Portia fimbriata
Portia fimbriata
| |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Fêmea de P. fimbriata. | |||||||||||||||||||||
| Classificação científica | |||||||||||||||||||||
| |||||||||||||||||||||
Portia fimbriata é uma espécie de aranha saltadora[1] da família Salticidae, encontrada na Austrália e no sudeste da Ásia. As fêmeas adultas têm corpos de 6,8 a 10,5 milímetros de comprimento, enquanto os dos machos adultos têm de 5,2 a 6,5 milímetros de comprimento.[2] Ambos os sexos têm uma carapaça geralmente marrom-escura, quelíceras ("presas") marrom-avermelhadas, uma parte inferior marrom, palpos marrom-escuros com pelos brancos e abdômen marrom-escuro com manchas brancas na parte superior. Ambos os sexos têm marcas finas e tênues e franjas macias de pelos, e as pernas são finas e com franjas. Entretanto, os espécimes da Nova Guiné e da Indonésia têm carapaças marrom-alaranjadas e abdomens amarelados. Em todas as espécies do gênero Portia, o abdome se distende quando a aranha está bem alimentada ou produzindo ovos.
As táticas de caça do gênero Portia são versáteis e adaptáveis. Todas as espécies de Portia possuem táticas de caça instintivas para suas presas mais comuns, mas podem improvisar por tentativa e erro contra presas desconhecidas ou em situações novas, lembrando-se da nova abordagem.[3] Há diferenças nas táticas de caça das populações regionais de P. fimbriata. As do território do norte da Austrália são ruins na caça de aranhas saltadoras e melhores contra aranhas construtoras de teias e contra insetos. A variante do Sri Lanka é razoável contra outras aranhas saltadoras e boa contra aranhas de teia e insetos. Em Queensland a espécie é um excelente predador de outras aranhas saltadoras e de aranhas de teia, mas é ruim contra insetos. A variante de Queensland usa uma técnica exclusiva de "perseguição críptica" que impede que a maioria das presas identifique essa P. fimbriata como predadora, ou mesmo como um animal. Algumas presas de aranhas saltadoras têm defesas parciais contra a técnica de perseguição críptica. Todos os tipos de aranhas presas ocasionalmente contra-atacam, mas todas as espécies de Portia têm defesas muito boas, a começar pela pele especialmente resistente.
Ao encontrar outro indivíduo da mesma espécie, P. fimbriata não utiliza a perseguição críptica, mas exibe movimentos rápidos e suaves. Em Queensland, disputas entre machos são geralmente muito breves e não causam danos. Disputas entre fêmeas são geralmente longas e violentas, e a vencedora pode expulsar a perdedora e comer seus ovos – mas fêmeas vitoriosas de P. fimbriata de Queensland não matam nem comem a fêmea derrotada. Se um macho se exibe para uma fêmea, ela pode fugir ou avançar contra ele.[c] Se o par chegar a um acordo após isso, eles copularão se ela for madura; se ela for subadulta, ele coabitará em seu ninho até que ela termine a muda, e então copularão. P. fimbriata geralmente copula muito mais rápido que outras aranhas saltadoras. Diferentemente de outras espécies de Portia, as fêmeas de P. fimbriata não comem seus parceiros durante o cortejo, nem durante ou após a cópula.
Estrutura e aparência corporal
As fêmeas da aranha saltadora P. fimbriata têm corpos de 6,8 a 10,5 milímetros de comprimento, enquanto os dos machos adultos têm de 5,2 a 6,5 mm.[3] A variedade de Queensland é tipicamente menor do que a variedade do território do norte.[1] O cefalotórax tem cerca de 4 mm de comprimento e 3 mm de largura, e o abdômen tem cerca de 4 mm de comprimento e 2,2 mm de largura.[4] A parte frontal do cefalotórax é grande e angular, e a face é larga, alta e plana.[5] Na Austrália e no Taiwan, ambos os sexos têm uma carapaça geralmente marrom-escura, quelíceras marrom-avermelhadas, uma parte inferior marrom e palpos marrom-escuros com pelos brancos.[4] Ambos os sexos também têm marcas finas e tênues e franjas macias de pelos.[6]
A fêmea tem duas listras brancas que formam um "M" no cefalotórax: essas listras começam no nível dos pedipalpos, passam entre os olhos anteriores laterais e medianos, sobem até os tufos do cefalotórax e se encontram logo acima do par de olhos principais. A metade posterior do cefalotórax do macho tem uma faixa branca ao redor da borda inferior e um sulco branco na parte posterior.[4] Embora os palpos dos machos sejam maiores que os das fêmeas,[7][8] os palpos das fêmeas da P. fimbriata têm uma franja de pelos que os fazem parecer tão grandes quanto os dos machos.[1] Os abdomens de ambos os sexos são marrom-escuros, com manchas brancas na parte superior.[4] As fêmeas da Nova Guiné tem carapaça e quelíceras alaranjadas com marcas de fuligem, palpos principalmente amarelo-claro, pernas marrom-alaranjadas e abdome amarelo-claro.[3]
Movimentação
Quando não está à procura de presas ou parceiros, as espécies de Portia adotam uma postura especial, chamada "postura de repouso críptica", puxando as pernas para perto do corpo e os palpos para trás, ao lado das quelíceras, o que obscurece os contornos desses apêndices. Ao caminhar, todas as espécies de Portia têm um andar lento e descompassado que preserva sua ocultação: pausando frequentemente em intervalos irregulares; agitando as pernas continuamente e os palpos de forma brusca para cima e para baixo; e movendo cada apêndice fora de sincronia com os outros[9]e variando continuamente a velocidade e o tempo.[10] O andar dessa espécie é diferente do de qualquer outra aranha, e esse andar e as franjas da aranha dão a aparência de luz cintilando através do dossel da floresta e refletindo a partir de um pedaço de detrito.[9][6] Em Queensland, P. fimbriata caminha e se agita mais bruscamente e cerca de duas vezes mais lentamente do que outras espécies de Portia, incluindo a própria P. fimbriata em outras áreas.[11]
Se perturbada, a maioria das espécies de Portia salta para cima cerca de 100 a 150 mm, frequentemente a partir da postura de repouso críptica, e muitas vezes em uma ampla trajetória. Geralmente, a aranha então "congela" ou corre cerca de 100 mm e depois para. No entanto, P. fimbriata em Queensland raramente corre ou salta.[11]
Sentidos
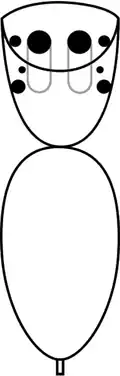
