Bignoniaceae
Bignoniaceae
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Ypresiano-recente[1] | |||||||||||||||||||
 Catalpa bignonioides). | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Género-tipo | |||||||||||||||||||
| Bignonia L. | |||||||||||||||||||
| Grupos monofiléticos e géneros | |||||||||||||||||||
| |||||||||||||||||||
| Sinónimos | |||||||||||||||||||
| |||||||||||||||||||







_(8182060215).jpg)


_Sandwith_(5164116042).jpg)
.jpg)




.jpg)
.jpg)





.jpg)



.jpg)






.jpg)
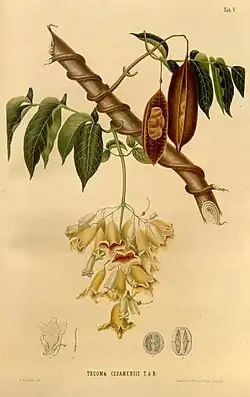
Bignoniaceae é uma família de plantas com flor da ordem das Lamiales, que agrupa cerca de 790 espécies, repartidas por 110 géneros e 8 clados,[3] que inclui maioritariamente plantas lenhosas (árvores, arbustos e lianas), com vasta distribuição natural nas regiões tropicais e subtropicais, sendo pouco frequente nas regiões temperadas. Os maiores géneros são Handroanthus, com 100 espécies, Arrabidaea, com 70 espécies, Adenocalymma, com 50 espécies e Jacaranda, com 40 espécies. Vários géneros tem grande importância económica como produtores de madeira e pela utilização de muitas das suas espécies como planta ornamental.
Descrição
A família Bignoniaceae, na qual várias espécies são conhecidas pelo nome comum de bignónias ou trepadeiras-trombeta,[4][5] está integrada na ordem Lamiales, embora a sua filogenia continue pouco conhecida, sendo complexo determinar com qual das outras famílias da ordem está mais intimamente relacionada.
Quase todas as Bignoniaceae são plantas lenhosas, mas algumas são sub-lenhosas, seja como trepadeiras ou sub-arbustos. Um pequeno números de espécies são plantas herbáceas de habitats montanhosos de alta altitude, agrupadas em três géneros exclusivamente herbáceos: Tourrettia, Argylia e Incarvillea.[6] A família inclui muitas lianas, que trepam por meio de gavinhas, entrelaçando-se ou, raramente, por meio de raízes aéreas. A maior tribo da família, a tribo Bignonieae, é composta principalmente por lianas e é conhecida pela sua anatomia da madeira única.[7]
De acordo com diferentes relatos, o número de espécies da família é de cerca de 790, cerca de 810,[8] ou cerca de 860.[7] A última monografia de toda a família foi publicada em 2004.[7] Nessa obra, foram reconhecidos 104 géneros. Desde então, as técnicas de filogenética molecular esclareceram bastante as relações genéticas dentro da família, e o número de géneros aceites está agora entre 80 e 85.[9]
A família tem uma distribuição quase cosmopolita, mas é principalmente tropical, com algumas espécies nativas das zonas temperadas. O mais importante centro de diversidade encontra-se no norte da América do Sul.[7] A família foi abordada em alguns grandes projetos florísticos, como Flora da China, Flora Malesiana e Flora Neotropica. Ainda não foi abordada em outros, entre os quais as floras da Austrália e da América do Norte.
As Bignoniaceae são mais conhecidas como plantas ornamentais, especialmente os géneros Jacaranda, Tabebuia e Spathodea, cultivados em parques e jardins e utilizados em arborização urbana pelas suas flores tubulares vistosas.[10] Também são utilizadas como plantas ornamentais algumas espécies dos géneros Campsis e Pyrostegia.
Muitas outras espécies são igualmente valiosas devido ao seu extenso cultivo em jardinagem[11] e vários outros usos têm sido feitos de membros desta família.[12] Várias espécies eram de grande importância para os povos indígenas das regiões tropicais americanas.[13]
As espécies Fridericia elegans, Tanaecium bilabiata e Tanaecium excitosum são tóxicas para o gado e capazes de causar graves perdas económicas.[13]
As flores são conspícuas e maioritariamente polinizadas por abelhas, vespas, borboletas, mariposas, pássaros e morcegos, e as sementes são dispersas principalmente pelo vento.
Morfologia
- Hábito, raízes e caules
Os membros da família Bignoniaceae são árvores ou lianas, por vezes arbustos e, raramente, subarbustos ou plantas herbáceas. O principal grupo de espécies são as lenhosas com hábito escandente, as lianas, especialmente as incluídas na tribo Bignonieae, as quais apresentam uma estrutura vascular única, na qual os canais do floema se estendem para baixo até ao xilema, porque alguns segmentos do câmbio cessam a produção de xilema numa fase inicial do desenvolvimento caulinar. O número dessas ramificações, ou braços, é quatro ou um múltiplo desse número, até às 32 ramificações.[14] Quando são em número de quatro, os braços do floema apresentam uma forma de cruz, daí o nome comum «vide cruzada» que por vezes se dá a estas espécies. O floema nos braços tem tubos crivosos mais largos e menos parênquima do que o floema comum.[15]
Anatomicamente, o caule destas lianas caracteriza-se pela presença de uma variação cambial denominada xilema interrompido, com a formação de cunhas de floema que conferem ao caule em secção transversal a forma de cruz-de-Malta. Essa variação cambial deriva de quatro (ou de múltiplos de quatro) porções de um câmbio que apresenta, inicialmente, atividade normal, mas que passa a produzir menos xilema e mais floema à medida que se alonga. O floema produzido por essas porções variantes, por sua vez, apresenta elementos de tubo crivado muito largos e menos parênquima que as porções com atividade normal. A presença desta variação cambial neste grupo parece conferir flexibilidade às plantas, auxiliar na recuperação dos tecidos após danos e contribuir para a formação de xilema, além de melhorar a condução de fotossintatos.
