Kryptops
Kryptops
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Cretáceo Inferior (Aptiano a Albiano), 115–110 Ma | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Nome binomial | |||||||||||||||||||
| †Kryptops palaios Sereno & Brusatte, 2008 | |||||||||||||||||||
![Maxila [en] do holótipo (osso da mandíbula superior) Kryptops](./_assets_/0c70a452f799bfe840676ee341124611/Kryptops.jpg)
Kryptops (que significa "rosto coberto") é um gênero extinto de dinossauros terópodes abelissaurídeos do Cretáceo Inferior do Níger. É conhecido a partir de uma maxila parcial (osso da mandíbula superior) encontrada na localidade de Gadoufaoua, no oeste do deserto de Ténéré, em rochas da formação Elrhaz, de idade Aptiano-Albiano. Os fósseis foram coletados em 2000 por uma expedição da Universidade de Chicago no Níger, liderada pelo paleontólogo americano Paul Sereno. Eles foram então descritos em 2008 por Sereno e Steve Brusatte. O gênero contém uma única espécie, Kryptops palaios. Sereno e Brusatte atribuíram vários restos pós-cranianos a Kryptops, mas estudos posteriores mostraram que esses elementos pertencem a um terópode alossauroide, deixando Kryptops conhecido apenas pela maxila incompleta.
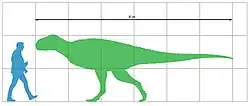
Kryptops é um dos membros mais antigos conhecidos de Abelisauridae, o que o torna vital para a compreensão da evolução desta família. Seu comprimento é estimado em cerca de 6 a 7 m, tornando-o menor que gêneros posteriores como Carnotaurus. No entanto, Kryptops é um dinossauro carnívoro de grande porte com dentes altamente serrilhados (com muitos dentículos) e mandíbulas superiores robustas. A superfície externa de sua maxila apresenta estrias profundas, sulcos e rugosidades, indicando que era coberta por uma estrutura tegumentar firmemente aderida, como queratina.
Descoberta e nomeação
Em 2000, uma expedição conduzida pela Universidade de Chicago, liderada pelo paleontólogo americano Paul Sereno e financiada pela National Geographic Society, explorou afloramentos de arenito fossilífero em um local conhecido como Gadoufaoua, na borda oeste do deserto de Ténéré, no Níger.[1][2] Essas camadas pertencem à formação Elrhaz, que data das idades Aptiana e Albiana do Cretáceo Inferior, cerca de 112 milhões de anos atrás. Durante a expedição, uma maxila (principal osso portador de dentes da mandíbula superior) de um terópode foi coletada a 15 m de distância de um conjunto de elementos pós-cranianos [en] consistindo em três vértebras dorsais (costas), duas costelas, o sacro e a pelve. Esses restos foram então transportados para a Universidade de Chicago para estudo e preparação [en] antes de serem devolvidos ao Museu Nacional do Níger [en] e depositados sob o número de catálogo MNN GAD1. A maxila foi catalogada como MNN GAD1-1, enquanto os fósseis pós-cranianos foram designados como MNN GAD1-2 a MNN GAD1-8.[3]
Esses restos de terópodes foram considerados como pertencentes ao mesmo indivíduo por Paul Sereno, que, com Jeffrey Wilson [en] e Jack Conrad, os mencionou posteriormente em um artigo de 2004 como um abelissaurídeo não descrito.[4] Sereno e Steve Brusatte descreveram os fósseis como o espécime holótipo (portador do nome) de um novo gênero e espécie de abelissaurídeo em 2008, nomeado Kryptops palaios. O nome genérico deriva das palavras gregas antigas krypto, que significa "coberto", e ops, que significa "rosto", referenciando a anatomia única da maxila. O nome específico, palaios, é um termo grego que significa "antigo", em referência à idade dos fósseis em comparação com táxons relacionados.[3]
Identidade do material pós-craniano
.png)