Embora outras aranhas também possam saltar, as espécies da família Salticidae, incluindo P. fimbriata, têm uma visão significativamente melhor que outras aranhas,[12][13] e seus olhos principais são mais aguçados à luz do dia do que os de um gato e 10 vezes mais do que os de uma libélula.[3] As aranhas saltadoras têm oito olhos, os dois grandes na posição central e frontal (os olhos médio-anteriores, também chamados de "olhos principais")[14] alojados em tubos no cefalotórax e que proporcionam visão acurada. Os outros seis olhos são secundários, posicionados ao longo das laterais da carapaça e que atuam principalmente como detectores de movimento.[3][15] Na maioria das aranhas saltadoras, o par médio de olhos secundários é muito pequeno e não tem função conhecida, mas os das espécies de Portia são relativamente grandes e funcionam tão bem quanto seus outros olhos secundários.[11][16] Os olhos principais focalizam com precisão um objeto a distâncias de aproximadamente 2 cm até o infinito,[14] e, na prática, podem enxergar até cerca de 75 cm.[14] Como todas as aranhas saltadoras, P. fimbriata pode captar apenas um pequeno campo visual de cada vez,[5] já que a parte mais acurada de um olho principal pode ver todo um círculo de até 12 mm de largura a 20 cm de distância, ou até 18 mm de largura a 30 cm de distância.[7][a]
Geralmente a subfamília de aranhas saltadoras Spartaeinae, que inclui o gênero Portia, não pode discriminar objetos a distâncias tão longas quanto os membros das subfamílias Salticinae ou Lyssomaninae. No entanto, os olhos principais de Portia têm uma visão quase tão aguçada quanto a das melhores aranhas saltadoras: a saltícide Mogrus neglectus pode distinguir presas e coespecíficos a até 320 mm de distância (42 vezes o comprimento de seu próprio corpo), enquanto P. fimbriata pode distinguir esses a até 280 mm (47 vezes o comprimento de seu próprio corpo).[17] Os olhos principais da espécie também podem identificar características do cenário até 85 vezes o comprimento de seu próprio corpo, o que ajuda a aranha a encontrar desvios.[6]
No entanto, uma aranha Portia leva um tempo relativamente longo para ver objetos, possivelmente porque obter uma boa imagem a partir de olhos tão pequenos é um processo complexo que requer muita varredura.[d] Isso torna a espécie vulnerável a predadores muito maiores, como aves, sapos e louva-a-deus, que Portia frequentemente não consegue identificar devido ao tamanho do outro predador.[3]
Aranhas, como outros artrópodes, possuem sensores, frequentemente "setas [en]" modificadas (como cerdas), para olfato, paladar, tato e vibração, que se projetam através de sua cutícula ("pele").[18] Aranhas do grupo Portia podem sentir as vibrações das superfícies e usá-las para acasalar e caçar outras aranhas na escuridão total. Ela pode usar os "cheiros" do ar e da superfície para detectar presas que encontra com frequência, identificar membros da mesma espécie, reconhecer membros familiares e determinar o sexo de outro membro da mesma espécie.[6]
Caça e alimentação
Táticas de caça do gênero Portia
Os membros do gênero Portia são chamados de "gatos de oito patas", pois suas táticas de caça são tão versáteis e adaptáveis quanto as de um leão.[3] Todos os membros do grupo têm táticas instintivas para suas presas mais comuns, mas podem improvisar por tentativa e erro contra presas desconhecidas ou em situações desconhecidas, e depois se lembrar da nova abordagem. Eles também podem fazer desvios para encontrar o melhor ângulo de ataque contra presas perigosas, mesmo quando o melhor desvio tira a aranha do contato visual direto com a presa,[3] e, às vezes, a rota planejada leva ao "rapel" por um fio de seda (teia) e à mordida da presa por trás. Esses desvios podem levar até uma hora,[19] e o indivíduo geralmente escolhe a melhor rota.[10]

Enquanto a maioria das aranhas saltadoras caça principalmente insetos por caça ativa,[20] as fêmeas de Portia também constroem teias para capturar presas diretamente.[3][19] Essas "teias de captura" têm formato de funil e são mais largas na parte superior[21][22] e têm cerca de 4.000 cm³ de volume.[11] Aranhas Portia geralmente constroem sua própria teia sobre uma teia de uma aranha não saltadora.[3] Quando não está unida à teia de outra aranha, a teia de captura de uma fêmea de P. fimbriata geralmente é suspensa em bases rígidas, como galhos e pedras.[11] Os machos de Portia não constroem teias de captura.[11]
Uma aranha do gênero Portia pode manipular a teia de outra aranha com uma gama praticamente ilimitada de sinais, seja para atrair a presa para a abertura da teia ou acalmar a presa repetindo monotonamente o mesmo sinal enquanto Portia se aproxima lentamente para mordê-la.[20] Essas táticas permitem que espécies do grupo capturem aranhas de teia, como Holocnemus pluchei[23] de 10% a 200% do seu tamanho,[3] e as espécies de Portia caçam em todos os tipos de teias.[22] Em contrapartida, outras aranhas cursoriais geralmente têm dificuldade de se movimentar em teias, e as aranhas construtoras de teias têm dificuldade de se movimentar em teias diferentes das que elas constroem. Ao caçar na teia de outra aranha, os movimentos lentos e descompassados da Portia e as abas de suas pernas fazem com que ela se pareça com detritos de folhas presos na teia e soprados pela brisa.[22] P. fimbriata e algumas outras espécies do gênero usam a brisa e outros distúrbios como "cortinas de fumaça" nas quais esses predadores podem se aproximar mais rapidamente das aranhas de teia e voltar a uma abordagem mais cautelosa quando o distúrbio desaparece.[24] Algumas aranhas de teia fogem para longe quando percebem o andar sem ritmo de uma Portia entrando na teia - uma reação que Wilcox e Jackson chamam de "pânico da Portia".[10]
Se um inseto grande fica preso em uma teia, Portia geralmente espera até um dia para que o inseto pare de lutar, mesmo que a presa esteja completamente presa na teia.[11] Quando um inseto está preso em uma teia pertencente às espécies P. labiata, P. schultzi ou qualquer variante regional de P. fimbriata, e ao lado da teia de uma aranha de teia, a aranha de teia às vezes entra na teia da Portia, e então a aranha Portia persegue e captura a aranha de teia invasora.[11] Além disso, as teias de aranhas nas quais espécies de Portia caçam às vezes contêm insetos mortos e outros artrópodes que estão intactos ou parcialmente consumidos. P. fimbriata (em Queensland) e algumas outras espécies do gênero, como P. labiata e P. schultzi, às vezes praticam necrofagia desses cadáveres se não estiverem obviamente decompostos.[11]
Ao usar sua própria teia para capturar outras aranhas saltadoras, P. fimbriata esconde seus palpos conspícuos, o que não faz quando persegue uma aranha de teia ou, ocasionalmente, uma mosca em movimento.[7] Todas as espécies de Portia comem ovos de outras aranhas, incluindo ovos de sua própria espécie e de outras aranhas cursoriais, e podem extrair ovos de invólucros que variam desde os frágeis de Pholcus até os resistentes e coriáceos de Philoponella. Enquanto apenas a espécie P. fimbriata (em Queensland) captura aranhas cursoriais em seus ninhos, todas as espécies de Portia roubam ovos de ninhos vazios de aranhas cursoriais.[11]