- Folhas
Os membros desta família apresentam folhas dispostas em filotaxia oposta, podendo, em raros casos, ser alterna ou verticilada em espiral. As folhas pecíoladas são geralmente compostas, bifoliadas, trifoliadas, pinadas, ou palmadas, ocasionalmente de lâmina foliar simples. Os bordos são inteiras a serreadas. Apresentam venação peninérvea e folíolos terminais por vezes modificados em gavinhas ou ganchos.
Não apresentam estípulas, mas ocorrem com frequência pseudoestípulas, formadas em torno de botões (brotos) axilares alargados com tricomas escamiformes. Em alguns géneros ocorrem domácias, originando pequenas câmaras que podem abrigar alguns tipos de artrópodes.
- Flores
As flores são solitárias ou ocorrem em inflorescências em racemo ou em inflorescências cimosas helicoidais ou dicásicas. As inflorescências apresentam invólucro formado por brácteas involucrais ou bracteolas, persistentes ou caducas.[7]
As flores são andróginas, hipogínicas, zigomórficas (ou seja, com simetria bilateral), bissexuais e geralmente conspícuas ou mesmo muito vistosas. O cálice e a corola são distintos e pentâmeros. O cálice é sinsépalo, com as cinco sépalas soldadas. A corola é simpétala, com as cinco pétalas, frequentemente bilabiadas, fundidas numa estrutura tubular em forma de "S". Os lóbulos da corola são imbricados ou, raramente, valvados no botão, e geralmente muito mais curtos do que o tubo da corola.[7]
As flores apresentam tamanhos grandes e vistosos, com as sépalas conatas imbricadas numa estrutura única e a corola mais ou menos bilabiada, a qual pode apresentar limbos com duas divisões que aparentam dois lábios.
O androceu apresenta quatro estames didínamos (um par de estames visivelmente mais longos que o outro par), inseridos no tubo da corola, alternando com os lóbulos desta. Os membros de cada par são frequentemente conatos e o estame adaxial é geralmente estaminodal atrofiado ou ausente. Embora raramente, ocorrem espécies com cinco estames férteis ou com dois estames férteis e três estaminódios.[7]
O gineceu tem dois carpelos, formando um ovário súpero e geralmente bilocular, com estigma bilamelado, em geral cercado por um disco nectarífero. Os filetes estão unidos à corola e apresentam anteras coniventes, sagitadas, com duas tecas divaricadas (em forma de "V"). O estigma é bilobado e geralmente sensível; com um estilete presente.[7]
O ovário é unilocular no género Tourrettia e quadrilocular no géneros Eccremocarpus . A placentação é axial, mas parietal em Tourrettia. Os óvulos são numerosos.
A família Bignoniaceae não apresenta padrão em sazonalidade. A flores da maioria das espécies apresentam síndrome de melitofilia, ornitofilia e quiropterofilia, pelo que a polinização pode ser entomófila (por meio de insetos), ornitófila (por meio de aves) ou quiropterófila (por meio de morcegos). Como as flores desta família são bissexuais, são capazes de realizar autofecundação em alguns casos.
- Frutos
Os frutos são, geralmente, longos, com cápsula septícida a loculicida, podendo ser também baga ou cápsula indeiscente. As sementes achatadas, aladas ou providas de pêlos. O endosperma está ausente. Os cotilédones são profundamente bilobados, porém há exceções, como nos géneros Kigelia, Crescentia e Colea, que apresentam fruto sem cápsula indiferente e com sementes não aladas. O fruto de Colea é uma baga.
O fruto são, geralmente, longos, em cápsula bivalve, frequentemente com um replum. A deiscência é septicida ou loculicida, com sementes achatadas, aladas ou providas de pelos. As exceções são os géneros Kigelia, Crescentia e seus parentes próximos, e Colea e seus parentes próximos. Nestes, o fruto é indeiscente, em baga, e as sementes não são aladas.[7]
As sementes são pequenas. O arilo está ausente. O endosperma está geralmente ausente e, quando presente, é por vezes escasso.[7]
O número cromossómico não varia muito nas Bignoniaceae. O haplóide (número cromossómico básico) é x = 20 para quase todas as espécies amostradas, mas algumas espécies têm cromossomas muito pequenos, dificultando uma contagem precisa.[16] A presença de cromossomas B é comum nas Bignoniaceae.
Fitoquímica
O lapachol, uma naftoquinona amarela que irrita a pele, é frequentemente encontrado na madeira. Outras naftoquinonas, bem como antraquinonas, também estão presentes em várias partes da planta. A jacaranona é um quinonoide proveniente das espécies do género Jacaranda. Não existem taninos verdadeiros. Os pigmentos são principalmente flavonas, antocianinas e carotenóides.[17] Os iridoides estão geralmente presentes.[18] Outros compostos detectados nas Bignoniaceae incluem verbascosídeos, cornosídeo, quercetina, ácido ursólico, saponinas e ácido catálpico.
Usos económicos e etnobotânica
A família apresenta grande importância económica, pois fornece madeiras nobres, especialmente de espécies dos géneros Jacaranda, Tabebuia e Catalpa. Além disso, existem algumas espécies que são muito utilizadas como plantas ornamentais, especialmente dos géneros Clytostoma, Crescentia, Jacaranda, Macfadyna e Tabebuia.
