tomić, Nenad; Victor Benno (18 de agosto de 2011). «Atavisms: Medical, Genetic, and Evolutionary Implications». Perspectives in Biology and Medicine. 54 (3): 332–353. ISSN 1529-8795. doi:10.1353/pbm.2011.0034
- ↑ Lu, Frank L; Pen-Jung. «The human tail». Pediatric Neurology. 19 (3): 230–233. doi:10.1016/s0887-8994(98)00046-0
- ↑ Tomić, Nenad; Meyer-Rochow, Victor Benno (18 de agosto de 2011). «Atavisms: Medical, Genetic, and Evolutionary Implications». Perspectives in Biology and Medicine. 54 (3): 332–353. ISSN 1529-8795. doi:10.1353/pbm.2011.0034
- ↑ «Human Pathology». www.humanpathol.com (em inglês). Consultado em 20 de abril de 2017
- ↑ Infante, Carlos R.; Mihala, Alexandra G.; Park, Sungdae; Wang, Jialiang S.; Johnson, Kenji K.; Lauderdale, James D.; Menke, Douglas B. (12 de outubro de 2015). «Shared Enhancer Activity in the Limbs and Phallus and Functional Divergence of a Limb-Genital cis-Regulatory Element in Snakes». Developmental Cell (em inglês) (1): 107–119. ISSN 1534-5807. PMID 26439399. doi:10.1016/j.devcel.2015.09.003. Consultado em 29 de novembro de 2021
- ↑ Gillingham, James C.; Chambers, Jeffrey A. (1982). «Courtship and Pelvic Spur Use in the Burmese Python, Python molurus bivittatus». Copeia (1): 193–196. ISSN 0045-8511. doi:10.2307/1444292. Consultado em 29 de novembro de 2021
- ↑ Mehrtens JM. 1987. Living Snakes of the World in Color. New York.
- ↑ Bejder, Lars; Hall, Brian K. (2002). «Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss». Evolution & Development (em inglês) (6): 445–458. ISSN 1525-142X. doi:10.1046/j.1525-142X.2002.02033.x. Consultado em 29 de novembro de 2021
- ↑ Dines, James P.; Otárola-Castillo, Erik; Ralph, Peter; Alas, Jesse; Daley, Timothy; Smith, Andrew D.; Dean, Matthew D. (2014). «Sexual selection targets cetacean pelvic bones». Evolution (em inglês) (11): 3296–3306. ISSN 1558-5646. PMID 25186496. doi:10.1111/evo.12516. Consultado em 29 de novembro de 2021
- ↑ a b Erro de citação: Etiqueta
<ref>inválida; não foi fornecido texto para as "refs" nomeadas:0 - ↑ a b c Hall, Brian K. (1 de fevereiro de 1984). «Developmental Mechanisms Underlying the Formation of Atavisms». Biological Reviews (em inglês). 59 (1): 89–122. ISSN 1469-185X. doi:10.1111/j.1469-185X.1984.tb00402.x
- ↑ TalkOrigins Archive. «29+ Evidences for Macroevolution: Part 2». Consultado em 8 de novembro de 2006. Cópia arquivada em 29 de outubro de 2006
- ↑ Kavanagh, Kathryn D.; Bailey, C. Scott; Sears, Karen E. (12 de fevereiro de 2020). «Evidence of five digits in embryonic horses and developmental stabilization of tetrapod digit number». Proceedings of the Royal Society B: Biological Sciences (1920). 20192756 páginas. PMID 32019446. doi:10.1098/rspb.2019.2756. Consultado em 29 de novembro de 2021
- ↑ Miller, Robert M. (1999). Understanding the Ancient Secrets of the Horse's Mind. Neenah, WI: Russell Meerdink Company Ltd. ISBN 978-0-929346-65-6. OCLC 42389612.