O veneno de Portia é excepcionalmente poderoso contra aranhas.[22] Quando uma aranha do gênero perfura uma aranha pequena a média (até o peso de Portia),[11] incluindo outra aranha do mesmo gênero, a presa geralmente foge por cerca de 100 a 200 mm, entra em convulsões, fica paralisada após 10 a 30 segundos e continua convulsionando por 10 segundos a 4 minutos. O indivíduo predador de Portia se aproxima lentamente da presa e a toma.[11] A predadora geralmente precisa infligir até 15 perfurações para imobilizar completamente uma aranha maior (1,5 a 2 vezes o seu peso), e então pode esperar a cerca de 20 a 200 mm de distância por 15 a 30 minutos após capturar a presa.[11] Insetos geralmente não são imobilizados tão rapidamente, mas continuam a lutar, às vezes por vários minutos.[11]
Ocasionalmente, uma aranha Portia é morta ou ferida enquanto persegue presas até duas vezes seu tamanho. Em testes, a espécie irmã P. labiata é morta em 2,1% das perseguições e ferida, mas não morta, em 3,9%, enquanto P. schultzi é morta em 1,7% e ferida, mas não morta, em 5,3%. Já a espécie P. fimbriata é morta em 0,06% de suas perseguições e ferida, mas não morta, em outros 0,06%. A pele especialmente resistente do gênero frequentemente previne ferimentos, mesmo quando seu corpo é capturado pelas presas de outra aranha. Quando ferida, Portia sangra e às vezes pode perder uma ou mais pernas. Os palpos e pernas das aranhas se desprendem facilmente quando atacados, os palpos e pernas de Portia se desprendem excepcionalmente fácil, o que pode ser um mecanismo de defesa, e espécies do grupo são frequentemente vistas com pernas ou palpos faltando, enquanto outras aranhas saltadoras no mesmo habitat não são vistas com pernas ou palpos faltando.[11] Um espécime de P. fimbriata, agora na coleção do Museu Australiano, regenerou um membro perdido cerca de 7 dias após a muda.[1]
Táticas de caça de P. fimbriata
Todas as estatísticas de desempenho resumem resultados de testes em laboratório, usando espécimes em cativeiro.[11] As táticas e o desempenho das fêmeas de P. fimbriata mostram diferenças regionais entre as populações de Queensland, território do norte da Austrália e Sri Lanka.[11] A tabela também inclui fêmeas de P. africana ao redor do lago Vitória, de P. schultzi em partes do Quênia e de P. labiata no Sri Lanka para comparação.[11]
| Presa | Desempenho | P. fimbriata (Q) |
P. fimbriata (NT) |
P. fimbriata (SL) |
P. africana | P. labiata | P. schultzi |
|---|---|---|---|---|---|---|---|
| Salticidae | Tendência a perseguir a presa | 87% | 50% | 94% | 77% | 63% | 58% |
| Eficiência na captura de presas | 93% | 10% | 45% | 29% | 40% | 36% | |
| Aranhas de teia | Tendência a perseguir a presa | 91% | 94% | 64% | 74% | 83% | 84% |
| Eficiência na captura de presas | 92% | 81% | 83% | 65% | 79% | 72% | |
| Insetos | Tendência a perseguir a presa | 27% | 30% | 43% | 48% | 35% | 52% |
| Eficiência na captura de presas | 41% | 83% | 78% | 67% | 71% | 69% | |
Notas sobre esta tabela:
| |||||||
Em todas as regiões, P. fimbriata fixa suas teias em superfícies sólidas, como rochas, troncos e galhos de árvores, enquanto algumas outras espécies de Portia geralmente fixam suas teias em caules e folhas flexíveis e nos galhos mais baixos das árvores.[11]
Um teste realizado em 2001 demonstrou que quatro espécies saltadoras tomam néctar, seja sugando o néctar livre da superfície das flores ou mordendo as flores com suas presas. As aranhas se alimentavam em ciclos de dois a quatro minutos, depois limpavam seus corpos e especialmente suas quelíceras, antes de outro ciclo. Uma parte mais formal do teste mostrou que 90 aranhas saltadoras juvenis, incluindo P. fimbriata, geralmente preferem sugar de papel absorvente embebido em uma solução de açúcar a 30% em vez de papel embebido em água destilada. Os autores sugerem que, na natureza, o néctar pode ser uma forma frequente e conveniente de obter alguns nutrientes, pois evitaria o trabalho, os riscos e os custos (como a produção de veneno) da predação. As aranhas saltadoras podem se beneficiar dos aminoácidos, lipídios, vitaminas e minerais normalmente encontrados no néctar.[25]
Táticas em Queensland
P. fimbriata de Queensland é a aranha saltadora araneofágica (comedora de aranhas) mais estudada.[26] Robinson (2010) afirmou que a espécie em Queensland possui as técnicas de captura de presas mais variadas de qualquer animal no mundo, exceto humanos e outros primatas.[1] Quando não usa sua própria teia, a espécie caça principalmente aranhas saltadoras de outros gêneros, geralmente usando contra elas a tática de "perseguição críptica".[26]
| P. fimbriata (Q) | Outras espécies de Portia | |
|---|---|---|
| Níveis de luz | Baixo | Alto |
| Abundância de aranhas de teia locais | Alta | Baixa |
| Predando aranhas de teia | Mais eficiente
Maior probabilidade de vibrar a teia Menor probabilidade de saltar |
Menos eficiente
Menor probabilidade de vibrar a teia Maior probabilidade de saltar |
| Abundância de saltícides locais | Alta | Baixa |
| Predação de saltícides cursoriais | Mais eficiente
Usa perseguição críptica Geralmente se aproxima |
Menos eficiente
Não usa perseguição críptica Nunca se aproxima |
| Perseguição e captura de insetos | Menos eficiente | Mais eficiente |
| Caminhada lenta e mecânica | Exagerada | Menos pronunciada |
| Quando levemente perturbada | Adota uma postura críptica | Dá grandes saltos |
| Durante o acasalamento | O cortejo usa menos vibrações
A fêmea raramente se torce para atacar e comer o macho |
O cortejo usa mais vibrações
A fêmea se torce e se lança e muitas vezes come o macho |
Notas sobre esta tabela:
| ||
Machos adultos de P. fimbriata estão menos propensos a perseguir e são menos eficientes em capturar presas do que fêmeas adultas, especialmente contra presas maiores. Os machos são bastante eficazes contra aranhas de teia pequenas e relutantes em enfrentar as grandes, embora as capturem em cerca de 50% das tentativas. Contra outras aranhas saltadoras, os machos não perseguem as grandes e perseguem cerca de 48% das pequenas, capturando 84% daquelas que perseguem.[11] Machos de Portia não constroem teias grandes para capturar presas.[11]
Um teste em 1997 mostrou que as preferências de P. fimbriata por diferentes tipos de presas estão na ordem: aranhas de teia; aranhas saltadoras; e insetos.[27] Essas preferências se aplicam a presas vivas e iscas imóveis, e a espécimes de P. fimbriata sem presas por 7 dias ("bem alimentados") e sem presas por 14 dias ("famintos").[27] Espécimes sem presas por 21 dias ("extremamente famintos") não mostraram preferência por diferentes tipos de presas. O teste incluiu como presas várias espécies de aranhas de teia e aranhas saltadoras, e a seleção das espécies de presas não mostrou evidências de afetar os resultados.[27] Nesse teste, os insetos foram representados pela mosca doméstica Musca domestica.
