Portia schultzi
Portia schultzi
| |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||||||||||||
| Classificação científica | |||||||||||||||||||||
| |||||||||||||||||||||
| Nome binomial | |||||||||||||||||||||
| Portia schultzi Karsch, 1878[1] | |||||||||||||||||||||
| Sinónimos[1] | |||||||||||||||||||||
| |||||||||||||||||||||
Portia schultzi é uma espécie da família Salticidae que ocorre desde a África do Sul, no sul, até o Quênia, no norte, além de ser encontrada na África Ocidental e em Madagascar. Essa espécie é ligeiramente menor que outras do gênero Portia. As fêmeas têm corpos de 5 a 7 mm de comprimento, enquanto os machos têm de 4 a 6 mm. A carapaça de ambos os sexos é laranja-acastanhada com manchas marrons escuras, coberta por pelos marrons escuros e esbranquiçados. Os machos possuem tufos brancos no tórax e uma faixa branca larga acima da base das pernas, características menos evidentes nas fêmeas. Ambos os sexos têm tufos de laranja a laranja-escuro acima dos olhos, com bordas de pelos laranja-claros. Os abdômenes dos machos são amarelo-laranja a laranja-acastanhado com manchas escuras, e na parte superior apresentam pelos pretos e laranja-claros, além de nove tufos brancos. Já os das fêmeas são amarelo-claros com marcas pretas e pelos esparsos brancos e laranja-acastanhados na parte superior. A P. schultzi tem pernas relativamente mais longas do que outras Portia.[2]
Enquanto a maioria das aranhas da família Salticidae foca com precisão até cerca de 75 cm de distância, a P. schultzi responde a alvos a no máximo 10 cm em boa iluminação, ignorando tudo em luz muito fraca. Para presas, prefere aranhas que vivem em teias, seguidas por outras aranhas da família Salticidae e, por último, insetos. As fêmeas de P. schultzi e outras espécies de Portia constroem "teias de captura" para apanhar presas, frequentemente conectando suas teias às de aranhas que vivem em teias para capturar essas aranhas ou suas presas.
Se uma fêmea de P. schultzi estiver madura, um macho tentará copular com ela ou coabitar com uma fêmea subadulta e copular durante a ecdise. Eles geralmente copulam em uma teia ou em um fio de arrasto feito pela fêmea, com a cópula de P. schultzi durando cerca de 100 segundos, enquanto outras espécies do gênero podem levar minutos ou até horas. As fêmeas tentam matar e comer seus parceiros durante ou após a cópula, e fêmeas subadultas imitam adultas para atrair machos como presas. Confrontos entre fêmeas de Portia são violentos, e embates em P. schultzi duram de 20 a 60 segundos. Às vezes, uma fêmea derruba a outra de costas, e a derrotada pode ser morta e comida se não se recuperar rapidamente e fugir. Durante a caça, fêmeas maduras de P. schultzi emitem sinais olfativos que reduzem o risco de competição por presas com outras fêmeas, machos ou filhotes da mesma espécie.
Estrutura e aparência do corpo
O corpo das fêmeas de P. schultzi mede 5 a 7 mm (menor que outras espécies de Portia), enquanto o dos machos mede 4 a 6 mm.[3]:433 A carapaça de ambos os sexos é laranja-acastanhada com manchas marrons escuras, coberta por pelos marrons escuros e esbranquiçados. Os machos têm tufos brancos no tórax e uma faixa branca larga acima da base das pernas, menos evidentes nas fêmeas. Ambos possuem tufos de laranja a laranja-escuro acima dos olhos, com bordas de pelos laranja-claros. As quelíceras das fêmeas são amarelo-claras com marcas pretas nas extremidades, enquanto as dos machos são laranja-acastanhadas com marcas escuras, ambas com pelos laranja-claros e brancos. O abdômen das fêmeas é amarelo-claro com marcas pretas e pelos esparsos brancos e laranja-acastanhados na parte superior, sem tufos. Nos machos, é amarelo-laranja a laranja-acastanhado com manchas escuras, com pelos pretos e laranja-claros e nove tufos brancos na parte superior.[4]:88-89 As pernas de ambos os sexos são excepcionalmente longas e finas,[5]:34 sendo laranja-acastanhadas com marcas escuras nos machos e amarelo-claras com marcas escuras nas fêmeas.[4]:88-89 Os dois últimos segmentos de cada perna não possuem outras decorações,[5]:34 mas os outros segmentos têm pelos acastanhados e espinhos robustos, com tufos brancos esparsos nos machos. Os palpos de ambos os sexos têm pelos amarelo-claros e bordas brancas.[4]:88-89 Todas as espécies do gênero Portia têm abdômenes elásticos, que podem ficar quase esféricos quando bem alimentados, e os das fêmeas podem se esticar durante a produção de ovos.[6]:495
Movimento