- ↑ Simpson, G. G. (1951), Horses: The story of the horse family in the modern world and through sixty million years of evolution, Oxford University Press
- ↑ Tyson, Reid; Graham, John P.; Colahan, Patrick T.; Berry, Clifford R. (1 de julho de 2004). «Skeletal Atavism in a Miniature Horse». Veterinary Radiology & Ultrasound (em inglês). 45 (4): 315–317. ISSN 1740-8261. doi:10.1111/j.1740-8261.2004.04060.x
- ↑ Carstanjen, Bianca; Abitbol, Marie; Desbois, Christophe (1 de junho de 2007). «Bilateral Polydactyly in a foal». Journal of Veterinary Science (em inglês) (2): 201–203. ISSN 1229-845X. PMID 17519577. doi:10.4142/jvs.2007.8.2.201. Consultado em 6 de abril de 2024
- ↑ Rashid, Dana J.; Chapman, Susan C.; Larsson, Hans CE; Organ, Chris L.; Bebin, Anne-Gaelle; Merzdorf, Christa S.; Bradley, Roger; Horner, John R. (29 de julho de 2014). «From dinosaurs to birds: a tail of evolution». EvoDevo (1). 25 páginas. ISSN 2041-9139. doi:10.1186/2041-9139-5-25. Consultado em 29 de novembro de 2021
- ↑ Fastovsky, David E.; Weishampel, David B. (7 de fevereiro de 2005). The Evolution and Extinction of the Dinosaurs (em inglês). [S.l.]: Cambridge University Press
- ↑ Deméré, Thomas A.; McGowen, Michael R.; Berta, Annalisa; Gatesy, John (fevereiro de 2008). «Morphological and molecular evidence for a stepwise evolutionary transition from teeth to baleen in mysticete whales». Systematic Biology (1): 15–37. ISSN 1063-5157. PMID 18266181. doi:10.1080/10635150701884632. Consultado em 29 de novembro de 2021
- ↑ McGowen, Michael R.; Gatesy, John; Wildman, Derek E. (1 de junho de 2014). «Molecular evolution tracks macroevolutionary transitions in Cetacea». Trends in Ecology & Evolution (em inglês) (6): 336–346. ISSN 0169-5347. PMID 24794916. doi:10.1016/j.tree.2014.04.001. Consultado em 29 de novembro de 2021
- ↑ Deméré, Thomas A.; McGowen, Michael R.; Berta, Annalisa; Gatesy, John (1 de fevereiro de 2008). «Morphological and Molecular Evidence for a Stepwise Evolutionary Transition from Teeth to Baleen in Mysticete Whales». Systematic Biology (1): 15–37. ISSN 1076-836X. doi:10.1080/10635150701884632. Consultado em 29 de novembro de 2021
- ↑ a b Adams, Jill U.; Kenna M. Shaw (2008). «Atavism: Embryology, Development and Evolution». Atavism: Embriology, Development and Evolution. Consultado em 14 de abril de 2011
- ↑ a b Perle, M A; Newman, S A (agosto de 1980). «Talpid2 mutant of the chicken with perturbed cartilage development has an altered precartilage-specific chromatin protein.». Proceedings of the National Academy of Sciences (em inglês) (8): 4828–4830. ISSN 0027-8424. PMC 349940. PMID 6933534. doi:10.1073/pnas.77.8.4828. Consultado em 13 de novembro de 2025
- ↑ Casale, Jarett; Giwa, Al O. (2025). «Embryology, Branchial Arches». Treasure Island (FL): StatPearls Publishing. PMID 30860722. Consultado em 13 de novembro de 2025
- ↑ Adegbiji, W. A.; Alabi, B. S.; Olajuyin, O. A.; Nwawolo, C. C. (dezembro de 2013). «Presentation of preauricular sinus and preauricular sinus abscess in southwest Nigeria». International journal of biomedical science: IJBS (4): 260–263. ISSN 1550-9702. PMC 3884798. PMID 24711764. Consultado em 13 de novembro de 2025
- ↑ Tan, T.; Constantinides, H.; Mitchell, T. E. (1 de novembro de 2005). «The preauricular sinus: A review of its aetiology, clinical presentation and management». International Journal of Pediatric Otorhinolaryngology (11): 1469–1474. ISSN 0165-5876. doi:10.1016/j.ijporl.2005.07.008. Consultado em 13 de novembro de 2025