| Presa | P. fimbriata (Q) | Oustras espécies de Portia | |
|---|---|---|---|
| Salticidae | Mediana | 26 min | 3 min |
| Intervalo | 1-318 min | 0-41 min | |
| Aranha de teia | Mediana | 16 min | 5 min |
| Intervalo | 0-583 min | 0-465 min | |
| Inseto | Mediana | 3 min | 3 min |
| Intervalo | 0-34 min | 0-45 min |
Ao caçar a maioria das outras aranhas saltadoras em Queensland, P. fimbriata exagera a lentidão e o caráter arrítmico de seu andar normal (às vezes chamado de "robótico")[6] e mantém seus palpos retraídos ao lado de suas quelíceras, como faz na postura de repouso críptico. Se a presa enfrenta seu predador P. fimbriata, o mesmo "congela" até que a presa se vire.[9][28] Essa perseguição críptica parece ser exclusiva de Queensland, onde a maioria das outras aranhas saltadoras não reconhece uma P. fimbriata disfarçada como predadora,[9] ou mesmo como um animal.[11] A espécie usa a perseguição críptica tanto contra outras aranhas saltadoras nativas de Queensland quanto de fora.[29]
Outras saltícides frequentemente se defendem quando perseguidas por outras espécies de Portia ou por P. fimbriata fora de Queensland, e a perseguição críptica da P. fimbriata de Queensland pode ser uma adaptação regional às presas saltícides abundantes, mas perigosas, especialmente a espécie Jacksonoides queenslandicus, na floresta tropical local.[9][28] P. fimbriata usa a perseguição críptica até mesmo contra algumas saltícides de formato estranho, como do gênero Holoplatys e a espécie Mantisatta longicauda, alongada e parecida com um louva-a-deus.[29]
Euryattus, outro gênero de aranha saltadora de Queensland, tem uma área de distribuição parcialmente sobreposta à de P. fimbriata[10] e é abundante em sua área comum. Aranhas P. fimbriata adultas e juvenis grandes caçam Euryattus adotando táticas específicas. Diferentemente da maioria das aranhas saltadoras, Euryattus faz um ninho suspendendo uma folha morta enrolada por linhas de seda da vegetação. P. fimbriata captura fêmeas de Euryattus imitando as vibrações feitas por machos de Euryattus como parte de seu cortejo, e esse engano atrai as fêmeas para fora de seus ninhos.[30] Em testes, indivíduos de Euryattus da área de atuação da P. fimbriata reconhecem o predador e se defendem, enquanto os espécimes de Euryattus de fora da área de atuação da P. fimbriata raramente reconhecem a ameaça. A predadora P. fimbriata tem mais facilidade para capturar Euryattus de fora da área de vida do predador do que capturar a mesma espécie que compartilha a área de vida da P. fimbriata.[10] Esse pode ser um exemplo de uma corrida armamentista evolutiva.[31]

P. fimbriata não persegue todas as espécies do gênero de aranhas saltadoras que imitam formigas, Myrmarachne [en],[32] e usa perseguição críptica apenas cerca de 20% do tempo contra outras saltícides que imitam formigas e besouros.[29] P. fimbriata também pode não usar perseguição críptica contra fêmeas da subfamília saltícide Lyssomaninae. Essas fêmeas são excepcionalmente translúcidas, e a cutícula translúcida faz com que os olhos médio-anteriores (principais) mostrem regiões claras e escuras que piscam quando vistas de frente. Machos de Lyssomaninae não são translúcidos e não produzem esse piscar, e P. fimbriata usa perseguição críptica consistentemente contra os machos. Isso sugere que os olhos médio-anteriores piscantes das fêmeas de Lyssomaninae podem reduzir a capacidade de P. fimbriata de identificá-las como aranhas saltadoras.[9]
Ao encontrar J. queenslandicus, P. fimbriata frequentemente primeiro nota pistas químicas nas linhas de segurança de seda de J. queenslandicus e então procura sua presa. O cheiro ajuda P. fimbriata a perceber sua presa mais rapidamente,[6] possivelmente por meio da redução dos limiares no sistema visual.[6] Às vezes, P. fimbriata não consegue ver J. queenslandicus através da camuflagem da presa e "caça por especulação", saltando alto no ar, de modo que o indivíduo de J. queenslandicus se trai ao se virar e procurar a perturbação.[6][28] A predadora P. fimbriata então se vira em direção à presa e agita seus palpos.[33] Parece que apenas a espécie P. fimbriata de Queensland se comporta dessa forma, enquanto espécies de Portia de outras áreas não o fazem, que P. fimbriata de Queensland reage dessa forma apenas a J. queenslandicus e que J. queenslandicus não percebe nenhum aviso químico de que P. fimbriata está por perto.[28]
Ao perseguir qualquer não salticídea, P. fimbriata não usa perseguição críptica e não retrai consistentemente seus palpos, nem congela quando enfrentada pela presa.[9] P. fimbriata adota a perseguição críptica somente depois de reconhecer a presa como uma aranha saltadora.[7]
Em Queensland, P. fimbriata é relutante em pular nas teias de aranhas que são suas presas, enquanto outras espécies de Portia fazem isso em qualquer oportunidade.[22] A aranha de teia orbicular Argiope sacode sua teia violentamente para afastar intrusos, e P. fimbriata encontra um desvio que permite descer por rapel até a presa.[10] Quando a aranha de teia Zosis genicularis está ocupada embrulhando sua própria presa e está menos atenta a outros predadores, P. fimbriata usa essa atividade como uma espécie de "cortina de fumaça" para se aproximar da aranha de teia.[34]
P. fimbriata usa perseguição não críptica contra aranhas das famílias Lycosidae, Clubionidae, Theridiidae e Desidae, e contra moscas, mas não persegue besouros ou formigas.[29]
Diferentemente de outras espécies de Portia, P. fimbriata de Queensland invadem prontamente os ninhos de aranhas cursoriais [en], manipulando ou cortando o ninho. Se a aranha residente eventualmente deixa o ninho, P. fimbriata a persegue. Se a aranha residente tenta contra-atacar e depois recua para o ninho, P. fimbriata pode atacar a outra aranha quando ela reentra no ninho ou pode esperar imóvel até que a presa saia. Se uma aranha presa perfurada recua para o ninho, P. fimbriata de Queensland nunca entra no ninho, mas espera que a presa saia, e então P. fimbriata a mata.[11]
As aranhas foram divididas em aquelas que:
- Saltam e depois nadam
- Apenas recompensadas
- Apenas penalizadas
- Apenas nadam
- Apenas recompensadas
- Apenas penalizadas