Quando não está caçando presas ou parceiros para acasalar, as espécies de Portia, incluindo P. schultzi,[5]:31 adotam uma postura chamada "postura de repouso críptica", recolhendo as pernas próximas ao corpo e os palpos ao lado das quelíceras, o que oculta seus apêndices. Ao caminhar, a maioria das espécies de Portia tem um andar lento que mantém sua camuflagem: pausando frequentemente em intervalos irregulares, movendo as pernas continuamente e os palpos de forma brusca para cima e para baixo, cada apêndice fora de sincronia com os outros,[7][8]:6 e variando continuamente a velocidade e o tempo.[9]:418 A P. schultzi usa um andar descrito por Forster e Murphy (1986) como "saltitante",[5]:34-35[2] flexionando e esticando as pernas. Eles sugerem que as pernas longas são vantajosas para se mover por teias, e o andar saltitante permite que a P. schultzi use as pernas longas mantendo o corpo próximo à superfície.[5]:34
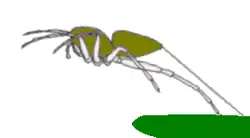
Como muitas aranhas, a P. schultzi produz um fio de arrasto contínuo de seda enquanto se move, ancorando-o ocasionalmente a uma superfície com um ponto de seda pegajosa. Isso permite que ela retorne à superfície se for desalojada. Antes de saltar, ela fixa um ponto de seda pegajosa e desenrola um fio de arrasto durante o salto.[10]:574 Diferentemente da maioria das aranhas saltadoras, os fios de arrasto da P. schultzi aderem uns aos outros, e, após traçar algumas linhas sobre um espaço, ela as usa como passagens, reforçando-as com mais seda enquanto se move.[5]:34
Quando perturbada, a maioria salta cerca de 100 a 150 mm para cima, muitas vezes a partir da postura de repouso críptica, geralmente em uma trajetória ampla. Normalmente, a Portia então congela ou corre cerca de 100 mm e congela novamente.[3]:434
Sentidos
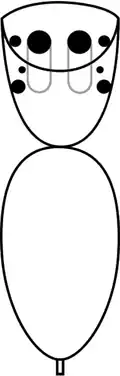
Embora outras aranhas também possam saltar, as saltadoras, incluindo a P. schultzi, têm uma visão significativamente melhor que outras aranhas,[11]:521[12] com olhos principais mais aguçados à luz do dia que os de um gato e dez vezes mais que os de uma libélula.[13] As aranhas saltadoras possuem oito olhos, com os dois grandes na posição central-frontal (olhos anteromedianos, ou "olhos principais")[14]:51 alojados em tubos na cabeça, proporcionando visão aguçada. Os outros seis são olhos secundários, posicionados nas laterais da carapaça, funcionando principalmente como detectores de movimento.[13][15]:16 Na maioria das aranhas saltadoras, o par médio de olhos secundários é muito pequeno e sem função conhecida, mas os da Portia são relativamente grandes e funcionam tão bem quanto os outros olhos secundários.[3]:424[16]:232 Os olhos principais das aranhas saltadoras enxergam do vermelho ao ultravioleta.[17]
Na maioria das aranhas saltadoras, os olhos principais focam com precisão objetos a até cerca de 75 cm de distância.[14]:51, 53 No entanto, a P. schultzi não reage a objetos quando a luz está abaixo de 100 lux. Entre 100 e 500 lux, ela detecta e se aproxima de objetos a até 6 cm; de 500 a 1500 lux, a distância de resposta aumenta gradualmente até cerca de 10 cm, sem aumento em luz mais forte. Para comparação, a Trite auricoma gira em direção a movimentos a até 75 cm e se aproxima de alvos a partir de 20 cm. Talvez a P. schultzi ganhe pouco ao ser alertada sobre objetos distantes, pois se move tão lentamente que é improvável alcançar um alvo mais distante a tempo.[5]:37
Como todas as aranhas saltadoras, a P. schultzi só consegue captar um pequeno campo visual de cada vez,[18] já que a parte mais aguçada de um olho principal pode ver um círculo de até 12 mm de largura a 20 cm de distância, ou até 18 mm a 30 cm.[19][a] A Portia leva um tempo relativamente longo para enxergar objetos, possivelmente porque obter uma boa imagem com olhos tão pequenos é um processo complexo que exige muita varredura.[b] Isso torna a Portia vulnerável a predadores maiores, como aves, sapos e louva-a-deus, que ela frequentemente não consegue identificar devido ao tamanho desses predadores.[13]
Aranhas, como outros artrópodes, possuem sensores, muitas vezes cerdas modificadas, para olfato, paladar, tato e vibração, que se projetam através da cutícula ("pele").[21]:532-533 Diferentemente dos insetos, aranhas e outros quelicerados não têm antenas.[22][23] A Portia pode sentir vibrações de superfícies, usando-as para acasalamento e caça de outras aranhas na escuridão total. Ela também usa "cheiros" do ar e de superfícies para detectar presas, identificar membros da mesma espécie, reconhecer indivíduos familiares e determinar o sexo de outros membros da mesma espécie.[8]:13
Táticas de caça
Táticas usadas pela maioria das aranhas saltadoras e pelo gênero Portia