Muitas espécies de Bignoniaceae têm alguma utilidade, comercial ou etnobotânica, mas as mais importantes, de longe, são aquelas plantadas como ornamentais, especialmente as árvores que produzem flores conspícuas. Os géneros Jacaranda, Campsis, Pyrostegia, Tabebuia, Catalpa, Roseodendron, Handroanthus e Crescentia têm espécies de grande importância em jardinagem e paisagismo, pelo menos em climas tropicais e subtropicais.[7][19] Várias outras, incluindo Tecoma, Podranea, Pandorea, Bignonia e Mansoa, são frequentemente cultivadas como plantas ornamentais, pelo menos em certas áreas dos trópicos e subtrópicos.[10] Muitas espécies são conhecidas em cultivo, embora raramente.[11]
A Jacaranda mimosifolia é comum como árvore de avenida. O pecíolo alado e a folha trifoliada da Crescentia alata assemelham-se a uma cruz, por isso, por vezes, é plantada nas Filipinas como símbolo religioso.
O género Handroanthus, e o género não relacionado Guaiacum (Zygophyllaceae), têm a madeira mais dura, pesada e durável dos trópicos americanos. Árvores importantes para a produção de madeira no género Handroanthus são Handroanthus heptaphyllus, Handroanthus serratifolius, Handroanthus guayacan, Handroanthus chrysanthus e Handroanthus billbergii.[13] A espécie Tabebuia rosea (incluindo Tabebuia pentaphylla) é colhida para madeira em toda a região tropical do Novo Mundo.[20] Tabebuia heterophylla e Tabebuia angustata são fontes importantes de madeira para algumas das ilhas das Caraíbas. Várias espécies de Catalpa também são árvores importantes para a produção de madeiras.
O género Paratecoma já foi a árvore madeireira mais importante da região do Rio de Janeiro, mas a exploração implacável levou-o à beira da extinção.[13] Várias das espécies raras de Bignoniaceae produzem madeira excelente, mas muitas vezes não são reconhecidas pelos madeireiros.[20]
Várias utilizações das plantas da família Bignoniaceae são conhecidas localmente. A espécie Parmentiera aculeata é cultivada pelos seus frutos comestíveis na América Central e no sul do México. As sementes em pó e, por vezes, a polpa dos frutos, de Crescentia cujete e de Crescentia alata são utilizadas na Nicarágua para fazer um refresco chamado semilla de jicaro. As espécies com aroma de cebola de Mansoa e as espécies com aroma de cravinho de Tynanthus são utilizadas como condimentos.[7]
No norte da Colômbia, lascas dos caules da Dolichandra quadrivalvis são adicionadas ao isco que é deixada durante a noite perto das tocas dos caranguejos. Os caranguejos ficam paralisados por algumas horas após comerem a isca e são recolhidos pelos pescadores pela manhã. Os caranguejos recuperam antes de chegarem ao mercado e não foram relatados danos decorrentes do seu consumo.[13]
O Tanaecium nocturnum é a fonte de uma droga alucinógena.[21] As suas folhas e caules esmagados são usados para acalmar as abelhas enquanto se recolhe o mel por elas produzido.
A espécie Fridericia chica é a fonte de um pigmento vermelho usado na Bacia Amazónica para pintura corporal e para corante em cestaria.[7] A espécie Cybistax antisyphilitica é a fonte de um corante azul frequentemente usado no Peru. A casca de Sparattosperma leucantha é utilizada na Bolívia para produzir um corante castanho para tingir fios de algodão.[13]
As alegações médicas são inúmeras e geralmente espúrias. Gentry descreve um exemplo especialmente ridículo:[13] a casca de várias espécies de Handroanthus é vendida nos mercados sul-americanos. Casca de aparência semelhante é frequentemente vendida fraudulentamente como Handroanthus. É usada de várias maneiras para aliviar certos sintomas de certos cancros.[13] Não há evidências que comprovem que ela previne a doença ou retarda a sua progressão, como frequentemente se afirma.
A espécie Adenocalymma flavida tem sido usada para aliviar dores nas articulações e nos músculos. Um extrato da raiz da Martinella é útil no tratamento da conjuntivite e, possivelmente, de outras doenças oculares.[22]
A identificação incorreta de plantas, mesmo por botânicos, continua a ser um grande problema para a etnobotânica, sendo especialmente grave no caso das Bignoniaceae. Os espécimes usados são frequentemente estéreis e fragmentários, tornando-os quase impossíveis de identificar. As alegações médicas falsas são frequentemente baseadas em identificações erradas.[13]
Taxonomia e filogenia
A família Bignoniaceae foi publicada pela primeira vez na literatura botânica (como Bignonieae) por Antoine Laurent de Jussieu em 1789, na sua obra clássica, Genera Plantarum.[23] A etimologia da designação da família assenta no nome do género Bignonia, o género tipo para esta família, que foi validado por Carl Linnaeus em Species Plantarum em 1753.[24] O nome teve origem com Joseph Pitton de Tournefort, que o batizou em 1694, na sua influente obra Eléments de botanique ou méthode pour connaître les plantes, em homenagem ao seu benfeitor, o padre e naturalista francês Jean-Paul Bignon, bibliotecário do rei Luís XIV de França.[25]
Importantes bases para estudos futuros sobre a família foram estabelecidas entre 1789 e 1837, principalmente por Antoine Laurent de Jussieu, Karl Sigismund Kunth, Wenceslas Bojer e George Don (1798–1856) (e não pelo homónimo George Don, mais velho).[12] George Bentham e Joseph Dalton Hooker investigaram a família para a sua obra Genera Plantarum em 1876.[26] Karl Moritz Schumann escreveu uma monografia sobre Bignoniaceae em 1894 para Heinrich Gustav Adolf Engler e Karl Anton Eugen Prantl em Die Natürlichen Pflanzenfamilien. Após a monografia de Schumann, nenhum tratamento taxonómico de toda a família foi publicado até 2004.[7]
À medida que o número de espécies conhecidas aumentava gradualmente, surgiu uma grande confusão quanto à circunscrição taxonómica dos géneros. Novos géneros eram frequentemente criados para espécies que não pertenciam claramente a nenhum dos géneros descritos anteriormente. Isso resultou na proliferação de géneros monotípicos. Alwyn Howard Gentry reduziu o número de géneros em 1973, 1976 e 1979.[14] No entanto, a revisão de 2004 descreveu 104 géneros, 38 dos quais monotípicos.[7]
Este problema era especialmente grave na tribo Bignonieae. Nessa tribo, muitas espécies de afinidade incerta foram atribuídas a um género vagamente definido, Arrabidaea, transformando esse género num «cesto de lixo» com cerca de 100 espécies.[14]
Desde 2004, estudos de filogenia molecular têm demonstrado que é necessária uma revisão substancial dos géneros. Muito do trabalho realizado com esse objetivo pode ser visto online,[9] mas pouco dele foi publicado em artigos científicos.