Um teste em um ambiente artificialmente projetado explorou a capacidade de P. fimbriata de Queensland de resolver um problema novo por tentativa e erro. Uma pequena ilha foi montada no meio de um atol em miniatura, e o espaço entre eles foi preenchido com água. O espaço era largo demais para as aranhas saltarem diretamente, e as opções das aranhas eram saltar e depois nadar ou apenas nadar. Os testadores incentivaram alguns espécimes usando uma pequena concha para criar ondas em direção ao atol quando as aranhas escolhiam a opção preferida pelos testadores (saltar e depois nadar, ou apenas nadar), e desincentivaram alguns espécimes criando ondas de volta para a ilha quando as aranhas escolhiam a opção que os testadores não queriam – em outras palavras, os testadores "recompensaram" um grupo por comportamento "bem-sucedido" e "penalizaram" o outro grupo por comportamento "indesejado".[35] Os espécimes de P. fimbriata de Queensland geralmente repetiam o comportamento bem-sucedido e mudavam se a primeira tentativa fosse mal-sucedida, independentemente de qual opção (saltar e depois nadar ou apenas nadar) os testadores escolheram como "boa" para cada espécime.[36]
Táticas no território do norte
No território do norte da Austrália, P. fimbriata não possui táticas especiais contra outras aranhas saltadoras e as trata como se fossem aranhas de teia. Ela tenta pular sobre elas ou desiste,[1] portanto, essa variante é ruim na captura de outras aranhas saltadoras. A variante do território do norte da espécie não é tão boa quanto a variante de Queensland na captura de aranhas de teia, mas é melhor que a variante do Sri Lanka e algumas outras espécies de Portia. Ela não se entusiasma em perseguir insetos, mas é muito boa em capturar aqueles que persegue, como mostra a tabela de desempenho acima.[11] Enquanto as perseguições da variante de Queensland normalmente levam 26 minutos, as da variante do território do norte normalmente levam de 3 a 5 minutos, como algumas outras espécies do gênero.[11]
Táticas no Sri Lanka
A variante do Sri Lanka persegue entusiasticamente outras aranhas saltadoras e é ligeiramente melhor que a maioria das espécies de Portia em testes, mas cerca de metade tão eficaz quanto a variante de Queensland. No Sri Lanka, P. fimbriata não é uma caçadora prolífica de aranhas de teia ou insetos, mas captura aqueles que persegue com bastante eficiência.[11] Como outras espécies do gênero, P. fimbriata do Sri Lanka geralmente leva 3 a 5 minutos para uma perseguição.[11]
Reprodução e ciclo de vida

Antes do cortejo, o macho de Portia tece uma pequena teia entre galhos ou ramos, pendura-se sob ela e ejacula sobre a teia.[11] Em seguida, o macho coleta o sêmen em reservatórios nos bulbos palpares de seus pedipalpos.[8]
Um teste em laboratório demonstrou como os machos de P. fimbriata de Queensland minimizam o risco de encontrar outros machos, reconhecendo pedaços novos de papel absorvente, alguns contendo suas próprias linhas de arrasto de seda e outros com linhas de arrasto de outro macho. Os machos também foram atraídos por papel absorvente novo contendo linhas de arrasto de fêmeas, enquanto as fêmeas não reagiram a papel absorvente novo com linhas de arrasto de machos. Isso sugere que os machos geralmente procuram pelas fêmeas, e não o contrário. Nenhum dos sexos reagiu a papel absorvente com uma semana de idade, independentemente de conter linhas de arrasto de machos ou fêmeas. Uma série semelhante de testes mostrou que P. labiata apresentou os mesmos padrões de respostas entre os sexos.[37]
Quando encontra outro indivíduo da mesma espécie, P. fimbriata não utiliza perseguição, mas exibe movimentos rápidos e suaves, exibindo-se a uma distância de 4 a 27 centímetros. A aranha levanta as pernas, seu corpo balança de um lado para o outro, e os palpos são abaixados abaixo das quelíceras . Isso é muito diferente da perseguição que utiliza ao encontrar outra saltcide de espécie diferente, apesar de receber o mesmo estímulo visual, que é a visão dos grandes olhos médio-anteriores do outro. Embora P. fimbriata seja influenciada por feromônios muito mais do que o comum entre aranhas saltadoras, os sinais visuais sozinhos são suficientes para iniciar exibições e distinguir membros da mesma espécie de outras saltícides, mesmo que nenhum dos parceiros se mova. As pernas finas e franjadas das espécies de Portia podem identificar membros da mesma espécie, além de ocultar essas aranhas de outras espécies de saltícides.
Em P. fimbriata de Queensland e em algumas outras espécies, disputas entre machos geralmente duram apenas 5 a 10 segundos, com contato apenas entre as pernas.[11] Disputas entre fêmeas de Portia são geralmente longas e violentas,[22] e em P. fimbriata de Queensland frequentemente incluem agarramentos que podem quebrar uma perna.[11] A vencedora pode expulsar a perdedora, comer seus ovos e tomar posse de sua teia.[11][22] Diferentemente de algumas outras espécies de Portia, as fêmeas vitoriosas de P. fimbriata de Queensland não matam nem comem as perdedoras.[11]
Uma fêmea que avista um macho pode se aproximar lentamente ou esperar. O macho então caminha ereto, exibindo-se ao acenar suas pernas e palpos. Se a fêmea não fugir, ela realiza primeiro uma "exibição propulsiva".[c] Se o macho mantém sua posição e ela não foge ou repete a exibição propulsiva, ele se aproxima e, se ela for madura, eles copulam, com o macho inserindo a ponta de um de seus bulbos palpais na abertura copulatória da fêmea, usando o primeiro palpo que fez contato raspante.[11] Se a fêmea for subadulta (a uma muda da maturidade), um macho ou, às vezes, um macho subadulto de P. fimbriata pode coabitar na teia de captura da fêmea.[11] As espécies de Portia geralmente copulam em uma teia ou em uma linha de arrasto feita pela fêmea.[22] P. fimbriata geralmente copula por cerca de 100 segundos, enquanto outros gêneros podem levar vários minutos ou até várias horas.[11][22] Diferentemente de algumas outras espécies de Portia, as fêmeas de P. fimbriata de Queensland não comem seus parceiros durante o cortejo, nem durante[11] ou após a cópula.[38]
Quando caçam, fêmeas maduras de P. fimbriata, P. africana, P. labiata e P. schultzi emitem sinais olfativos que reduzem o risco de que outras fêmeas, machos ou juvenis da mesma espécie disputem a mesma presa. Esse efeito inibe o mimetismo agressivo contra uma presa aranha, mesmo que a presa esteja visível, ou se estiver habitando qualquer parte de uma teia. Se uma fêmea dessas espécies de Portia detecta o cheiro de um macho da mesma espécie, ela estimula os machos a cortejar. Essas espécies de Portia não exibem esse comportamento quando recebem sinais olfativos de membros de outras espécies de Portia.[39]