Sereno e Brusatte atribuíram o material pós-craniano ao mesmo indivíduo da maxila com base em sua associação próxima e em supostas características de abelissaurídeos basais nas vértebras e na pelve. Kryptops não é o único terópode conhecido da formação Elrhaz; outros gêneros incluem Eocarcharia, Suchomimus e Afromimus.[3][5] Em 2012, Matthew Carrano e colegas consideraram Kryptops palaios uma quimera (espécime composto por múltiplas espécies) e afirmaram que seus restos pós-cranianos, especialmente a pelve e o sacro, poderiam na verdade pertencer a um carcharodontossaurídeo, possivelmente Eocarcharia.[6] No entanto, esses ossos não se sobrepõem ao holótipo de Eocarcharia, que consiste apenas em um elemento isolado do crânio.[3] Essa hipótese foi apoiada por estudos posteriores, que concordaram que os restos pós-cranianos pertenciam a um alossauroide, ou mais especificamente, a um carcharodontossaurídeo.[7][8] Em 2018, o paleontólogo Rafael Delcourt questionou a validade de Kryptops, sugerindo que, devido à presença de apenas uma autapomorfia (característica única) da maxila, Kryptops pode ser um nomen dubium.[9]
Em 2025, os pesquisadores Andrea Cau [en] e Alessandro Paterna concordaram com as afinidades alossauroides para o pós-crânio. No entanto, sua análise filogenética colocou este material na família Metriacanthosauridae, em vez de Carcharodontosauridae. Com base nesses resultados, eles sugeriram ainda que os metriacantossaurídeos se irradiaram pelo mundo durante o Cretáceo Inferior, como demonstrado pela presença aproximadamente simultânea de Erectopus [en] na Europa, o pós-crânio de "Kryptops" na África e Siamotyrannus na Ásia.[10]
Descrição
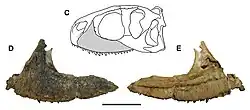
.png)
.png)
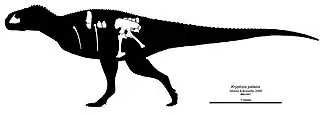
O indivíduo holótipo é grande para um abelissaurídeo, pertencendo a um adulto de cerca de 6 a 7 m de comprimento. Como outros abelissaurídeos, Kryptops possui um crânio curto e profundo e um corpo longo e esguio. A maxila tem um comprimento de fileira de dentes preservado de 15 cm e está faltando sua ponta, parte da margem alveolar (a borda superior da maxila que suporta os alvéolos dentários) e as coroas dos dentes. A maxila esquerda preserva 11 alvéolos (posições dos dentes), mas provavelmente continha 17 ou 18 alvéolos quando completa, com base no gênero relacionado Rugops. Há uma fileira de forames neurovasculares [en] acima da margem alveolar, uma característica dos abelissauroides.[3]
A face externa da maxila é extremamente rugosa, com fossas e sulcos vasculares curtos adornando as superfícies. Essa condição é semelhante à de outros abelissaurídeos e alguns carcharodontossaurídeos. Isso pode implicar um tegumento queratinoso nas maxilas em vez de escamas, o que deu nome ao gênero, embora o crânio de Carnotaurus fosse escamoso.[11][12] Sereno e Brusatte (2008) interpretaram a textura externa da maxila, com seus sulcos lineares curtos, como uma característica diagnóstica de Kryptops, distinguindo-o de outros abelissaurídeos.[3] No entanto, Delcourt (2018) observou que essa mesma condição é observável em Rugops e Majungasaurus e, portanto, não é única de Kryptops.[9]
A maxila holótipo arqueia medialmente (para dentro) em direção à articulação com a pré-maxila, resultando em um crânio largo e curto, como em seus parentes. A seção frontal da maxila é curta e profunda, ainda mais curta que a de outros abelissaurídeos.[3] A porção proximal (em direção ao centro do corpo) do ramo posterior tem margens dorsal (superior) e ventral (inferior) subparalelas, mas estas são recortadas em vez de lisas, uma característica única do gênero e da espécie. Delcourt identificou isso como a única autapomorfia válida de Kryptops.[9] As bordas dorsal e ventral da fenestra promaxilar ficam ocultas em vista lateral pela parede lateral da fossa antorbital, semelhante a Majungasaurus, Abelisaurus e outros abelissaurídeos.[3]
