Ambondro mahabo
Ambondro mahabo
| |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Jurássico Médio Bathoniano | |||||||||||
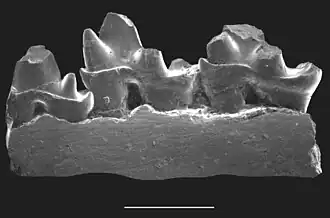 Figura 1. Mandíbula de Ambondro, vista na perspectiva lingual (do lado da língua). A escala é de 1 mm. | |||||||||||
| Classificação científica | |||||||||||
| |||||||||||
| Nome binomial | |||||||||||
| †Ambondro mahabo''' | |||||||||||
| Clado | |||||||||||
Ambondro mahabo é um mamífero do Jurássico Médio (Batoniano) da formação Isalo III [en] (cerca de 167 milhões de anos atrás) de Madagascar. Única espécie descrita do gênero Ambondro, é conhecido a partir de uma mandíbula inferior fragmentária com três dentes, interpretados como o último pré-molar e os dois primeiros molares. O pré-molar consiste em uma cúspide central com uma ou duas cúspides menores e um cíngulo [en] (plataforma) no lado interno, ou lingual, do dente. Os molares também possuem tal cíngulo lingual. Eles consistem em dois grupos de cúspides: um trigonídeo de três cúspides na frente e um talonídeo com uma cúspide principal, uma cúspide menor e uma crista na parte de trás. As características do talonídeo sugerem que Ambondro tinha molares tribosfênicos, o arranjo básico de características molares também presente em mamíferos marsupiais e placentários. É o mamífero mais antigo conhecido com dentes supostamente tribosfênicos; na época de sua descoberta, antecedia o segundo exemplo mais antigo em cerca de 25 milhões de anos.
Após sua descrição em 1999, Ambondro foi interpretado como um parente primitivo de Tribosphenida (marsupiais, placentários e seus parentes extintos com dentes tribosfênicos). Em 2001, no entanto, foi publicada uma sugestão alternativa que o unia ao Ausktribosphenos do Cretáceo da Austrália e aos monotremados (as equidnas, o ornitorrinco e seus parentes extintos) no clado Australosphenida, que teria adquirido molares tribosfênicos independentemente dos marsupiais e placentários. Os gêneros argentinos do Jurássico Asfaltomylos e Henosferus [en] e o australiano do Cretáceo Bishops [en] foram posteriormente adicionados aos Australosphenida, e novos trabalhos sobre o desgaste em dentes de australosfenídeos questionaram se esses animais, incluindo Ambondro, de fato tinham dentes tribosfênicos. Outros paleontólogos desafiaram este conceito de Australosphenida e, em vez disso, propuseram que Ambondro não está intimamente relacionado com Ausktribosphenos mais monotremados, ou que os monotremados não são australosfenídeos e que os australosfenídeos restantes estão relacionados aos placentários.