Em testes em laboratório, espécies de Portia, incluindo P. fimbriata, copulam com outras espécies, mas as fêmeas então não produzem ovos.[11] Em Queensland, P. fimbriata prefere botar ovos em folhas secas e marrons de cerca de 20 mm de comprimento, suspensas próximo ao topo de sua teia de captura, e depois cobri-los com uma camada de seda. Se não houver uma folha seca disponível, a fêmea constrói uma pequena plataforma horizontal de seda na teia de captura, bota os ovos sobre ela e depois cobre os ovos. No território do norte, P. fimbriata ocasionalmente bota ovos em uma folha seca, mas mais frequentemente em um saco de ovos de seda em uma pequena teia horizontal suspensa na teia principal.[11]
Como todos os artrópodes, as aranhas passam por mudas, e, após a eclosão, o estágio de vida antes de cada muda é chamado de "instar".[40] Exemplares de P. fimbriata atingem a maturidade no instar 7, 8 ou 9. Em um experimento com filhotes de P. fimbriata de Queensland, 64% daqueles alimentados apenas com aranhas sobreviveram até a maturidade, 37% dos alimentados com uma mistura de aranhas e insetos sobreviveram, e todos os alimentados exclusivamente com insetos morreram antes de alcançar o 6º instar.[41] Para a muda, todas as espécies de Portia tecem uma teia horizontal com diâmetro cerca de duas vezes o comprimento do corpo da aranha, suspensa a apenas 1 a 4 mm abaixo de uma folha. A aranha fica de cabeça para baixo e frequentemente desliza 20 a 30 mm durante a muda.[22] As espécies de Portia tecem uma teia temporária semelhante para descansar.[22] P. fimbriata em Queensland pode ser muito sedentária, em alguns casos permanecendo na mesma teia por mais de 48 dias durante uma série de mudas.[16]
Ecologia
P. fimbriata é encontrada nas florestas tropicais da Índia, Nepal, Sri Lanka, Hong Kong, Taiwan, Nova Guiné, Ilhas Salomão, Malásia, Indonésia e no território do norte e em Queensland, na Austrália.[4][42][11] [3,43] Vive em folhagens, troncos de árvores, pedregulhos e paredes de rocha.[5] Em toda a sua área de distribuição, essa é a espécie mais comum do gênero Portia.[43] Os espécimes de P. fimbriata de Queensland vivem perto de água corrente e onde há luz moderada, enquanto os espécimes do território do norte vivem em cavernas onde a luz varia de bastante escura na parte de trás a muito mais brilhante ao redor das bocas. Outras populações de Portia também vivem com níveis de luz mais altos do que em Queensland, e alguns membros dessas outras populações são encontrados em teias expostas à luz solar direta durante parte do dia.[11] Em Queensland, a espécie compartilha seu ambiente com uma presa comum, a muito abundante Jacksonoides queenslandicus,[9] e com grandes populações de outras aranhas construtoras de teias saltícides e não saltícides que não são Portia.[11]
As formigas atacam P. fimbriata, enquanto a aranha não persegue as formigas, considerando-as venenosas ou muito desagradáveis.[29] P. fimbriata também é predada por pássaros, sapos e louva-a-deus.[3] Muitas vezes é difícil encontrar essa espécie na natureza, pois sua forma e seus movimentos são bem disfarçados. A variedade de Queensland é bastante fácil de criar, enquanto a variedade do território do norte é bastante problemática de manter.[1]
Taxonomia
P. fimbriata é uma das 17 espécies do gênero Portia [maio de 2016].[44] Wanless dividiu o gênero em dois grupos de espécies: o grupo "schultzi", no qual os palpos dos machos têm uma apófise tibial fixa; e o grupo "kenti", no qual a apófise de cada palpo nos machos tem uma articulação separada por uma membrana.[3] O grupo schultzi inclui P. schultzi, P. africana, P. fimbriata e P. labiata.[3]
A espécie P. fimbriata foi originalmente descrita por Carl Ludwig Doleschall como Salticus fimbriata em 1859.[45] A espécie também foi denominada Attus fimbriatus (Doleschall, 1859), Sinis fimbriatus (Doleschall, 1859), Linus fimbriatus (Doleschall, 1859) e Boethoportia ocellata (Hogg, 1915), e finalmente Portia fimbriata (Doleschall, 1859),[46][4] sendo o último nome usado até a atualidade.[3] De acordo com Jackson e Hallas, P. fimbriata, conforme definido atualmente, provavelmente inclui duas ou mais espécies distintas. Em particular, a aranha P. fimbriata de Queensland é provavelmente uma espécie distinta da P. fimbriata do Sri Lanka, pois os acasalamentos entre os dois grupos são inférteis.[11]
Portia faz parte da subfamília Spartaeinae,[47] que se acredita ser primitiva.[22] A filogenia molecular, uma técnica que compara o DNA de organismos para reconstruir a árvore da vida, indica que Portia é um membro do clado Spartaeinae, que Spartaeinae é basal (bastante semelhante aos ancestrais de todas as aranhas saltadoras) e que os parentes mais próximos de Portia são os gêneros Spartaeus, Phaeacius e Holcolaetis.[48]
Notas
a: ↑ Jackson e Blest (1982) afirmam: “A resolução do mosaico de receptores da camada I na retina central foi estimada num ângulo visual de 2,4 minutos de arco, o que corresponde a 0-12 mm à 20 cm de frente da aranha, ou 0/18 mm à 30 cm."[7]
b: ↑ Várias espécies de aranhas cursoriais bebem néctar como suplemento ocasional da sua dieta, e os juvenis de algumas aranhas de teia orbital digerem pólen enquanto reciclam as suas teias.[49] Uma aranha saltadora (avaliação de 2010), Bagheera kiplingi, foi registrada como quase totalmente herbívora.[50]
c: ↑ As “manifestações propulsivas” são movimentos súbitos e rápidos que incluem golpes, cargas, golpes de aríete e saltos.[11]
d: ↑ A retina encontra-se na extremidade de um tubo. A extremidade interna do tubo move-se de um lado para o outro em um a dois ciclos por segundo, e gira 50° num ciclo que demora 10 segundos.[51]
Referências
- ↑ a b c d e f g Robinson, Martyn (2010). «Raising Portia fimbriata» (PDF). Invertebrate Rearing. 1 (1): 5–9. ISSN 2042-633X. Consultado em 24 de maio de 2011
- ↑ Wanless, F. R. (1978). «A revision of the spider genus Portia (Araneae: Salticidae)» (PDF). Bulletin of the British Museum (Natural History) Zoology. 34 (3). Consultado em 11 de agosto de 2011. Cópia arquivada (PDF) em 12 de agosto de 2011
- ↑ a b c d e f g h i j k l m n o Harland, D.P.; Jackson, R.R. (2000). «"Eight-legged cats" and how they see: a review of recent research on jumping spiders (Araneae: Salticidae)» (PDF). Cimbebasia. 16: 231–240. Consultado em 5 de maio de 2011. Cópia arquivada (PDF) em 28 de setembro de 2006
- ↑ a b c d e f Chang, Yung-Hau; I-Min Tso (2004). «Six newly recorded spiders of the genera Araneus, Larinia, Eriophora, Thanatus, Portia and Dolichognatha (Araneae: Araneidae, Philodromidae, Salticidae and Tetragnathidae) from Taiwan». Acta Arachnologica. 53 (1): 27–33. doi:10.2476/asjaa.53.27