- ↑ Rosa, Rafael Fabiano Machado; Zen, Paulo Ricardo Gazzola; Graziadio, Carla; Paskulin, Giorgio Adriano (fevereiro de 2009). «Síndrome brânquio-óculo-facial (BOFS) e cardiopatias congênitas». Arquivos Brasileiros de Cardiologia (em inglês) (2). ISSN 0066-782X. doi:10.1590/S0066-782X2009000200015. Consultado em 7 de novembro de 2025
- ↑ Arends, Gunna; Schramm, Uda (1 de junho de 2004). «The structure of the human semilunar plica at different stages of its development a morphological and morphometric study». Annals of Anatomy - Anatomischer Anzeiger (3): 195–207. ISSN 0940-9602. doi:10.1016/S0940-9602(04)80002-5. Consultado em 13 de novembro de 2025
- ↑ Montagna, W.; Machida, H.; Perkins, E. M. (1966). «The skin of primates. XXXIII. The skin of the angwantibo (Arctocebus calabarensis)». American Journal of Physical Anthropology (em inglês) (3): 277–290. ISSN 1096-8644. doi:10.1002/ajpa.1330250307. Consultado em 13 de novembro de 2025
- ↑ Heralgi, Mallikarjun; Thallangady, Ashok; Venkatachalam, Kavitha; Vokuda, Hariprasad (2017). «Persistent unilateral nictitating membrane in a 9-year-old girl: A rare case report». Indian Journal of Ophthalmology (em inglês) (3). 253 páginas. ISSN 0301-4738. doi:10.4103/ijo.IJO_436_15. Consultado em 13 de novembro de 2025
- ↑ Ahuja, Nisha; Katara, Pushkal (1 de março de 2024). «Pseudonictitating Membrane». Ophthalmology (em inglês) (3). 393 páginas. ISSN 0161-6420. PMID 37294234. doi:10.1016/j.ophtha.2023.05.015. Consultado em 13 de novembro de 2025
- ↑ Teplica, David; Kovich, Grant; Srock, Jamey; Whitaker, Robert; Jeffers, Eileen; Wagstaff, David A. (14 de outubro de 2021). «Newly Identified Gross Human Anatomy: Eight Paired Vestigial Breast Mounds Run along the Embryological Mammary Ridges in Lean Adults». Plastic and Reconstructive Surgery - Global Open (em inglês) (10): e3863. ISSN 2169-7574. doi:10.1097/GOX.0000000000003863. Consultado em 13 de novembro de 2025
- ↑ Gabriel, Allen (6 de janeiro de 2025). «Breast Anatomy: Overview, Vascular Anatomy and Innervation of the Breast, Breast Parenchyma and Support Structures». Consultado em 13 de novembro de 2025
- ↑ Goyal, Tarang; Bakshi, S. K.; Varshney, Anupam (setembro de 2012). «Seven nipples in a male: World's second case report». Indian Journal of Human Genetics (3): 373–375. ISSN 0971-6866. PMID 23716953. doi:10.4103/0971-6866.108051. Consultado em 13 de novembro de 2025
- ↑ Vidal, Célio da Silva Rocha; Paulinelli, Régis Resende; Freitas Júnior, Ruffo de; Sousa, Juarez Antônio de; Moreira, Marise Amaral Rebouças; Araújo, Luiz Murilo Martins de (2000). «Carcinoma Primário de Mama Acessória Axilar: Relato de Caso». Revista Brasileira de Ginecologia e Obstetrícia: 307–310. ISSN 0100-7203. doi:10.1590/S0100-72032000000500009. Consultado em 13 de novembro de 2025
- ↑ Bertó, Josep; M. Luisa. «Polimastia y tejido mamario accesorio». Piel. 20 (10): 483–484. doi:10.1016/s0213-9251(05)72333-9
- ↑ Universidade Federal do Piauí; Filho, Walberto Monteiro Neiva Eulálio; Neto, Antônio Marques de Medeiros; Universidade Federal do Piauí; Fé, Taíla Sousa de Moura; Centro Universitário Uninovafapi; Coelho, Lívio Portela Cardoso; Medcenter; Junior, Luiz Ayrton Santos (junho de 2018). «Surgical correction of polymastia and polytelia with an uncommon presentation» (PDF). Mastology (2): 106–109. doi:10.29289/2594539420180000362. Consultado em 7 de novembro de 2025