Uma história taxonómica detalhada da família Bignoniaceae foi publicada em 1980.[12] Um resumo daquela história foi publicado em 1999.[27]
Sistemática
No sistema APG IV de classificação das plantas com flor, a família Bignoniaceae é uma das 26 famílias da ordem Lamiales (a ordem Lamiales teria 27 famílias se a família Rehmanniaceae fosse aceite). Dentro da ordem, Bignoniaceae está num clado de oito famílias que consiste em Thomandersiaceae, Pedaliaceae, Martyniaceae, Schlegeliaceae, Bignoniaceae, Verbenaceae, Acanthaceae e Lentibulariaceae.[28] Este grupo é descrito como uma politomia, o que significa que não se conhecem dois dos seus membros que sejam mais próximos entre si do que qualquer um dos outros.[3] O suporte estatístico (reamostragem) para este grupo continua fraco, indicando que foram aplicados dados insuficientes ou que o grupo é um artefacto de algum método matemático de optimização usado na detrminação da sua filogenética.
A circunscrição taxonómica das Bignoniaceae tem sido relativamente estável e não sofreu nenhuma alteração no século XXI.[29] No século XX, as únicas questões relativas à circunscrição limitaram-se à determinação se as famílias Paulowniaceae e Schlegeliaceae deveriam ser fundidas na família Bignoniaceae ou aceites como famílias separadas.[6] A família Paulowniaceae era então composta por um a quatro géneros: Paulownia, Shiuyinghua, Wightia e Brandisia.[8] Independentemente da sua circunscrição, sabe-se agora que as Paulowniaceae estão mais próximas das Phrymaceae e das Orobanchaceae do que das Bignoniaceae. A família Schlegeliaceae foi incluída nas Bignoniaceae, como tribo Schlegelieae, ainda em 1980.[12] Atualmente, é aceite como uma família distinta, mas as suas relações com várias outras famílias permanecem por resolver.[5]
Em análises filogenéticas moleculares, a família Bignoniaceae apresenta um suporte bootstrap surpreendentemente fraco, dada a sua coerência morfológica. A tribo Jacarandeae (Digomphia e Jacaranda) é um grupo irmão do resto da família, conhecido como «Bignoniaceae nucelares» (em inglês: Core Bignoniaceae). O clado «Bignoniaceae nucleares» é fortemente apoiada em todas as análises filogenéticas moleculares, mas não tem nenhuma sinapomorfia morfológica conhecida.[6]
Nenhuma subfamília foi proposta para as Bignoniaceae em taxonomias recentes, mas em 2004, Fischer et al. dividiram a família em sete tribos: Tourrettieae, Eccremocarpeae, Tecomeae (sensu lato), Bignonieae, Oroxyleae, Crescentieae e Coleeae.[7] Desde então, Tourrettieae e Eccremocarpeae foram fundidas sob o nome Tourrettieae.[6] Tecomeae sensu lato revelou-se polifilético, consistindo nos seguintes grupos: Astianthus, Jacarandeae, Argylia, Delostoma, Perianthomega, Catalpeae, Tecomeae sensu stricto e toda a Crescentiina, exceto os géneros colocados em Crescentieae ou Coleeae. Todos estes grupos são monofiléticos, exceto Crescentiina pro parte. Contudo, Crescentiina é monofilética quando vista na sua totalidade.
Crescentiina é um tipo de nome sem nível taxonómico definido.[30] Crescentiina é composta por dois clados fortemente apoiados, informalmente denominados «Aliança Tabebuia» e «Clado paleotropical». A tribo Crescentieae está incorporada na «Aliança Tabebuia» e pode ser expandida para incluir Spirotecoma.[19] Coleeae sensu Fischer et al. (2004) é polifilético devido à inclusão de Kigelia e está aninhado dentro do «Clado paleotropical».[31] Perianthomega foi transferida de Tecomeae sensu stricto para Bignonieae, onde é o grupo irmão do restante da tribo.[14] Assim, as Bignoniaceae podem ser divididas em 10 grupos monofiléticos.
Filogenia
A árvore filogenética apresentada abaixo baseia-se nos resultados de quatro estudos filogenéticos.[6][14][19][31] Para todos os clados, a probabilidade posterior é de pelo menos 0,95 e o suporte bootstrap é de pelo menos 70%, exceto quando indicado de outra forma.
| Bignoniaceae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Caracteres evolutivos são apresentados são a presença de tricomas não glandulares, tricomas glandulares peltados, tricomas glandulares estipados e tricomas glandulares pateliformes ou cupulares. Três destes tricomas – tricomas não glandulares, tricomas glandulares peltados e tricomas glandulares pateliformes ou cupulares –, provavelmente, já estavam presentes em indivíduos ancestrais, enquanto os tricomas glandulares estipados surgiram mais recentemente e diversas vezes ao longo da evolução.