- ↑ a b c Piper, Ross (2007). «The Quest for Food: Portia spider» (PDF). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. Westport, CT 06881: Greenwood Press. pp. 98–100. ISBN 978-0-313-33922-6. Consultado em 31 de março de 2011
- ↑ a b c d e f g h Harland, Duane P.; Robert R. Jackson (2004). «Portia Perceptions: The Umwelt of an Aranephagic Jumping Spider». In: Frederick R. Prete. Complex worlds from simpler nervous systems. [S.l.]: MIT Press. pp. 5–40. ISBN 978-0-262-66174-4. Consultado em 12 de abril de 2011
- ↑ a b c d e Jackson, R.R.; A.D. Blest (1982). «The distances at which a primitive jumping spider, Portia fimbriata, makes visual discriminations» (PDF). Journal of Experimental Biology. 97 (1): 441–445. doi:10.1242/jeb.97.1.441a. Consultado em 12 de maio de 2011
- ↑ a b Ruppert, E.E., Fox, R.S., and Barnes, R.D. (2004). «Chelicerata: Araneae». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. pp. 571–584. ISBN 978-0-03-025982-1
- ↑ a b c d e f g h Wilcox, R. Stimson; Robert R. Jackson (1998). «Cognitive Abilities of Araneophagic Jumping Spiders». In: Russell P. Balda; Irene Maxine Pepperberg; Alan C. Kamil. Animal cognition in nature: the convergence of psychology and biology in laboratory and field. [S.l.]: Academic Press. ISBN 978-0-12-077030-4
- ↑ a b c d e f Wilcox, R. Stimson; Robert R. Jackson (1998). «Cognitive Abilities of Araneophagic Jumping Spiders». In: Russell P. Balda; Irene Maxine Pepperberg; Alan C. Kamil. Animal cognition in nature: the convergence of psychology and biology in laboratory and field. [S.l.]: Academic Press. ISBN 978-0-12-077030-4
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay Jackson, Robert R.; Susan E. A. Hallas (1986). «Comparative biology of jumping spiders Portia africana, P. albimana, P. fimbriata, P. labiata and P. schultzi, areanophagic, web-building jumping spiders (Araneae: Salticidae) utilisation of webs, predatory versatility, and intraspecfic interactions». New Zealand Journal of Zoology. 13 (4): 423–489. ISSN 0301-4223. doi:10.1080/03014223.1986.10422978. Consultado em 17 de maio de 2011
- ↑ Jarman, Elizabeth A.R.; Robert R. Jackson (1986). «The biology of Taieria erebus (Araneae, Gnaphosidae), an araneophagic spider from New New Zealand: silk utilisation and predatory versatility». New Zealand Journal of Zoology. 13 (4): 521–540. ISSN 0301-4223. doi:10.1080/03014223.1986.10422980. Consultado em 1 de abril de 2011
- ↑ Hoefler, Chad D.; Andy Chen; Elizabeth M. Jakob (2006). «The Potential of a Jumping Spider, Phidippus clarus, as a Biocontrol Agent» (PDF). J. Econ. Entomol. 99 (2): 432–436. ISSN 0022-0493. PMID 16686143. doi:10.1603/0022-0493-99.2.432. Consultado em 25 de abril de 2011. Cópia arquivada (PDF) em 14 de junho de 2010
- ↑ a b c Forster, Lyndsay M. (1977). «A qualitative analysis of hunting behaviour in jumping spiders (Araneae: Salticidae)». New Zealand Journal of Zoology. 4: 51–62. doi:10.1080/03014223.1977.9517936
- ↑ Hill, David Edwin (2010). «Use of location (relative direction and distance) information by jumping spiders (Araneae, Salticidae, Phidippus) during movement toward prey and other sighted objectives» (PDF). Peckhamia. 83 (1): 1–103. ISSN 1944-8120. Consultado em 12 de abril de 2011
- ↑ a b Jackson, Robert R. (1986). «Web Building, Predatory Versatility, and the Evolution of the Salticidae». In: William A. Shear. Spiders – webs, behavior, and evolution. [S.l.]: Stanford University Press. ISBN 978-0-8047-1203-3. Consultado em 22 de maio de 2011
- ↑ Harland, Duane P.; Robert R. Jackson; Aynsley M. Macnab (1999). «Distances at which jumping spiders (Araneae: Salticidae) distinguish between prey and conspecific rivals». Journal of Zoology. 247 (3): 357–364. doi:10.1111/j.1469-7998.1999.tb00998.x
- ↑ Ruppert, E.E., Fox, R.S., and Barnes, R.D. (2004). «Arthropoda: Sense organs». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. pp. 532–537. ISBN 978-0-03-025982-1
- ↑ a b Wilcox, S.; Jackson, R. (2002). «Jumping Spider Tricksters» (PDF). In: Bekoff, M.; Allen, C.; Burghardt, G.M. The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition. [S.l.]: MIT Press. pp. 27–34. ISBN 978-0-262-52322-6. Consultado em 12 de maio de 2011. Cópia arquivada (PDF) em 21 de julho de 2011
- ↑ a b Jackson, Robert R.; Simon D. Pollard (1997). «Jumping spiders mating strategies: sex among cannibals in and out of webs». In: Jae C. Choe; Bernard J. Crespi. The Evolution of Mating Systems in Insects and Arachnids. [S.l.]: Cambridge University Press. pp. 340–351. ISBN 978-0-521-58976-5. Consultado em 6 de junho de 2011
- ↑ Richman, David B.; Robert R. Jackson (1992). «A review of the ethology of jumping spiders (Araneae, Salticidae)» (PDF). Bulletin of the British Arachnology Society. 9 (2): 33–37. Consultado em 12 de maio de 2011
- ↑ a b c d e f g h i j k l Jackson, Robert R.; Susan E.A. Hallas (1986). «Predatory versatility and intraspecies interactions of spartaeine jumping spiders (Araneae, Salticidae): Brettus adonis, B. cingulata, Cyrba algerina, and Phaeacius sp. indet». New Zealand Journal of Zoology. 13 (4): 491–520. ISSN 0301-4223. doi:10.1080/03014223.1986.10422979. Consultado em 1 de abril de 2011
- ↑ Jackson, R. R.; Jakob, E. M.; Willey, M. B.; Campbell, G. E. (1993). «Anti‐predator defences of a web‐building spider, Holocnemus pluchei (Araneae, Pholcidae)». Journal of Zoology (em inglês) (2): 347–352. ISSN 0952-8369. doi:10.1111/j.1469-7998.1993.tb02641.x. Consultado em 4 de junho de 2025
- ↑ Wilcox, R. Stimson; Robert R. Jackson; Kristen Gentile (1996). «Spiderweb smokescreens: spider trickster uses background noise to mask stalking movements» (PDF). Animal Behaviour. 51 (2): 313–326. CiteSeerX 10.1.1.583.2268. ISSN 0003-3472. doi:10.1006/anbe.1996.0031. Consultado em 23 de maio de 2011
- ↑ Jackson, Robert R.; Simon D. Pollard; Ximena J. Nelson; G. B. Edwards; Alberto T. Barrion (2001). «Jumping spiders (Araneae: Salticidae) that feed on nectar» (PDF). Journal of Zoology. 255: 25–29. doi:10.1017/s095283690100108x. Consultado em 18 de junho de 2011
- ↑ a b Harland, Duane P.; Robert R. Jackson (2002). «Influence of cues from the anterior medial eyes of virtual prey on Portia fimbriata, an araneophagic jumping spider». The Journal of Experimental Biology. 205 (13): 1861–1868. PMID 12077162. doi:10.1242/jeb.205.13.1861. Consultado em 22 de maio de 2011