As placas interdentais [en] são fundidas e texturizadas com estrias semelhantes à anatomia das placas interdentais de outros abelissaurídeos. A superfície medial (voltada para dentro) da maxila está quebrada, expondo os dentes de substituição. Existem vários dentes completos preservados dentro da maxila e expostos ao longo da fileira de dentes. Suas coroas são relativamente planas e largas, como em outros abelissaurídeos, e têm cerca de 15 serrilhas [en] a cada 5 mm. Isso é comparável à contagem de serrilhas de abelissaurídeos indeterminados de Marrocos e do Egito, mas maior que a de Rugops.[3]
Classificação
Abelisauridae é um grupo de dinossauros terópodes que existiu durante o Cretáceo (e potencialmente antes, no Jurássico)[13][14] até o evento de extinção do Cretáceo-Paleogeno.[15] Os abelissaurídeos foram recuperados como a família irmã dos noassaurídeos dentro da superfamília Abelisauroidea.[16] Kryptops e Rugops são alguns dos abelissaurídeos mais antigos conhecidos, tornando-os cruciais para a compreensão da evolução e diversificação da Abelisauridae. Sereno e Brusatte observaram que a maxila apresentava uma mistura de características vistas em outros abelissaurídeos basais, bem como em membros derivados do Cretáceo Superior.[3]
A posição filogenética de Kryptops tem sido instável, pois o gênero é conhecido definitivamente apenas por uma maxila, embora seja geralmente considerado um abelissaurídeo basal. Em sua análise filogenética de 2008, Sereno e Brusatte recuperaram Kryptops como o abelissaurídeo de divergência mais precoce, seguido por Rugops e Rajasaurus, mais basal que Majungasaurus e Carnotaurus. Esta análise filogenética assumiu que os restos pós-cranianos pertenciam a Kryptops.[3] Resultados semelhantes foram recuperados por Filippi e colegas em sua descrição de 2016 de Viavenator, mesmo com Kryptops pontuado apenas para a maxila.[17] Algumas análises filogenéticas posteriores excluíram Kryptops devido à falta de material e sua instabilidade nas análises filogenéticas.[9][18][19]

Múltiplas análises filogenéticas de 2024 e 2025 recuperaram Kryptops como um táxon mais derivado, mais proximamente relacionado a brachyrostrans e majungasaurinos do que em trabalhos anteriores, e divergindo após Rugops.[20][10] Os resultados de uma dessas análises, da descrição de 2024 do abelissaurídeo Koleken [en] por Pol et al., são exibidos no cladograma abaixo. Kryptops foi recuperado como o táxon irmão do clado formado por Majungasaurinae e Brachyrostra.[21]
| Abelisauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleoambiente
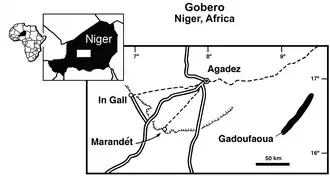
Kryptops é conhecido da formação Elrhaz do Grupo Tegama em uma área do deserto de Ténéré chamada Gadoufaoua. Apenas um espécime de Kryptops é conhecido. A formação Elrhaz consiste principalmente de arenitos fluviais com baixo relevo, grande parte dos quais está obscurecida por dunas de areia.[22] Os sedimentos são de grão grosso a médio, quase sem horizontes [en] de grão fino.[23] Kryptops viveu há cerca de 120 a 112 milhões de anos, durante as idades Aptiana a Albiana do Cretáceo médio.[3] Provavelmente viveu em habitats dominados por planícies de inundação interiores (uma zona ribeirinha).[23]

Kryptops viveu ao lado de vários outros dinossauros. Estes incluíam outros terópodes, como Eocarcharia (conhecido a partir de uma combinação quimérica de material de carcharodontossaurídeo e provável espinossaurídeo),[10] o espinossaurídeo Suchomimus e o suposto noassaurídeo Afromimus.[24] Vários megaherbívoros [en] como os hadrossauriformes Ouranosaurus e Lurdusaurus, o driossaurídeo Elrhazosaurus e dois saurópodes, o rebbachissaurídeo Nigersaurus e um titanossauro não nomeado, foram desenterrados de Gadoufaoua. Juntos, eles compõem uma das poucas associações de megaherbívoros com um equilíbrio de saurópodes e grandes ornitópodes. Crocodilomorfos como Sarcosuchus, Anatosuchus, Araripesuchus e Stolokrosuchus [en] também viveram lá. Além disso, restos de um pterossauro ornitocheirídeo, tartarugas, peixes ósseos, um tubarão hibodonte [en] e bivalves foram encontrados. A fauna aquática consiste inteiramente em habitantes de água doce.[3][5][23]