A família Bignoniaceae é classificada como um grupo monofilético, por apresentar as seguintes sinapomorfias: folhas transformadas em gavinhas (hábito liano) e crescimento secundário anómalo, Sendo esta última característica considerada uma característica derivada. Além disso, estudos moleculares de sequências dos genes rbcL e ndhF, comprovam que esta família pertence a um grupo monofilético. Nesse contexto, a presença de folhas pinadas compostas, na família Bignoniaceae, é considerada uma sinapomorfia deste grupo. Contudo algumas espécies dos géneros Crescentia, Tabebuia e Catalpa apresentam folhas reduzidas, trifoliadas e, às vezes, até unifoliadas. Para além disso, somente espécies dos géneros Crescentia, Parmentiera e Kigelia produzem frutos grandes e indeiscentes.
Géneros
Na última revisão taxonómica das Bignoniaceae, 104 géneros foram descritos em The Families and Genera of Vascular Plants (As Famílias e Géneros das Plantas Vasculares). Vinte e cinco desses géneros, todos da tribo Bignonieae, foram posteriormente sinonimizados sob outros géneros, com base numa análise cladística de sequências de ADN, publicada em 2006.[14] Os géneros Roseodendron e Handroanthus foram ressuscitados (repristinados) de Tabebuia em 2007.[19][32] Mayodendron e Pachyptera também foram repristinados.[9]
Em 2009, um estudo filogenético dividiu as Bignoniaceae em 10 grupos monofiléticos, conforme mostrado na lista de géneros abaixo. Seis desses grupos foram reconhecidos como tribos em algum momento e são representados pelos seus nomes tribais. Dois dos grupos são monogenéricos e são designados pelos seus géneros constituintes, Argylia e Delostoma. Os outros dois grupos receberam nomes informais, enquanto se aguarda uma revisão formal da classificação infra-familiar.
O género Astianthus nunca foi amostrada para ADN e a sua posição sistemática dentro da família permanece obscura. Da mesma forma, a colocação de Romeroa na aliança Tabebuia e a colocação de Sphingiphila em Bignonieae estão em dúvida.
O género Tecomaria não está incluída na lista abaixo, e o seu reconhecimento é controverso. É um táxon monotípico (Tecomaria capensis), e foi aceite durante muito tempo, mas foi devolvida ao género Tecoma em 1980.[12] Um estudo filogenético molecular resolveu-o como táxom irmão de outro género da África do Sul, Podranea, mas com apenas um fraco apoio bootstrap.[6] Tecomaria ainda não foi ressuscitado ou transferido para outro género.[9]
A tribo Bignonieae tem sido objeto de uma revisão considerável desde 2006. Fischer et al. colocou 46 géneros nesta tribo.[7] Posteriormente, Perianthomega foi transferido para o agrupamento a partir de Tecomeae sensu lato[14] e Pachyptera foi ressuscitado de Mansoa. Vinte e cinco dos géneros de Fischer foram incluídos noutros géneros, como se segue: Gardnerodoxa em Neojobertia; Memora em Adenocalymma; Leucocalantha em Pachyptera; Pseudocatalpa, Paragonia, Periarrabidaea, Spathicalyx, e Ceratophytum em Tanaecium; Arrabidaea e Piriadacus em Fridericia; Clytostoma, Cydista, Macranthisiphon, Mussatia, Phryganocydia, Potamoganos, Roentgenia e Saritaea em Bignonia; também Distictis, Glaziovia, Haplolophium e Pithecoctenium em Amphilophium. Assim, atualmente são reconhecidos 23 géneros em Bignonieae.[9]
- Incertae sedis
- Astianthus
- Tribo Jacarandeae
- Jacaranda (sin.: Digomphia)
- Tribo Tourrettieae
- Eccremocarpus
- Tourrettia
- Género Argylia
- Tribo Tecomeae
- Campsidium
- Campsis
- Deplanchea
- Dinklageodoxa
- Incarvillea
- Lamiodendron
- Neosepicaea
- Pandorea
- Podranea
- Tecoma
- Tecomanthe
- Género Delostoma
- Tribo Bignonieae
- Adenocalymma
- Amphilophium (sin.: Distictella)
- Anemopaegma
- Bignonia
- Callichlamys
- Cuspidaria
- Dolichandra
- Fridericia
- Lundia
- Manaosella
- Mansoa
- Martinella
- Neojobertia
- Pachyptera
- Perianthomega
- Pleonotoma
- Pyrostegia
- Stizophyllum
- Tanaecium (sin.: Sphingiphila)
- Tynanthus
- Xylophragma
- Tribo Oroxyleae
- Hieris
- Millingtonia
- Nyctocalos
- Oroxylum
- Tribo Catalpeae
- Chilopsis
- Catalpa
- Grupo Crescentiina
- Aliança Tabebuia
- Amphitecna
- Crescentia
- Cybistax
- Ekmanianthe
- Godmania
- Handroanthus
- Paratecoma
- Parmentiera
- Romeroa
- Roseodendron
- Sparattosperma
- Spirotecoma
- Tabebuia
- Zeyheria
- Clado paleotropical
- Catophractes
- Colea (sin.: Ophiocolea)
- Dolichandrone
- Fernandoa (sin.: Haplophragma)
- Heterophragma
- Kigelia
- Markhamia
- Mayodendron
- Newbouldia
- Pajanelia
- Pauldopia
- Perichlaena
- Phyllarthron
- Phylloctenium
- Radermachera
- Rhigozum
- Rhodocolea
- Tecomella
- Santisukia
- Spathodea
- Stereospermum
- Aliança Tabebuia
Géneros obsoletos
- Arrabidaea
- Ceratophytum
- Clytostoma
- Cydista
- Distictis
- Gardnerodoxa
- Glaziovia
- Haplolophium
- Leucocalantha
- Macfadyena
- Macranthisiphon
- Melloa
- Memora
- Mussatia
- Parabignonia
- Paragonia
- Periarrabidaea
- Phryganocydia
- Piriadacus
- Pithecoctenium
- Potamoganos
- Pseudocatalpa
- Roentgenia
- Saritaea
- Spathicalyx
Tribos e sua distribuição geográfica
A família Bignoniaceae apresenta a seguinte distribuição natural:
- Tribo Bignonieae Dum.: De acordo com Lohmann et al. 2014, contém cerca de 21 géneros com aproximadamente 393 espécies:[33]
- Adenocalymma Mart. ex Meisn. nom. cons. (sin.: Memora Miers, Odontotecoma Bureau & K.Schum., Pharseophora Miers, Sampaiella J.C.Gomes): As 81[33] a 82 espécies estão amplamente distribuídas na região neotropical, desde o México até à América do Sul tropical.[34][35]
- Amphilophium Kunth[33] (sin.: Anomoctenium Pichon, Bothriopodium Rizzini, Distictella Kuntze, Distictis Mart. ex Meisn., Distictis Bureau nom. illeg., Endoloma Raf., Glaziova Bureau, Haplolophium Cham., Leiogyne K.Schum., Macrodiscus Bureau, Neves-armondia K.Schum., Phaedranthus Miers, Pithecoctenium Mart. ex DC., Sererea Raf., Urbanolophium Melch., Wunschmannia Urb.): As cerca de 47 espécies estão amplamente distribuídas na região neotropical.[34] Entre as quais:
- Amphilophium buccinatorium (DC.) L.G.Lohmann: Do México.