Descoberta e contexto
Ambondro mahabo foi descrito por uma equipe liderada por John Flynn em um artigo de 1999 na revista Nature. O nome científico deriva da aldeia de Ambondromahabo [en], perto da qual o fóssil foi encontrado. É conhecido do Batoniano (Jurássico Médio, cerca de 167 milhões de anos atrás) da bacia de Mahajanga, no noroeste de Madagascar, na unidade Isalo III [en], a mais jovem das três camadas de rocha que compõem o "Grupo" Isalo. Esta unidade também rendeu dentes de crocodiliformes e plesiossauros e restos do saurópode Lapparentosaurus [en].[1]
Descrição
Ambondro foi descrito com base em uma mandíbula direita (maxilar inferior) fragmentária com três dentes, interpretados como o último pré-molar (p-último) e os dois primeiros molares (m1 e m2). Está na coleção da Universidade de Antananarivo como espécime UA 10602. Em relação a outros mamíferos primitivos, é pequeno. Cada um dos dentes tem um cíngulo [en] (plataforma) proeminente no lado interno (lingual).[2] O p-último tem uma forte cúspide central. Há uma cúspula (pequena cúspide) na parte de trás do dente e provavelmente outra no canto frontal interno. Este dente se assemelha aos molares dos simetrodontes, um grupo de mamíferos primitivos, mas a cúspide traseira é menor que o metaconídeo [en] dos simetrodontes.[3]
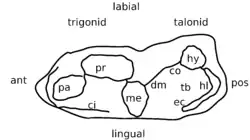
A metade da frente do m1 e do m2 consiste no trigonídeo, um grupo de três cúspides formando um triângulo: o paraconídeo na frente do lado interno, o protoconídeo no meio do lado externo (labial) e o metaconídeo na parte de trás do lado interno (ver Figura 2). As três cúspides formam um ângulo reto entre si no protoconídeo, de modo que o trigonídeo é descrito como "aberto".[2] O paraconídeo é mais alto que o metaconídeo.[4] Na margem frontal, está presente um cíngulo que é dividido em duas pequenas cúspides.[5] Ao contrário de vários mamíferos tribosfênicos primitivos e parentes próximos, não há cúspula adicional atrás do metaconídeo.[6] Na parte de trás do trigonídeo, a crista conhecida como metacristídeo distal está localizada relativamente perto do lado externo do dente e é contínua com outra crista, a crista oblíqua, que por sua vez está conectada à parte de trás do dente.[2]
O talonídeo, outro grupo de cúspides, constitui a parte de trás do dente. É mais largo do que longo[4] e contém uma cúspide bem desenvolvida, o hipoconídeo, no lado externo e uma depressão, a bacia do talonídeo, no meio. A crista oblíqua se conecta ao hipoconídeo. A cúspide menor do hipoconulídeo está presente em direção ao lado interno do dente, e o hipoconídeo e o hipoconulídeo são conectados por uma aresta de corte que é sugestiva da presença de uma cúspide metacone nos molares superiores. Mais para o lado interno, uma crista, o entocristídeo, margeia a bacia do talonídeo; no m1, ela é inchada e no m2, contém duas pequenas cúspulas, mas uma cúspide entoconídea distinta está ausente.[2] Este entocristídeo é contínuo com o cíngulo lingual.[3]
Facetas de desgaste são áreas de um dente que mostram evidência de contato com um dente na mandíbula oposta quando os dentes são unidos (o que é conhecido como oclusão).[7] Flynn e colegas identificaram duas facetas de desgaste nas margens frontal e posterior da bacia do talonídeo; eles argumentam que essas facetas de desgaste sugerem a presença de um protocone (outra cúspide no lado externo do dente) nos molares superiores.[8] Em um artigo de 2005 sobre Asfaltomylos, um mamífero primitivo aparentado da Argentina, Thomas Martin e Oliver Rauhut contestaram a presença dessas facetas de desgaste dentro da bacia do talonídeo em Ambondro e, em vez disso, identificaram facetas de desgaste nas cúspides e cristas que cercam a bacia. Eles propuseram que o desgaste no talonídeo dos australosfenídeos ocorre principalmente nas bordas, não na própria bacia do talonídeo, e que os australosfenídeos podem não ter tido um protocone funcional.[9]
Interpretações
| |||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||
| Figura 3. Visões alternativas das relações do Ambondro. Acima, Rougier et al., (2007, fig. 9): os australosfenídeos, incluindo monotremados e Ambondro, são distintos dos boreosfenídeos.[Note 1] Abaixo, Woodburne et al. (2003, fig. 3): os australosfenídeos, incluindo Ambondro, mas excluindo os monotremados, estão intimamente relacionados aos placentários. Muitos táxons foram omitidos de ambas as árvores para maior clareza. |