As espécies do gênero Portia têm táticas de caça tão versáteis e adaptáveis quanto as de um leão.[13] Todas as Portia possuem táticas instintivas para suas presas mais comuns, mas podem improvisar por tentativa e erro contra presas desconhecidas ou em situações novas, memorizando a nova abordagem. Elas também fazem desvios para encontrar o melhor ângulo de ataque contra presas perigosas, mesmo que o melhor desvio as tire do contato visual com a presa,[13] e às vezes o trajeto planejado envolve "rapel" por um fio de seda para morder a presa por trás. Esses desvios podem levar até uma hora,[24] e a Portia geralmente escolhe a melhor rota, mesmo que precise passar por uma incorreta.[9]:422 Se cometer um erro ao caçar outra aranha, a própria Portia pode ser morta.[24]
Enquanto a maioria das aranhas saltadoras caça principalmente insetos por caça ativa,[25]:340 as fêmeas de Portia também constroem teias para capturar presas diretamente.[13][24]:513 Essas "teias de captura" são em forma de funil, mais largas no topo[17][6]:429-431 e têm cerca de 4.000 cm³ de volume.[3] A teia é inicialmente construída em cerca de 2 horas e gradualmente reforçada.[16]:239 A Portia frequentemente conecta sua teia à de uma aranha que vive em teias.[13] Quando não conectada a outra teia, a teia de captura de uma fêmea de P. schultzi pode ser suspensa em fundações rígidas, como galhos e rochas, ou em bases flexíveis, como caules de arbustos.[3]:432 Machos de Portia não constroem teias de captura.[3]:429
As aranhas Portia caçam em todos os tipos de teias,[6]:491 enquanto outras aranhas cursoras geralmente têm dificuldade em se mover em teias, e aranhas tecelãs têm problemas em teias diferentes das que constroem.[3]:424 Onde a teia é esparsa, a Portia usa "sondagem rotativa", movendo uma perna livre até encontrar um fio.[3]:433-434 Ao caçar na teia de outra aranha, os movimentos lentos da Portia e as abas em suas pernas fazem-na parecer detritos foliares presos na teia, movidos pela brisa.[6]:514 A P. schultzi e algumas outras Portia usam brisas e outras perturbações como "cortinas de fumaça" para se aproximar mais rapidamente de aranhas em teias, revertendo para uma abordagem cautelosa quando a perturbação cessa.[26]:313 Algumas aranhas de teia fogem ao sentir o andar arrítmico de uma Portia entrando na teia, uma reação chamada "pânico de Portia" por Wilcox e Jackson.[9]:418
Se um inseto grande está preso em uma teia, a Portia geralmente não o pega, esperando até um dia até que ele pare de lutar, mesmo que esteja bem preso.[3]:448 Quando um inseto está preso em uma teia de P. schultzi, P. labiata ou qualquer variante regional de P. fimbriata, ao lado da teia de uma aranha tecelã, a aranha tecelã às vezes entra na teia da Portia, e a Portia a persegue e captura.[3]:440-441, 444
As teias de aranhas nas quais as Portia caçam às vezes contêm insetos mortos e outros artrópodes não consumidos ou parcialmente consumidos. A P. schultzi e algumas outras Portia, como P. fimbriata (em Queensland) e P. labiata, às vezes se alimentam desses corpos se não estiverem visivelmente deteriorados.[3]:448
A Portia geralmente leva de 3 a 5 minutos para perseguir uma presa, mas algumas perseguições podem ser muito mais longas, em casos extremos até 10 horas ao perseguir uma aranha de teia.[3]:439
Todas as Portia comem ovos de outras aranhas, incluindo os de sua própria espécie e de outras aranhas cursoras, e podem extrair ovos de casulos, desde os frágeis de Pholcus até os resistentes de Philoponella. Embora apenas a P. fimbriata (em Queensland) capture aranhas cursoras em seus ninhos, todas as Portia roubam ovos de ninhos vazios de aranhas cursoras.[3]:448
O veneno da Portia é excepcionalmente poderoso contra aranhas.[6]:491 Quando uma Portia ataca uma aranha pequena a média (até seu próprio peso),[3]:428 a presa geralmente foge por 100 a 200 mm, entra em convulsões, fica paralisada após 10 a 30 segundos e continua convulsionando por 10 segundos a 4 minutos. A Portia se aproxima lentamente e a captura.[3]:441-443 Para imobilizar completamente uma aranha maior (1,5 a 2 vezes seu peso),[3]:428 a Portia pode precisar de até 15 picadas, podendo esperar a 20 a 200 mm de distância por 15 a 30 minutos após a captura.[3]:441-443 Insetos geralmente não são imobilizados tão rapidamente, continuando a lutar, às vezes por vários minutos. Se a Portia não consegue mais contato, todos os tipos de presas geralmente se recuperam, fazendo movimentos lentos após alguns minutos, mas voltando ao movimento normal após uma hora.[3]:441-443
Ocasionalmente, uma Portia é morta ou ferida ao perseguir presas até duas vezes seu tamanho. Em testes, a P. schultzi é morta em 1,7% das perseguições e ferida, mas não morta, em 5,3%, enquanto a P. labiata é morta em 2,1% e ferida em 3,9%, e a P. fimbriata em Queensland é morta em 0,06% e ferida em outros 0,06%. A pele especialmente resistente da Portia frequentemente previne ferimentos, mesmo quando seu corpo é preso nas presas de outra aranha. Quando ferida, a Portia sangra e pode perder uma ou mais pernas. Os palpos e pernas das aranhas se quebram facilmente quando atacados, os da Portia ainda mais, o que pode ser um mecanismo de defesa, e é comum ver Portia com pernas ou palpos faltando, enquanto outras saltadoras no mesmo habitat não apresentam essas perdas.[3]:440-450