- ↑ a b c Li, Daiqin; Robert R. Jackson; Alberto Barrion (1997). «Prey preferences of Portia labiata, P. africana, and P. schultzi, araneophagic jumping spiders (Araneae: Salticidae) from the Philippines, Sri Lanka, Kenya, and Uganda». New Zealand Journal of Zoology. 24 (4): 333–349. doi:10.1080/03014223.1997.9518129
- ↑ a b c d Jackson, Robert R.; Robert J. Clark; Duane P. Harland (2002). «Behavioural and cognitive influences of kairomones on an araneophagic jumping spider» (PDF). Behaviour. 139 (6): 749–775. doi:10.1163/156853902320262808. Consultado em 17 de maio de 2011. Cópia arquivada (PDF) em 27 de setembro de 2011
- ↑ a b c d e Harland, Duane P.; Robert R. Jackson (2001). «Prey classification by Portia fimbriata, a salticid spider that specializes at preying on other salticids: species that elicit cryptic stalking» (PDF). Journal of Zoology. 255 (4): 445–460. doi:10.1017/s0952836901001534. Consultado em 19 de maio de 2011. Cópia arquivada (PDF) em 27 de setembro de 2011
- ↑ Nelson, Ximena J.; Robert R. Jackson; Godfrey Sune (2005). «Use of Anopheles-specific prey-capture behavior by the small juveniles of Evarcha culicivora, a mosquito-eating jumping spider» (PDF). The Journal of Arachnology. 33 (2): 541–548. doi:10.1636/05-3.1. Consultado em 20 de junho de 2011. Cópia arquivada (PDF) em 16 de agosto de 2011
- ↑ Jackson, Robert R.; R. Stimson Wilcox (1990). «Aggressive mimicry, prey-specific predatory behaviour and predator-recognition in the predator-prey interactions of Portia fimbriata and Euryattus sp., jumping spiders from Queensland». Behavioral Ecology and Sociobiology. 26 (2): 111–119. doi:10.1007/BF00171580
- ↑ Harland, Duane P.; Robert R. Jackson (2001). «Prey classification by Portia fimbriata, a salticid spider that specializes at preying on other salticids: species that elicit cryptic stalking» (PDF). Journal of Zoology. 255 (4): 445–460. doi:10.1017/s0952836901001534. Consultado em 25 de maio de 2011. Cópia arquivada (PDF) em 27 de setembro de 2011
- ↑ Clark, Robert J.; Duane P. Harland; Robert J. Jackson (2000). «Speculative hunting by an araneophagic salticid spider». Behaviour. 137 (12): 1601–1612. JSTOR 4535796. doi:10.1163/156853900502736
- ↑ Jackson, Robert R.; Simon D. Pollard; Ana M. Cerveira (2002). «Opportunistic use of cognitive smokescreens by araneophagic jumping spiders». Animal Cognition. 5 (3): 147–157. ISSN 1435-9456. PMID 12357287. doi:10.1007/s10071-002-0144-9
- ↑ Jackson, Robert R.; Fiona R. Cross; Chris M. Carter (2006). «Geographic Variation in a Spider's Ability to Solve a Confinement Problem by Trial and Error». International Journal of Comparative Psychology. 19 (3): 282–296. doi:10.46867/IJCP.2006.19.03.06. Consultado em 8 de junho de 2011
- ↑ Jackson, Robert R.; Chris M. Carter; Michael S. Tarsitano (2001). «Trial-and-error solving of a confinement problem by a jumping spider, Portia fimbriata». Behaviour. 138 (10): 1215–1234. ISSN 0005-7959. JSTOR 4535886. doi:10.1163/15685390152822184
- ↑ Clark, R.J.; R.R. Jackson (1995). «Dragline-mediated sex recognition in two species of jumping spiders (Araneae Salticidae), Portia labiata and P. fimbriata». Ethology Ecology & Evolution. 7: 73–77. doi:10.1080/08927014.1995.9522970. Consultado em 13 de junho de 2011
- ↑ Jackson, Andrea . Phil Myers, ed. «ADW: Portia fimbriata: Information». Animal Diversity Web. University of Michigan Museum of Zoology. Consultado em 13 de maio de 2011
- ↑ Willey, Marianne B.; Robert R. Jackson (1993). «Olfactory cues from conspecifics inhibit the web-invasion behavior of Portia, web-invading araneophagic jumping spiders (Araneae: Salticidae)». Canadian Journal of Zoology. 71 (7): 1415–1420. doi:10.1139/z93-195
- ↑ Ruppert, E.E., Fox, R.S., and Barnes, R.D. (2004). «Arthropoda: Body wall». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. pp. 521–525. ISBN 978-0-03-025982-1
- ↑ Li, Daiqin; Jackson, Robert R. (1997). «Influence of diet on survivorship and growth in Portia fimbriata , an araneophagic jumping spider (Araneae: Salticidae)». Canadian Journal of Zoology (em inglês) (10): 1652–1658. ISSN 0008-4301. doi:10.1139/z97-792. Consultado em 5 de junho de 2025
- ↑ Žabka, Marek M.; Wolfgang Nentwig (2000). «Salticidae (Arachnida: Araneae) of the Krakatau Islands (Indonesia): a preliminary approach» (PDF). Ekológia (Bratislava). 19 (suppl. 3): 293–306. Consultado em 25 de maio de 2011
- ↑ Logunov, Dmitry V.; Galina N. Azarkina (2007). «New species and new records for jumping spiders of the subfamily Spartaeinae (Aranei: Salticidae)» (PDF). Arthropoda Selecta. 16 (2): 97–114. Consultado em 23 de maio de 2011. Cópia arquivada (PDF) em 24 de agosto de 2011
- ↑ «Gen. Portia Karsch, 1878». World Spider Catalog. Natural History Museum Bern. Consultado em 13 de maio de 2016
- ↑ Doleschall, C. Ludwig (1859). «Tweede Bijdrage tot de kennis der Arachniden van den Indischen Archipel». Acta Societatis Scientiarum Indo-Neêrlandicae. 5 (5): 1–60
- ↑ «Taxon details Portia fimbriata Doleschall, 1859». World Spider Catalog. Natural History Museum Bern. Consultado em 13 de maio de 2016
- ↑ Barrion, A.T.; J.A. Litsinger (1995). Riceland spiders of South and Southeast Asia. [S.l.]: International Rice Research Institute / C.A.B. International. p. 45. ISBN 978-0-85198-967-9
- ↑ Maddison, Wayne P.; Melissa R. Bodner; Karen M. Needham (2008). «Salticid spider phylogeny revisited, with the discovery of a large Australasian clade (Araneae: Salticidae)» (PDF). Zootaxa. 1893: 49–64. ISSN 1175-5334. doi:10.11646/zootaxa.1893.1.3. Consultado em 14 de junho de 2011
- ↑ Chen, Xiaoqiong; Yuanchun Chen; Lingbing Wu; Yu Peng; Jian Chen; Fengxiang Liu (2010). «A survey of nectar feeding by spiders in three different habitats» (PDF). Bulletin of Insectology. 63 (2): 203–208. ISSN 1721-8861. Consultado em 20 de maio de 2011. Cópia arquivada (PDF) em 22 de julho de 2011
- ↑ Meehan, Christopher J.; Eric J. Olson; Matthew W. Reudink; T. Kurt Kyser; Robert L. Curry (2009). «Herbivory in a spider through exploitation of an ant–plant mutualism». Current Biology. 19 (19): R892–R893. PMID 19825348. doi:10.1016/j.cub.2009.08.049
- ↑ Land, Michael F.; Dan-Eric Nilsson (2006). «General-purpose and special-purpose visual systems» (PDF). In: Eric Warrant; Dan-Eric Nilsson. Invertebrate vision. [S.l.]: Cambridge University Press. pp. 167–210. ISBN 978-0-521-83088-1. Consultado em 21 de junho de 2011
Ligações externas
- Vídeos do HowStuffWorks "Enganados pela natureza: Aranha saltadora australiana" - Portia fimbriata captura uma aranha de teia
- Salticidae: Biblioteca de desenhos diagnósticos - Portia fimbriata por Jerzy Proszynski