- Anemopaegma Mart. ex Meisn.[33] (sin.: Cupulissa Raf., Platolaria Raf., Pseudopaegma Urb.): As cerca de 45 espécies estão amplamente distribuídas desde o México até à América Central e à América do Sul tropical, e são encontradas na ilha de Trinidad.[34]
- Bignonia L.[33] (sin.: Anisostichus Bureau nom. superfl., Clytostoma Miers ex Bureau, Clytostomanthus Pichon, Cydista Miers, Levya Bureau ex Baill., Macranthisiphon Bureau ex K.Schum., Micropaegma Pichon, Mussatia Bureau ex Baill., Osmhydrophora Barb.Rodr., Phryganocydia Mart. ex Bureau, Phrygiobureaua Kuntze nom. superfl., Pongelia Raf., Potamoganos Sandwith, Roentgenia Urb., Saritaea Dugand, Temnocydia Mart. ex DC. nom. superfl.): As 28 a 31 espécies[33] estão amplamente distribuídas desde o centro e leste dos Estados Unidos até a América Central, ilhas das Caraíbas e Argentina.[34] Entre as quais:
- Bignonia capreolata L.
- Callichlamys Miq.: Contém apenas uma espécie:[33]
- Callichlamys latifolia (Rich.) K.Schum.: Está presente desde o sul do México, passando pela América Central, até à América do Sul tropical.[34]
- Cuspidaria DC. nom. cons. (sin.: Blepharitheca Pichon, Cremastus Miers, Lochmocydia Mart. ex DC., Nouletia Endl., Saldanhaea Bureau nom. illeg., Setilobus Baill., Tetrastichella Pichon): As cerca de 19 espécies[33] estão distribuídas desde o sul do México até à América do Sul tropical e ocorrem na ilha de Trinidad.[34]
- Dolichandra Cham.: As cerca de nove espécies estão distribuídas desde o México até à América do Sul tropical.[34]
- Fridericia Mart.: As cerca de 67 espécies[33] estão distribuídas desde o México até à América do Sul tropical.[34]
- Lundia DC.: As 13[33] a 14 espécies estão distribuídas desde o México até à América do Sul tropical e ocorrem na ilha de Trinidad.[34]
- Manaosella J.C.Gomes: Contém apenas uma espécie:[33]
- Mansoa DC.: As 12[33] a 18 espécies estão distribuídas desde o México até à América do Sul tropical.[34]
- Mansoa alliacea (Lam.) A.H.Gentry: Do norte da América do Sul.
- Martinella Baill.: As apenas três espécies estão distribuídas desde o México até à América do Sul tropical.[34]
- Neojobertia Baill.: As duas espécies[33] ocorrem no leste do Brasil.[34]
- Pachyptera DC. ex Meisn.: As cerca de quatro espécies estão distribuídas na região neotropical.[34]
- Perianthomega Bureau ex Baill.: Contém apenas uma espécie:[33]
- Perianthomega vellozoi Bureau: Ocorre desde a Bolívia até o Brasil.[34]
- Pleonotoma Miers: As cerca de 17 espécies 17[33] estão distribuídas desde a América Central até à América do Sul tropical e ocorrem na ilha de Trinidad.[34]
- Pyrostegia C.Presl: As apenas duas espécies estão distribuídas desde o México até à América do Sul tropical.[34]
- Pyrostegia venusta (Ker Gawl.) Miers: Da América Central até ao oeste e sul do México, bem como do norte da América do Sul até à Argentina.
- Stizophyllum Miers: As cerca de três espécies estão distribuídas desde o sul do México até à América do Sul tropical.[34]
- Tanaecium Sw. (sin.: Sphingiphila A.H.Gentry): As cerca de 17 espécies estão distribuídas na região neotropical.[34]
- Tynanthus Miers: As cerca de 15 espécies estão distribuídas desde o sul do México até à América do Sul tropical.[34]
- Tynanthus panurensis (Bureau) Sandwith: Do noroeste da América do Sul, Brasil, Equador, Peru e Bolívia.