Em seu artigo, Flynn e colegas descreveram Ambondro como o mamífero mais antigo com molares tribosfênicos — o tipo de molar básico dos mamíferos metatérios (marsupiais e seus parentes extintos) e eutérios (placentários e seus parentes extintos), caracterizado pelo contato da cúspide do protocone nos molares superiores com a bacia do talonídeo nos molares inferiores durante a mastigação. A descoberta de Ambondro foi considerada como uma extensão do alcance temporal conhecido dos mamíferos tribosfênicos em 25 milhões de anos para o passado.[10] Consequentemente, Flynn e colegas argumentaram contra a visão predominante de que os mamíferos tribosfênicos se originaram nos continentes do norte (Laurásia) e, em vez disso, propuseram que sua origem estaria no sul (Gondwana).[11] Eles citaram a retenção de um metacristídeo distal e um trigonídeo "aberto" como caracteres que separam Ambondro de tribosfenídeos mais modernos.[2]
Em 2001, Zhe-Xi Luo [en] e colegas propuseram alternativamente que um padrão de molar tribosfênico surgiu duas vezes — uma vez dando origem aos marsupiais e placentários (Boreosphenida), e outra vez produzindo Ambondro, o Ausktribosphenos do Cretáceo da Austrália e os monotremados vivos, que apareceram pela primeira vez no Cretáceo (unidos como Australosphenida).[12] Eles caracterizaram Australosphenida pela presença compartilhada de um cíngulo no canto frontal externo dos molares inferiores, um talonídeo curto e largo, um trigonídeo relativamente baixo e um último pré-molar inferior triangulado.[13]
Também em 2001, Denise Sigogneau-Russell e colegas, em sua descrição do mais antigo mamífero tribosfênico da Laurásia, Tribactonodon, concordaram com a relação entre Ausktribosphenos e monotremados, mas argumentaram que Ambondro era mais próximo dos tribosfenídeos da Laurásia do que de Ausktribosphenos e monotremados. Como evidência contra a integridade dos Australosphenida, eles citaram a presença de cíngulos linguais em vários mamíferos não australosfenídeos; a presença de duas cúspides no cíngulo anterior em Ambondro, bem como em alguns boreosfenídeos; a aparência diferente do pré-molar em Ambondro (plano) e Ausktribosphenos (quadrado); e o contraste entre os talonídeos de Ambondro (com um hipocônido bem desenvolvido no lado labial) e Ausktribosphenos (quadrado).[5]
No ano seguinte, Luo e colegas publicaram uma análise mais completa, confirmando sua conclusão anterior e adicionando o Bishops [en] do Cretáceo da Austrália aos Australosphenida.[14] Eles mencionaram a condição do hipoconulídeo, que é inclinado para a frente, em vez de para trás como nos boreosfenídeos, como um caráter adicional dos australosfenídeos[15] e notaram que Ausktribosphenos e monotremados estavam unidos, com exclusão de Ambondro, pela presença de um entalhe em forma de V no metacristídeo distal.[16] No mesmo ano, Asfaltomylos foi descrito do Jurássico da Argentina como outro australosfenídeo. Em contraste com Ambondro, este animal não tinha um metacristídeo distal e não tinha um cíngulo lingual tão bem desenvolvido.[17]
No entanto, em 2003, Michael Woodburne [en] e colegas revisaram a análise cladística publicada por Luo e colegas, fazendo várias alterações nos dados, particularmente nos monotremados.[18] Seus resultados (Figura 3, baixo) desafiaram a divisão entre Australosphenida e Boreosphenida, conforme proposto por Luo et al. Em vez disso, eles excluíram os monotremados dos Australosphenida e colocaram os australosfenídeos restantes próximos aos Eutheria, com Ambondro mais aparentado a Asfaltomylos.[19] Em 2007, Guillermo Rougier e colegas descreveram outro australosfenídeo, Henosferus [en], do Jurássico da Argentina; eles argumentaram contra uma relação entre Eutheria e Australosphenida, mas foram ambivalentes sobre a colocação dos monotremados dentro de Australosphenida.[20] Com base em parte no trabalho anterior de Martin e Rauhut sobre facetas de desgaste em australosfenídeos, eles questionaram a presença de um verdadeiro protocone funcional nos molares superiores de australosfenídeos não monotremados — nenhum dos quais é conhecido a partir de dentes superiores — e, consequentemente, sugeriram que os australosfenídeos podem, afinal, não ter tido dentes verdadeiramente tribosfênicos.[21]
Notas
- ↑ Compare árvores semelhantes em Luo et al. (2001, fig. 1), Luo et al. (2002, fig. 1), Rauhut et al. (2002, fig. 3), que incluíram menos espécies de australosfenídeos.