- ↑ Requena, Luis; Sangüeza, Omar (2017). Requena, Luis; Sangüeza, Omar, eds. «Supernumerary Nipple». Cham: Springer International Publishing (em inglês): 47–54. ISBN 978-3-319-45704-8. doi:10.1007/978-3-319-45704-8_6. Consultado em 7 de novembro de 2025
- ↑ Goyal, Tarang; Bakshi, Sk; Varshney, Anupam (2012). «Seven nipples in a male: World′s second case report». Indian Journal of Human Genetics (em inglês) (3). 373 páginas. ISSN 0971-6866. doi:10.4103/0971-6866.108051. Consultado em 7 de novembro de 2025
- ↑ Two sources: 'Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees'. and 'Quantitative Estimates of Sequence Divergence for Comparative Analyses of Mammalian Genomes' "[1] [2]"
- ↑ A foto com a legenda “Human Chromosome 2 and its analog in the apes” no artigo Comparison of the Human and Great Ape Chromosomes as Evidence for Common Ancestry Arquivado agosto 20, 2011 no WebCite é literalmente um retrato do elo nos humanos que liga dois cromossomos separados em primatas não humanos, criando um único cromossomo em humanos. É considerado um “elo perdido”, uma conexão primita-homem é de interesse particular. Embora o termo se refira originalmente a evidências fósseis, essa descoberta também é um traço do passado correspondendo aos seres vivos que quando vivos foram a personificação física desse elo.
- ↑ A reportagem do The New York Times Still Evolving, Human Genes Tell New Story, baseado em um mapa de seleções positivas recentes no genoma humano, afirma que o International HapMap Project está “fornecendo as mais fortes evidências de que o ser humano ainda está evoluindo” e apresenta detalhadamente algumas dessas evidências.
- ↑ Inouye, I. and Okamoto N., 2006. "Hatena arenicola gen. et sp. nov., a Katablepharid Undergoing Probable Plastid Acquisition.". Protist, Vol. 157, 401—419, August 2006
- ↑ Evolução de peseudogenes e seleção natural de um genoma compacto. “[3]”
- ↑ Feder et al (2003). "Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella.". Genetics 163 (3): 939-953.
- ↑ Berlocher, S.H. and G.L. Bush. 1982. An electrophoretic analysis of Rhagoletis (Diptera: Tephritidae) phylogeny. Systematic Zoology 31:136-155; Berlocher, S.H. and J.L. Feder. 2002. Sympatric speciation in phytophagous insects: moving beyond controversy? Annual Review of Entomology 47:773-815; Bush, G.L. 1969. Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera: Tephritidae). Evolution 23:237-251; Prokopy, R.J., S.R. Diehl and S.S. Cooley. 1988. Behavioral evidence for host races in Rhagoletis pomonella flies. Oecologia 76:138-147. Proc. Natl. Acad. Sci. USA - Vol. 94, pp. 11417-11421, October 1997 - Evolution article Selective maintenance of allozyme differences among sympatric host races of the apple maggot fly
Ligações externas
Livro: Evolução
- Em inglês
- «Plos Genetics: Quão importante é a isolação geográfica na especiação?» (em inglês)
- «29+ evidências da macroevolução: O argumento cientifico para a descendência comum». (em inglês)
- «FAQ - fósseis transicionais de vertebrados» (em inglês)
- «Evolução por Seleção Natural». - Uma introdução a lógica da teoria da evolução por seleção natural. (em inglês)
- Em português
- «Biociência.org». - Com vários artigos sobre a teoria da evolução.