- Xylophragma Sprague: As cerca de sete espécies estão distribuídas desde o sul do México até à América do Sul tropical e ocorrem na ilha de Trinidad.[34]
- Tribo Catalpeae DC. ex Meisn.: Foi reativada em 2009. Contém apenas dois (ou três) géneros com cerca de onze espécies:[36]
- Catalpa Scop. s. l. (sin.: Cumbalu Adans., Catalpium Raf., Macrocatalpa Britton): Das cerca de dez espécies, duas ocorrem no sudoeste dos Estados Unidos, quatro nas Grandes Antilhas e quatro na Ásia Oriental (na China[34]).[36]
- Chilopsis D.Don: Contém apenas uma espécie:
- Chilopsis linearis (Cav.) Sweet: Ocorre com duas subespécies e duas variedades, desde o sul dos EUA até ao norte do México.[34]
- Tribo Coleeae Bojer ex Reveal: De acordo com Olmstead 2009, ela contém cerca de quatro géneros com aproximadamente 60 espécies,[36] de acordo com Fischer 2012[37] um género monotípico a mais e, segundo Callmander 2015, novamente apenas quatro géneros.[38] Os quatro géneros com cerca de 35 espécies existentes desde 2015 ocorrem em Madagáscar e nas ilhas do Oceano Índico ocidental.[34][38]
- Colea Bojer ex Meisn. (sin.: Odisca Raf. nom. rej., Ophiocolea H.Perrier, Uloma Raf. nom. rej.). Com cerca de 35 espécies, que ocorrem em ilhas no Oceano Índico ocidental.[34]
- Phyllarthron DC. (sin.: Phylloctenium Baill., Zaa Baill.): É monofilética apenas se incluir o género Phylloctenium.[36] As cerca de 20 espécies ocorrem em Madagáscar e nas Comores.[34]
- Phylloctenium Baill. (sin.: Arthrophyllum Bojer ex DC. nom. illeg. non Arthrophyllum Blume, Paracolea Baill.): As duas espécies existentes só ocorrem em Madagáscar.[34]
- Rhodocolea Baill.: As cerca de 16 espécies existentes desde 2011 só ocorrem em Madagáscar.[34]
- Tribo Crescentieae G.Don: Contém apenas três géneros com cerca de 35 espécies:[36]
- Amphitecna Miers (sin.: Dendrosicus Raf., Enallagma (Miers) Baill., Neotuerckheimia Donn.Sm.): As cerca de 20 espécies existentes desde 2016 estão distribuídas na região neotropical, desde o sul da Flórida até ao Equador, passando pela América Central e pelas Ilhas das Caraíbas.[34] Muitas espécies são endémicas. No México, existem cerca de dez espécies.[39]
- Crescentia L. (sin.: Cuiete Mill., Daubentona Buc'hoz, Pteromischus Pichon): As cerca de seis espécies estão distribuídas desde o México até à América do Sul tropical.[34] Darunter:
- Crescentia alata Kunth: Do México e da América Central.
- Crescentia cujete L.: Da Venezuela, Colômbia, Caribe, América Central e México.
- Parmentiera DC. (sin.: Zenkeria Rchb.): As cerca de dez espécies estão distribuídas desde o sul do México até à América Central, passando pela Colômbia e pelo Uruguai.[34]
- Parmentiera cereifera Seem.: Da América Central até às Caraíbas.
- Tribo Jacarandeae Fenzl: Foi reativada em 2009. Contém apenas dois géneros com 50 a 55 espécies:
- Digomphia Benth.: As cerca de três espécies são comuns na América do Sul tropical.[34]
- Jacaranda Juss. (sin.: Etorloba Raf., Icaranda Pers. orth. var., Kordelestris H.Kost., Pteropodium DC., Rafinesquia Raf.): As cerca de 47 espécies estão amplamente distribuídas desde a América Central tropical e as ilhas das Caraíbas até ao norte da Argentina.[34]
- Tribo Oroxyleae A.H.Gentry ex Reveal & L.G.Lohmann: Contém cerca de quatro géneros com cerca de seis espécies:[36]
- Hieris Steenis: Contém apenas uma espécie:
- Millingtonia L.f.: Contém apenas uma espécie:
- Nyctocalos Teijsm. & Binn.: As cerca de três espécies estão distribuídas desde o centro-sul da China até à Indochina e à Malésia.[34]
- Oroxylum Vent.: Contém apenas uma espécie:
- Oroxylum indicum (L.) Benth. ex Kurz: Ocorre desde a Malésia até ao sul da China.[34]
- Tribo Tecomeae Endl.: Em Olmstead 2009[36] contém apenas cerca de 12 géneros com cerca de 55 espécies, mas em Fischer 2012 contém 43 géneros:[37]
- Argylia D.Don (sin.: Oxymitus C.Presl): As cerca de 13 espécies estão distribuídas desde o sul do Peru até ao Chile e à Argentina.[34]
- Astianthus D.Don: Contém apenas uma espécie:[34]
- Campsidium Seem.: Contém apenas uma espécie:
- Campsidium valdivianum (Phil.) W.Bull: Ocorre desde o sul do Chile até o sudoeste da Argentina.[34]
- Campsis Lour.: Das duas espécies existentes, uma ocorre desde o leste da China até ao Japão e a outra na região central e leste dos EUA.[34] As variedades são utilizadas como plantas ornamentais.