Referências
- ↑ Koppes, Steve (2000). «Dinosaur Expedition 2000 allows us to join team in Niger». chronicle.uchicago.edu. Consultado em 21 de março de 2025
- ↑ National Geographic Press Release (7 de setembro de 2003). «Huge fossil crocodilian discovered». North Texas e-News. Consultado em 21 de março de 2025
- ↑ a b c d e f g h i j k l m Sereno, Paul C.; Brusatte, Stephen L. (2008). «Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger». Acta Palaeontologica Polonica. 53 (1): 15–46. doi:10.4202/app.2008.0102
 . hdl:20.500.11820/5d55ed2e-52f2-4e4a-9ca1-fd1732f2f964
. hdl:20.500.11820/5d55ed2e-52f2-4e4a-9ca1-fd1732f2f964
- ↑ Sereno, Paul C.; Wilson, Jeffrey A.; Conrad, Jack L. (7 de julho de 2004). «New dinosaurs link southern landmasses in the Mid–Cretaceous». Proceedings of the Royal Society of London. Series B: Biological Sciences. 271 (1546): 1325–1330. PMC 1691741. PMID 15306329. doi:10.1098/rspb.2004.2692
- ↑ a b Sereno, Paul C. (2017). «Early Cretaceous Ornithomimosaurs (Dinosauria: Coelurosauria) from Africa». Ameghiniana. 54 (5): 576–616. Bibcode:2017Amegh..54..576S. doi:10.5710/AMGH.23.10.2017.3155
- ↑ Carrano, Matthew T.; Benson, Roger B. J.; Sampson, Scott D. (2012). «The phylogeny of Tetanurae (Dinosauria: Theropoda)». Journal of Systematic Palaeontology. 10 (2): 211–300. Bibcode:2012JSPal..10..211C. doi:10.1080/14772019.2011.630927
- ↑ Novas, Fernando E.; Agnolín, Federico L.; Ezcurra, Martín D.; Porfiri, Juan; Canale, Juan I. (1 de outubro de 2013). «Evolution of the carnivorous dinosaurs during the Cretaceous: The evidence from Patagonia». Cretaceous Research. 45: 174–215. Bibcode:2013CrRes..45..174N. ISSN 0195-6671. doi:10.1016/j.cretres.2013.04.001. hdl:11336/102037
- ↑ Farke, Andrew A.; Sertich, Joseph J. W. (18 de abril de 2013). «An Abelisauroid Theropod Dinosaur from the Turonian of Madagascar». PLOS ONE (em inglês). 8 (4). Bibcode:2013PLoSO...862047F. ISSN 1932-6203. PMC 3630149. PMID 23637961. doi:10.1371/journal.pone.0062047
- ↑ a b c d Delcourt, Rafael (27 de junho de 2018). «Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers». Scientific Reports (em inglês). 8 (1). Bibcode:2018NatSR...8.9730D. ISSN 2045-2322. PMC 6021374. PMID 29950661. doi:10.1038/s41598-018-28154-x
- ↑ a b c Cau, Andrea; Paterna, Alessandro (maio de 2025). «Beyond the Stromer's Riddle: the impact of lumping and splitting hypotheses on the systematics of the giant predatory dinosaurs from northern Africa». Italian Journal of Geosciences. 144 (2): 162–185. doi:10.3301/IJG.2025.10
- ↑ Cerroni, M.A.; Canale, J. I.; Novas, F. E. (3 de outubro de 2021). «The skull of Carnotaurus sastrei Bonaparte 1985 revisited: insights from craniofacial bones, palate and lower jaw». Historical Biology. 33 (10): 2444–2485. Bibcode:2021HBio...33.2444C. ISSN 0891-2963. doi:10.1080/08912963.2020.1802445 – via M.A. Cerroni
- ↑ Czerkas, Stephen A.; Czerkas, Sylvia J. (1997). «The Integument and Life Restoration of Carnotaurus». In: Wolberg, D. I.; Stump, E.; Rosenberg, G. D. Dinofest International. Filadélfia: Academy of Natural Sciences. pp. 155–158
- ↑ Hendrickx, Christophe; Mateus, Octávio (30 de janeiro de 2014). «Abelisauridae (Dinosauria: Theropoda) from the Late Jurassic of Portugal and dentition-based phylogeny as a contribution for the identification of isolated theropod teeth». Zootaxa (em inglês). 3759 (1): 1–74. ISSN 1175-5334. PMID 24869965. doi:10.11646/zootaxa.3759.1.1