Táticas usadas por Portia schultzi
Todas as estatísticas de desempenho resumem resultados de testes em laboratório com espécimes em cativeiro.[3]:429-430 A tabela a seguir mostra o desempenho de caça de fêmeas adultas. Além da P. schultzi, a tabela compara o desempenho de caça de P. africana, P. labiata e três variantes regionais de P. fimbriata.[3]:424, 432, 434
| Presa | Desempenho | P. schultzi | P. africana | P. labiata | P. fimbriata
(Q) |
P. fimbriata
(TN) |
P. fimbriata
(SL) |
|---|---|---|---|---|---|---|---|
| Salticídeos | Tendência a perseguir a presa | 58% | 77% | 63% | 87% | 50% | 94% |
| Eficiência na captura da presa | 36% | 29% | 40% | 93% | 10% | 45% | |
| Aranhas construtoras | Tendência a perseguir a presa | 84% | 74% | 83% | 91% | 94% | 64% |
| Eficiência na captura da presa | 72% | 65% | 79% | 92% | 81% | 83% | |
| Inseto | Tendência a perseguir a presa | 52% | 48% | 35% | 27% | 30% | 43% |
| Eficiência na captura da presa | 69% | 67% | 71% | 41% | 83% | 78% |
- "Tendência a perseguir a presa" é a porcentagem de testes em que o sujeito persegue a presa potencial, e a perseguição começa quando a Portia se aproxima da presa ou sacode sua teia.[3]:428-429
- "Eficiência na captura da presa" é a porcentagem de perseguições em que o sujeito captura a presa.[3]:428-429
- "(Q)", "(TN)" e "(SL)" identificam P. fimbriata de Queensland, Território do Norte e Sri Lanka.[3]:425
- As presas utilizadas foram: aranhas saltadoras não especificadas; aranhas das famílias Amaurobiidae e Theridiidae; e moscas-domésticas.[3]:428
Para descansar, todas as Portia tecem uma teia horizontal com diâmetro cerca de duas vezes o comprimento do corpo, suspensa a apenas 1 a 4 mm abaixo de uma folha.[6]:496-513 A P. schultzi retorna à sua plataforma de descanso à noite. Ao comer uma presa ao entardecer sem uma plataforma por perto, uma P. schultzi construiu uma plataforma de seda enquanto segurava a presa e continuou comendo.[5]:34
A P. schultzi não responde a presas se a luz está abaixo de 100 lux (como em um restaurante com iluminação artificial suave),[27] mas responde a presas a distâncias de 6 a 10 cm conforme a luz aumenta. Um teste sugeriu que a caça da P. schultzi é estimulada apenas pela visão, pois presas próximas, mas escondidas, não provocaram resposta.[5]:37 Um teste preliminar mostrou que iscas obtiveram as mesmas respostas da P. schultzi que presas vivas, e testes mais detalhados com iscas mostraram que: se o alvo se movia erraticamente, a P. schultzi não o perseguia; se o alvo estava totalmente parado, a aranha se aproximava muito lentamente (entre 5 e 100 mm por minuto), com pausas longas nas etapas finais, e a sequência não foi completada em 43% dos casos; se a isca tremia no mesmo lugar, a P. schultzi se aproximava muito mais rápido, e a sequência era quase sempre completada; se a isca era puxada diretamente para longe da P. schultzi, a aranha a seguia, mais rápido se a isca fosse puxada, até um limite (a P. schultzi geralmente se move muito lentamente).[5]:34-35:37-38
Uma fêmea de P. schultzi persegue mais frequentemente aranhas saltadoras pequenas e aranhas de teia do que presas maiores. Embora capture aranhas saltadoras pequenas com mais frequência do que as maiores, é igualmente eficaz com aranhas de teia de todos os tamanhos até o dobro do tamanho da P. schultzi.[3]:437-439 Uma fêmea de P. schultzi é eficaz contra insetos até o dobro de seu tamanho quando estão presos em uma teia de uma aranha não saltadora, e contra insetos fora de teias até o tamanho da P. schultzi, enquanto raramente persegue ou captura insetos maiores no aberto. Ela raramente persegue ou captura insetos maiores em sua própria teia e é ligeiramente menos eficaz contra insetos menores em sua teia do que em outras situações.[3]:439 Os machos são menos eficientes em todos os casos.[3]:436
Um teste em 1997 mostrou que as preferências da P. schultzi por diferentes tipos de presas seguem a ordem: aranhas de teia; aranhas saltadoras; e insetos.[28]:337-339 Essas preferências se aplicam tanto a presas vivas quanto a iscas imóveis, e a espécimes de P. schultzi sem presas por 7 dias ("bem alimentados")[28]:335 e por 14 dias ("famintos").[28]:335 Espécimes sem presas por 21 dias ("extremamente famintos") não mostraram preferência por tipos de presas.[28]:339 O teste incluiu várias espécies de aranhas de teia e saltadoras como presas, e a seleção das espécies de presas não mostrou evidências de afetar os resultados.[28]:337-339 Insetos foram representados pela mosca doméstica Musca domestica.[28]:335