Referências
- ↑ Flynn et al., 1999, pp. 57–58
- ↑ a b c d e Flynn et al., 1999, p. 58
- ↑ a b Flynn et al., 1999, fig. 3
- ↑ a b Rauhut et al., 2002, p. 167
- ↑ a b Sigogneau-Russell et al., 2001, p. 146
- ↑ Flynn et al., 1999, fig. 2
- ↑ Luo et al., 2002, pp. 22, 29
- ↑ Flynn et al., 1999, p. 59
- ↑ Martin e Rauhut, 2005, pp. 422–423
- ↑ Flynn et al., 1999, p. 57; Rougier et al., 2007, p. 23
- ↑ Flynn et al., 1999, p. 60
- ↑ Luo et al., 2001, p. 56
- ↑ Luo et al., 2001, pp. 53, 56
- ↑ Luo et al., 2002, fig. 1
- ↑ Luo et al., 2002, p. 23
- ↑ Luo et al., 2002, p. 22
- ↑ Rauhut et al., 2002, p. 166
- ↑ Woodburne, 2003, pp. 233–235
- ↑ Woodburne, 2003, fig. 5; Woodburne et al., 2003, fig. 3
- ↑ Rougier et al., 2007, p. 31
- ↑ Rougier et al., 2007, pp. 24–25
Bibliografia
- Flynn, J.J., Parrish, J.M., Rakotosamimanana, B., Simpson, W.F. e Wyss, A.R. 1999. A Middle Jurassic mammal from Madagascar (assinatura necessária). Nature 401:57–60.
- Luo, Z.-X., Cifelli, R.L. and Kielan-Jaworowska, Z. 2001. Dual origin of tribosphenic mammals (assinatura necessária). Nature 409:53–57.
- Luo, Z.-X., Kielan-Jaworowska, Z. e Cifelli, R.L. 2002. In quest for a phylogeny of Mesozoic mammals. Acta Palaeontologica Polonica 47(1):1–78.
- Martin, T. e Rauhut, O.W.M. 2005. Mandible and dentition of Asfaltomylos patagonicus (Australosphenida, Mammalia) and the evolution of tribosphenic teeth (assinatura necessária). Journal of Vertebrate Paleontology 25(2):414–425.
- Rauhut, O.W.M., Martin, T., Ortiz-Jaureguizar, E. e Puerta, P. 2002. A Jurassic mammal from South America (assinatura necessária). Nature 416:165–168.
- Rougier, G.W., Martinelli, A.G., Forasiepi, A.M. e Novacek, M.J. 2007. New Jurassic mammals from Patagonia, Argentina: A reappraisal of australosphenidan morphology and interrelationships. American Museum Novitates 3566:1–54.
- Sigogneau-Russell, D., Hooker, J.J. e Ensom, P.C. 2001. The oldest tribosphenic mammal from Laurasia (Purbeck Limestone Group, Berriasian, Cretaceous, UK) and its bearing on the 'dual origin' of Tribosphenida (assinatura necessária). Comptes Rendus de l'Académie des Sciences, Series IIA (Earth and Planetary Science) 333(2):141–147.
- Woodburne, M.O. 2003. Monotremes as pretribosphenic mammals (assinatura necessária). Journal of Mammalian Evolution 10(3):195–248.
- Woodburne, M.O., Rich, T.H. e Springer, M.S. 2003. The evolution of tribospheny and the antiquity of mammalian clades (assinatura necessária). Molecular Phylogeny and Evolution 28(2):360–385.
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||