- ↑ Pol, Diego; Rauhut, Oliver W. M. (23 de maio de 2012). «A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs». Proceedings of the Royal Society B: Biological Sciences. 279 (1741): 3170–3175. PMC 3385738. PMID 22628475. doi:10.1098/rspb.2012.0660
- ↑ Longrich, Nicholas R.; Pereda-Suberbiola, Xabier; Jalil, Nour-Eddine; Khaldoune, Fatima; Jourani, Essaid (1 de agosto de 2017). «An abelisaurid from the latest Cretaceous (late Maastrichtian) of Morocco, North Africa». Cretaceous Research. 76: 40–52. Bibcode:2017CrRes..76...40L. ISSN 0195-6671. doi:10.1016/j.cretres.2017.03.021
- ↑ Rauhut, Oliver W. M.; Carrano, Matthew T. (1 de novembro de 2016). «The theropod dinosaur Elaphrosaurus bambergi Janensch, 1920, from the Late Jurassic of Tendaguru, Tanzania». Zoological Journal of the Linnean Society. 178 (3): 546–610. ISSN 0024-4082. doi:10.1111/zoj.12425
- ↑ Filippi, Leonardo S.; Méndez, Ariel H.; Juárez Valieri, Rubén D.; Garrido, Alberto C. (1 de junho de 2016). «A new brachyrostran with hypertrophied axial structures reveals an unexpected radiation of latest Cretaceous abelisaurids». Cretaceous Research. 61: 209–219. Bibcode:2016CrRes..61..209F. ISSN 0195-6671. doi:10.1016/j.cretres.2015.12.018. hdl:11336/149906
- ↑ Zaher, Hussam; Pol, Diego; Navarro, Bruno A.; Delcourt, Rafael; Carvalho, Alberto B. (2020). «An Early Cretaceous theropod dinosaur from Brazil sheds light on the cranial evolution of the Abelisauridae». PaleoVol (6). ISSN 1631-0683. doi:10.5852/cr-palevol2020v19a6. hdl:11336/153682
- ↑ Gianechini, Federico A.; Apesteguía, Sebastián; Landini, Walter; Finotti, Franco; Juárez Valieri, Rubén; Zandonai, Fabiana (1 de maio de 2015). «New abelisaurid remains from the Anacleto Formation (Upper Cretaceous), Patagonia, Argentina». Cretaceous Research. 54: 1–16. Bibcode:2015CrRes..54....1G. ISSN 0195-6671. doi:10.1016/j.cretres.2014.11.009. hdl:11336/37727
- ↑ Hendrickx, Christophe; Cerroni, Mauricio A; Agnolín, Federico L; Catalano, Santiago; Ribeiro, Cátia F; Delcourt, Rafael (1 de dezembro de 2024). «Osteology, relationship, and feeding ecology of the theropod dinosaur Noasaurus leali, from the Late Cretaceous of North-Western Argentina». Zoological Journal of the Linnean Society (em inglês). 202 (4). ISSN 0024-4082. doi:10.1093/zoolinnean/zlae150
- ↑ Pol, Diego; Baiano, Mattia Antonio; Černý, David; Novas, Fernando; Cerda, Ignacio A.; Pittman, Michael (21 de maio de 2024). «A new abelisaurid dinosaur from the end Cretaceous of Patagonia and evolutionary rates among the Ceratosauria». Cladistics. 40 (3): 307–356. PMID 38771085. doi:10.1111/cla.12583
- ↑ Sereno, Paul C.; Beck, Allison L.; Dutheil, Didier B.; Larsson, Hans C. E.; Lyon, Gabrielle H.; Moussa, Bourahima; Sadleir, Rudyard W.; Sidor, Christian A.; Varricchio, David J.; Wilson, Gregory P.; Wilson, Jeffrey A. (12 de novembro de 1999). «Cretaceous Sauropods from the Sahara and the Uneven Rate of Skeletal Evolution Among Dinosaurs» (PDF). Science. 286 (5443): 1342–1347. PMID 10558986. doi:10.1126/science.286.5443.1342
- ↑ a b c Sereno, Paul C.; Wilson, Jeffrey A.; Witmer, Lawrence M.; Whitlock, John A.; Maga, Abdoulaye; Ide, Oumarou; Rowe, Timothy A. (21 de novembro de 2007). «Structural Extremes in a Cretaceous Dinosaur». PLOS ONE (em inglês). 2 (11). Bibcode:2007PLoSO...2.1230S. ISSN 1932-6203. PMC 2077925. PMID 18030355. doi:10.1371/journal.pone.0001230
- ↑ Cerroni, M.A.; Agnolin, F.L.; Brissón Egli, F.; Novas, F.E. (2019). «The phylogenetic position of Afromimus tenerensis Sereno, 2017 and its paleobiogeographical implications». Journal of African Earth Sciences. 159. Bibcode:2019JAfES.15903572C. doi:10.1016/j.jafrearsci.2019.103572

.jpg)