Em um teste, filhotes de P. schultzi capturaram Drosophila ("moscas-das-frutas")[29] quase tão frequentemente quanto aranhas. A P. schultzi recua dos voos súbitos de moscas-domésticas encontradas no aberto, mas às vezes captura moscas presas em teias.[5]:38-39 Fora de sua teia, a P. schultzi raramente captura aranhas da família Thomisidae (predadoras de emboscada sem teia, geralmente com menos de 13 mm de comprimento)[30] no aberto, pois elas frequentemente agitam as pernas dianteiras quando ameaçadas.[5]:38-39
Se uma aranha passa sob a teia de captura de uma fêmea de P. schultzi e a distância vertical é inferior a 8 cm, a P. schultzi frequentemente desce sobre a presa rápido demais para o olho humano acompanhar. Se errar, ela retorna rapidamente por sua linha de segurança até seu ponto de observação e busca outra chance, raramente errando na segunda vez. A maioria dos saltos cobre de 2 a 4 cm, pois saltos mais longos são frequentemente obstruídos pela teia.[5]:36
Ao caçar uma aranha de teia em sua própria teia, a P. schultzi caminha muito lentamente em direção à presa e, a 2 a 3 cm de distância, pausa por alguns minutos. Durante esse tempo, ela treme rapidamente com todo o corpo em intervalos regulares. Em quase todos os casos, a presa permanece imóvel. A P. schultzi nunca toca a teia como a Portia fimbriata faz.[5]:37
Diferentemente da variante de Queensland de P. fimbriata, a P. schultzi não tem táticas especiais ao caçar outras aranhas saltadoras.[28]:343
Ao caçar, fêmeas maduras de P. fimbriata, P. africana, P. labiata e P. schultzi emitem sinais olfativos que reduzem o risco de competição por presas com outras fêmeas, machos ou juvenis da mesma espécie. O efeito inibe a mímica agressiva contra uma aranha presa, mesmo que ela esteja visível ou ocupando qualquer parte de uma teia. Se uma fêmea dessas espécies de Portia detecta um macho da mesma espécie, ela estimula o macho a cortejar. Essas espécies de Portia não exibem esse comportamento ao receber sinais olfativos de membros de outras espécies de Portia.[31]
Reprodução e ciclo de vida
Antes do cortejo sexual, um macho de Portia tece uma pequena teia entre galhos ou ramos, pendura-se sob ela e ejacula sobre ela.[3]:467 Ele então absorve o sêmen em reservatórios em seus palpos,[10]:581-583 que são maiores que os das fêmeas.[10]:572-573 Fêmeas de muitas espécies de aranhas, incluindo P. schultzi, emitem feromônios voláteis no ar, que geralmente atraem machos à distância.[6]:517[32]:36
Entre a P. schultzi e algumas outras Portia, quando adultos de sexos opostos da mesma espécie se reconhecem, exibem-se a 10 a 30 cm. Os machos geralmente esperam de 2 a 15 minutos antes de iniciar uma exibição, mas às vezes a fêmea começa primeiro.[3]:461 As Portia às vezes usam "exibições propulsivas", com as quais um membro ameaça um rival da mesma espécie e sexo, e fêmeas não receptivas também ameaçam machos dessa forma.[25]:343 Uma exibição propulsiva é uma série de movimentos rápidos, como golpes, investidas, colisões e saltos.[3]:455 Em P. schultzi e algumas outras espécies, confrontos entre machos geralmente duram apenas 5 a 10 segundos, com contato apenas entre as pernas.[3]:466 Confrontos entre fêmeas de Portia são violentos[6]:518 e embates em P. schultzi duram de 20 a 60 segundos. Esses embates ocasionalmente incluem lutas que podem quebrar uma perna, mas geralmente o movimento final é uma investida. Às vezes, uma fêmea derruba a outra de costas, e a derrotada pode ser morta e comida se não se recuperar rapidamente e fugir. Se a perdedora tem um ninho, a vencedora toma posse e come os ovos presentes.[3]:466-467
Uma fêmea de P. schultzi que vê um macho pode se aproximar lentamente ou esperar. O macho então caminha ereto, exibindo-se ao agitar as pernas e palpos. Se a fêmea não fugir, ela faz uma exibição propulsiva primeiro. Se o macho mantém sua posição e ela não foge ou repete a exibição propulsiva, ele se aproxima e, se ela estiver madura, eles copulam.[3]:461-464 Se a fêmea for subadulta (a uma muda de atingir a maturidade), um macho pode coabitar em sua teia de captura.[3]:467 As Portia geralmente copulam em uma teia ou em um fio de arrasto feito pela fêmea.[6]:518 A P. schultzi copula por cerca de 100 segundos,[3]:465 enquanto outros gêneros podem levar minutos ou até horas.[6]:518[3]:465
Fêmeas de P. schultzi, como as de P. labiata, tentam matar e comer seus parceiros durante ou após a cópula, girando e investindo. Os machos esperam até que as fêmeas tenham flexionado as pernas, tornando esse ataque menos provável. Os machos também tentam descer por um fio de seda para se aproximar por cima, mas as fêmeas podem manobrar para ficar na posição superior. Se a fêmea se move, o macho salta e foge.[25]:343
Antes de estarem maduras para acasalar, fêmeas de P. schultzi e P. labiata imitam fêmeas adultas para atrair machos como presas.[33]
A P. schultzi geralmente deposita ovos em folhas mortas e marrons de cerca de 20 mm de comprimento, suspensas perto do topo de sua teia de captura, cobrindo os ovos com uma folha de seda. Se não houver uma folha morta disponível, a fêmea faz uma pequena plataforma de seda horizontal na teia de captura, deposita os ovos sobre ela e os cobre.[3]:434-435 A P. schultzi foi vista depositando ovos em uma folha enrolada na teia de Ischnothele karschi.[5][c]:33
Para a ecdise, todas as Portia tecem uma teia horizontal com diâmetro cerca de duas vezes o comprimento do corpo, suspensa a apenas 1 a 4 mm abaixo de uma folha. A aranha fica de cabeça para baixo e frequentemente desliza 20 a 30 mm durante a muda.[6]:496 As Portia tecem uma teia temporária semelhante para descansar.[6]:513 Como todos os artrópodes, as aranhas passam por mudas, e o estágio de vida antes de cada muda é chamado de "instar".[36] Os tufos distintos dos juvenis de P. schultzi aparecem no terceiro instar.[5]:33
As exúvias (peles descartadas) de P. schultzi foram encontradas tanto em suas próprias teias quanto nas de I. karschi, sugerindo que a P. schultzi realiza a muda ao ar livre. Em um caso, enquanto sua nova pele ainda estava pálida e macia, seus fieiras estavam presas na pele descartada, e a aranha girou lentamente por cerca de 90 segundos até se libertar. O corpo da aranha então escureceu rapidamente para a coloração normal, e algum tempo depois ela pendurou-se em sua postura usual de cabeça para baixo na teia.[5]:35
Ecologia
A distribuição da P. schultzi vai de Durban (na África do Sul) no sul até Melinde (no Quênia) no norte, estendendo-se para o oeste até o Rifte da África Oriental, e também na África Ocidental e Madagascar.[5]:30
A P. schultzi, junto com uma grande variedade de aranhas e insetos, é frequentemente encontrada nas teias densas e grandes de Ischnothele karschi[5][c] (cerca de 15 mm de comprimento),[5]:31 que são especialmente abundantes em áreas de mata secundária parcialmente desmatada onde florestas tropicais foram cortadas, geralmente a cerca de 1 metro acima do solo. Uma pesquisa sugeriu que há cerca de uma P. schultzi para cada três teias de I. karschi. A P. schultzi também é encontrada em sua própria teia, nas teias de outras aranhas, em troncos de árvores, nas paredes de construções e em detritos foliares.[5]:30-32
Taxonomia
A P. schultzi era uma das 17 espécies do gênero Portia até junho de 2011.[37] A Portia schultzi Karsch, 1878 é sinônima de Brettus martini Simon, 1900, Linus lesserti Lawrence, 1937 e Linus alboguttatus Lawrence, 1938.[1][3]:424[4]:88
Wanless dividiu o gênero Portia em dois grupos de espécies: o grupo schultzi, no qual os palpos dos machos têm uma apófise tibial fixa; e o grupo kenti, no qual a apófise de cada palpo dos machos tem uma articulação separada por uma membrana.[4]:87-88 O grupo schultzi inclui P. schultzi, P. africana, P. fimbriata e P. labiata.[4]:93-94, 99-100, 102-105
A Portia pertence à subfamília Spartaeinae,[38] considerada primitiva.[6]:491A filogenética molecular, uma técnica que compara o DNA de organismos para reconstruir a árvore da vida, indica que a Portia é um membro do clado Spartaeinae, que Spartaeinae é basal (muito semelhante aos ancestrais de todas as aranhas saltadoras), que o parente mais próximo da Portia é o gênero Spartaeus [en], e que os próximos mais próximos são Phaeacius e Holcolaetis [en].[39]:53
Notas
- ↑ Jackson e Blest (1982) afirmam que a resolução do mosaico receptor na retina central é um ângulo visual de 2,4 minutos de arco, correspondendo a 0-12 mm a 20 cm à frente da aranha, ou 0-18 mm a 30 cm.[19]
- ↑ A retina está no final de um tubo. A extremidade interna do tubo move-se de um lado para o outro em um a dois ciclos por segundo e gira 50° em um ciclo que leva 10 segundos.[20]
- ↑ a b O que Forster e Murphy (1986) chamam de Ischnothele karschi[5] foi chamado de Thelechoris striatipes de 1995 a 2011.[34][35]
Referências
- ↑ a b c «Taxon details Portia schultzi Karsch, 1878». World Spider Catalog. Natural History Museum Bern. Consultado em 25 de fevereiro de 2017
- ↑ a b «Lolloping - Definition and More from the Free Merriam-Webster Dictionary». Merriam-Webster, Incorporated. Consultado em 5 de julho de 2011
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao Jackson, Robert R.; Hallas, Susan E. A. (1986). «Comparative biology of jumping spiders Portia africana, P. albimana, P. fimbriata, P. labiata and P. schultzi, areanophagic, web-building jumping spiders (Araneae: Salticidae) utilisation of webs, predatory versatility, and intraspecfic interactions». New Zealand Journal of Zoology. 13 (4): 423–489. ISSN 0301-4223. doi:10.1080/03014223.1986.10422978
 . Consultado em 17 de maio de 2011
. Consultado em 17 de maio de 2011
- ↑ a b c d e f Wanless, F.R. (1978). «A revision of the spider genus Portia (Araneae: Salticidae)» (PDF). Londres: British Museum. Bulletin of the British Museum (Natural History) Zoology. 34 (3): 83–124. Consultado em 14 de junho de 2011. Cópia arquivada (PDF) em 12 de agosto de 2011
- ↑ a b c d e f g h i j k l m n o p q r s t u v Forster, Lyn M.; Murphy, Frances M. (1986). «Ecology and behaviour in Portia schultzii, with notes on related species (Araneae, Salticidae)» (PDF). The Journal of Arachnology. 14: 29–42. Consultado em 9 de setembro de 2010. Cópia arquivada (PDF) em 9 de agosto de 2017
- ↑ a b c d e f g h i j k l m Jackson, Robert R.; Hallas, Susan E.A. (1986). «Predatory versatility and intraspecies interactions of spartaeine jumping spiders (Araneae, Salticidae): Brettus adonis, B. cingulata, Cyrba algerina, and Phaeacius sp. indet». New Zealand Journal of Zoology. 13 (4): 491–520. ISSN 0301-4223. doi:10.1080/03014223.1986.10422979. Consultado em 1 de abril de 2011
- ↑ Harland, D.P.; Jackson, R.R. (Novembro de 2000). «Cues by which Portia fimbriata, an araneophagic jumping spider, distinguishes jumping-spider prey from other prey» (PDF). The Journal of Experimental Biology. 203 (Pt 22): 3485–3494. PMID 11044386. doi:10.1242/jeb.203.22.3485. Consultado em 4 de maio de 2011
- ↑ a b Harland, Duane P.; Jackson, Robert R. (2004). «Portia Perceptions: The Umwelt of an Aranephagic Jumping Spider». In: Prete, Frederick R. Complex worlds from simpler nervous systems. [S.l.]: MIT Press. pp. 5–40. ISBN 978-0-262-66174-4. Consultado em 12 de abril de 2011
- ↑ a b c Wilcox, R. Stimson; Jackson, Robert R. (1998). «Cognitive Abilities of Araneophagic Jumping Spiders». In: Balda, Russell P.; Pepperberg, Irene Maxine; Kamil, Alan C. Animal cognition in nature: the convergence of psychology and biology in laboratory and field. [S.l.]: Academic Press. ISBN 978-0-12-077030-4. Consultado em 23 de maio de 2011
- ↑ a b c Ruppert, E.E.; Fox, R.S.; Barnes, R.D. (2004). «Chelicerata: Araneae». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. pp. 571–584. ISBN 978-0-03-025982-1
- ↑ Jarman, Elizabeth A.R.; Jackson, Robert R. (1986). «The biology of Taieria erebus (Araneae, Gnaphosidae), an araneophagic spider from New New Zealand: silk utilisation and predatory versatility». New Zealand Journal of Zoology. 13 (4): 521–540. ISSN 0301-4223. doi:10.1080/03014223.1986.10422980. Consultado em 1 de abril de 2011
- ↑ Hoefler, Chad D.; Chen, Andy; Jakob, Elizabeth M. (2006). «The Potential of a Jumping Spider, Phidippus clarus, as a Biocontrol Agent» (PDF). J. Econ. Entomol. 99 (2): 432–436. ISSN 0022-0493. PMID 16686143. doi:10.1603/0022-0493-99.2.432. Consultado em 25 de abril de 2011. Cópia arquivada (PDF) em 14 de junho de 2010
- ↑ a b c d e f g Harland, D.P.; Jackson, R.R. (2000). «"Eight-legged cats" and how they see - a review of recent research on jumping spiders (Araneae: Salticidae)» (PDF). Cimbebasia. 16: 231–240. Consultado em 5 de maio de 2011. Cópia arquivada (PDF) em 18 de março de 2009
- ↑ a b Forster, Lyndsay M. (1977). «A qualitative analysis of hunting behaviour in jumping spiders (Araneae: Salticidae)». New Zealand Journal of Zoology. 4: 51–62. doi:10.1080/03014223.1977.9517936
- ↑ Hill, David Edwin (Outubro de 2010). «Use of location (relative direction and distance) information by jumping spiders (Araneae, Salticidae, Phidippus) during movement toward prey and other sighted objectives» (PDF). Peckhamia. 83 (1): 1–103. ISSN 1944-8120. Consultado em 12 de abril de 2011
- ↑ a b Jackson, Robert R. (1986). «Web Building, Predatory Versatility, and the Evolution of the Salticidae». In: Shear, William A. Spiders - webs, behavior, and evolution. [S.l.]: Stanford University Press. 492 páginas. ISBN 978-0-8047-1203-3. Consultado em 22 de maio de 2011
- ↑ a b Richman, David B.; Jackson, Robert R. (1992). «A review of the ethology of jumping spiders (Araneae, Salticidae)» (PDF). Bulletin of the British Arachnology Society. 9 (2): 33–37. Consultado em 12 de maio de 2011
- ↑ Piper, Ross (2007). «The Quest for Food: Portia spider» (PDF). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. Westport, CT 06881: Greenwood Press. pp. 98–100. ISBN 978-0-313-33922-6. Consultado em 31 de março de 2011
- ↑ a b Jackson, R.R.; Blest, A.D. (1982). «The distances at which a primitive jumping spider, Portia fimbriata, makes visual discriminations» (PDF). Journal of Experimental Biology. 97: 441–445. doi:10.1242/jeb.97.1.441a. Consultado em 12 de maio de 2011
- ↑ Land, Michael F.; Nilsson, Dan-Eric (2006). «General-purpose and special-purpose visual systems» (PDF). In: Warrant, Eric; Nilsson, Dan-Eric. Invertebrate vision. [S.l.]: Cambridge University Press. pp. 167–210, 547. ISBN 978-0-521-83088-1. Consultado em 21 de junho de 2011 Parâmetro desconhecido
|page1=ignorado (ajuda) - ↑ Ruppert, E.E.; Fox, R.S.; Barnes, R.D. (2004). «Arthropoda: Sense organs». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. pp. 532–537. ISBN 978-0-03-025982-1
- ↑ Ruppert, E.E.; Fox, R.S.; Barnes, R.D. (2004). «Chelicerata: Form». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. p. 555. ISBN 978-0-03-025982-1
- ↑ Ruppert, E.E.; Fox, R.S.; Barnes, R.D. (2004). «Chelicerata: Arachnida». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. pp. 559–564. ISBN 978-0-03-025982-1
- ↑ a b c Wilcox, S.; Jackson, R. (2002). «Jumping Spider Tricksters» (PDF). In: Bekoff, M.; Allen, C.; Burghardt, G.M. The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition. [S.l.]: MIT Press. pp. 27–34. ISBN 978-0-262-52322-6. Consultado em 12 de maio de 2011. Cópia arquivada (PDF) em 21 de julho de 2011
- ↑ a b c Jackson, Robert R.; Pollard, Simon D. (1997). «Jumping spiders mating strategies: sex among cannibals in and out of webs». In: Choe, Jae C.; Crespi, Bernard J. The Evolution of Mating Systems in Insects and Arachnids. [S.l.]: Cambridge University Press. pp. 340–351, 387. ISBN 978-0-521-58976-5. Consultado em 6 de junho de 2011
- ↑ Wilcox, R. Stimson; Jackson, Robert R.; Gentile, Kristen (1996). «Spiderweb smokescreens: spider trickster uses background noise to mask stalking movements» (PDF). Animal Behaviour. 51 (2): 313–326. CiteSeerX 10.1.1.583.2268. ISSN 0003-3472. doi:10.1006/anbe.1996.0031. Consultado em 23 de maio de 2011
- ↑ Story, Derrick (2003). «Quick Reference Tables». Digital Video Pocket Guide. [S.l.]: O'Reilly Media, Inc. p. 99. ISBN 978-0-596-00523-8
- ↑ a b c d e f g Li, Daiqin; Jackson, Robert R.; Barrion, Alberto (1997). «Prey preferences of Portia labiata, P. africana, and P. schultzi, araneophagic jumping spiders (Araneae: Salticidae) from the Philippines, Sri Lanka, Kenya, and Uganda». New Zealand Journal of Zoology. 24 (4): 333–349. doi:10.1080/03014223.1997.9518129
- ↑ Solomon, Eldra Pearl; Berg, Linda R.; Martin, Diana W. (2004). «Genes and Development». Biology 7 ed. [S.l.]: Cengage Learning. p. 319. ISBN 978-0-534-49276-2. Consultado em 12 de julho de 2011
- ↑ Burton, Maurice; Burton, Robert (2002). «Crab spider». The international wildlife encyclopedia, Volume 10 3 ed. [S.l.]: Marshall Cavendish. p. 580. ISBN 978-0-7614-7266-7. Consultado em 12 de julho de 2011
- ↑ Willey, Marianne B.; Jackson, Robert R. (1993). «Olfactory cues from conspecifics inhibit the web-invasion behavior of Portia, web-invading araneophagic jumping spiders (Araneae: Salticidae)». Canadian Journal of Zoology. 71 (7): 1415–1420. doi:10.1139/z93-195
- ↑ Gaskett, A.C. (2007). «Spider sex pheromones: emission, reception, structures, and functions». Biological Reviews. 82 (1): 27–48. PMID 17313523. doi:10.1111/j.1469-185X.2006.00002.x
- ↑ Roberts, J. Andrew; Taylor, Phillip W.; Uetz, George W. (2003). «Kinship and food availability influence cannibalism tendency in early-instar wolf spiders (Araneae: Lycosidae)» (PDF). Behavioral Ecology and Sociobiology. 54 (4): 416–422. doi:10.1007/s00265-003-0646-8. Consultado em 18 de junho de 2011
- ↑ Coyle, F. A. (1995). «A revision of the funnelweb mygalomorph spider subfamily Ischnothelinae (Aranea: Dipluridae)» (PDF). American Museum of Natural History. Bulletin of the American Museum of Natural History. 226: 117–128. Consultado em 5 de julho de 2011
- ↑ Platnick, Norman I.; Raven, Robert; Schuh, Toby; Choi, Ryan (2011). «World Spider Catalog - Thelechoris striatipes». The American Museum of Natural History. Consultado em 5 de junho de 2011
- ↑ Ruppert, E.E.; Fox, R.S.; Barnes, R.D. (2004). «Arthropoda: Body wall». Invertebrate Zoology 7 ed. [S.l.]: Brooks / Cole. pp. 521–525. ISBN 978-0-03-025982-1
- ↑ «Gen. Portia Karsch, 1878». World Spider Catalog. Natural History Museum Bern. Consultado em 25 de fevereiro de 2017
- ↑ Barrion, A.T.; Litsinger, J.A. (1995). Riceland spiders of South and Southeast Asia. [S.l.]: International Rice Research Institute / C.A.B. International. p. 45. ISBN 978-0-85198-967-9
- ↑ Maddison, Wayne P.; Bodner, Melissa R.; Needham, Karen M. (2008). «Salticid spider phylogeny revisited, with the discovery of a large Australasian clade (Araneae: Salticidae)» (PDF). Zootaxa. 1893: 49–64. ISSN 1175-5334. doi:10.11646/zootaxa.1893.1.3. Consultado em 14 de junho de 2011
Ligações externas
- Salticidae: Diagnostic Drawings Library - Portia schultzi por Jerzy Proszynski.