- Catophractes D.Don: Contém apenas uma espécie:
- Cybistax Mart. ex Meisn.: Contém apenas uma espécie:
- Cybistax antisyphilitica (Mart.) Mart.: Ocorre na América do Sul tropical.[34]
- Delostoma D.Don (sin.: Codazzia H.Karst. & Trian): As cerca de quatro espécies estão distribuídas desde a Colômbia, oeste da Venezuela até ao Equador e Peru.[34]
- Deplanchea Vieill. (sin.: Bulweria F.Muell., Diplanthera Banks & Sol. ex R.Br., Montravelia Montrouz. ex Beauvis.): As cerca de oito espécies estão distribuídas desde a Malésia até à Austrália e Nova Caledónia.[34]
- Dinklageodoxa Heine & Sandwith: Contém apenas uma espécie:
- Dolichandrone (Fenzl) Seem.: As cerca de dez espécies estão distribuídas desde a Ásia tropical até às ilhas do sudoeste do Pacífico e em Moçambique.[34]
- Ekmanianthe Urb.: As duas únicas espécies ocorrem em Cuba e Hispaniola.[34]
- Fernandoa Welw. ex Seem. (sin.: Haplophragma Dop, Hexaneurocarpon Dop, Kigelianthe Baill., Spathodeopsis Dop, Tisserantodendron Sillans): As cerca de 15 espécies estão distribuídas desde a África tropical até à Ásia e à Malásia ocidental.[34]
- Godmania Hemsl. (sin.: Xerotecoma J.C.Gomes): As duas espécies estão distribuídas desde o México, passando pela América Central, até à América do Sul tropical.[34]
- Handroanthus Mattos: As cerca de 33 espécies estão distribuídas desde o México, passando pela América Central e pelas ilhas das Caraíbas, até à América do Sul tropical.[34] Entre as quais:
- Handroanthus chrysotrichus (Mart. ex DC.) Mattos: Do nordeste da Argentina e do leste ao sudeste do Brasil.
- Handroanthus impetiginosus (Mart. ex DC.) Mattos: Desde o norte da América do Sul central e América Central até ao sul do México.
- Heterophragma DC.: As duas espécies estão distribuídas desde a Índia até à Indonésia.[34]
- Incarvillea Juss.: As cerca de 17 espécies estão amplamente distribuídas desde a Ásia Central até ao Himalaia e à China.[34]
- Incarvillea semiretschenskia Grierson: Ocorre na Ásia Central, no Cazaquistão.
- Kigelia DC. (sin.: Kigelkeia Raf. orth. var., Sotor Fenzl): Contém apenas uma espécie:
- Kigelia africana (Lam.) Benth.: Está presente desde as regiões tropicais da África até à África Austral.[34] É possível distinguir duas subespécies.[34]
- Lamiodendron Steenis: Contém apenas uma espécie:
- Lamiodendron magnificum Steenis: Ela ocorre na parte oriental da Nova Guiné.[34]
- Markhamia Seem. ex Baill.: As cinco espécies estão distribuídas desde a África tropical até à África Austral e desde a Indochina até ao sul da China.[34]
- Mayodendron Kurz: Contém apenas uma espécie:
- Neosepicaea Diels: As quatro espécies estão distribuídas desde a Nova Guiné até Queensland.[34]
- Newbouldia Seem.: Contém apenas uma espécie:
- Newbouldia laevis (P.Beauv.) Seem.: Ocorre na região tropical da África Ocidental.[34]
- Pajanelia DC.: Contém apenas uma espécie:
- Pajanelia longifolia (Willd.) K.Schum.: Ocorre desde o sul da Índia até ao oeste da Malásia.[34]
- Pandorea Spach: As sete[36] a nove espécies estão distribuídas desde a Indonésia até à Austrália e Nova Caledónia.[34]
- Pandorea jasminoides (Lindl.) K.Schum.
- Paratecoma Kuhlm.: Contém apenas uma espécie:
- Paratecoma peroba (Record) Kuhlm.: Ocorre no Brasil.[34]
- Pauldopia Steenis: Contém apenas uma espécie:
- Pauldopia ghonta (Buch.-Ham. ex G.Don) Steenis: Ocorre desde a Índia até à Indochina.[34]
- Perichlaena Baill.: Contém apenas uma espécie:
- Perichlaena richardii Baill.: Apenas encontrada no norte de Madagáscar.[34]
- Podranea Sprague: As duas únicas espécies estão distribuídas desde o sul da África tropical até ao sul da África.[34][36]
- Radermachera Zoll. & Moritzi: As cerca de 17 espécies estão distribuídas desde a Ásia tropical até ao sul da China.[34]
- Rhigozum Burch.: As sete espécies estão distribuídas desde a África tropical até à África Austral, em Madagáscar e da Somália ao Iémen.[34]
- Romeroa Dugand: Contém apenas uma espécie:
- Romeroa verticillata Dugand: Ocorre na Colômbia.[34]
- Roseodendron Miranda: As duas espécies estão distribuídas desde o México até à Colômbia.[34]
- Santisukia Brummitt: As duas espécies só ocorrem na Tailândia.[34]
- Sparattosperma Mart. ex Meisn.: As duas únicas espécies ocorrem na América do Sul tropical.[34]
- Spathodea P.Beauv.: Contém apenas uma espécie:
- Spathodea campanulata P.Beauv.: As três subespécies ocorrem desde a África Ocidental tropical até ao Uganda e a Angola.[34]
- Spirotecoma (Baill.) Dalla Torre & Harms: As cerca de quatro espécies ocorrem em Cuba e Hispaniola.[34]
- Stereospermum Cham.: As cerca de 25 espécies estão amplamente distribuídas no Velho Mundo, na África tropical, em Madagáscar e desde o subcontinente indiano até à Malásia ocidental e ao sul da China.[34]
- Tabebuia Gomes ex DC.: As cerca de 74 espécies estão amplamente distribuídas na região neotropical.[34]
- Tecoma Juss.: As 9[34] a 14[36] espécies ocorrem desde a Tanzânia até à África Austral e na região neotropical.[34]
- Tecomanthe Baill.: As cerca de cinco espécies ocorrem desde Maluku até às ilhas Salomão, em Queensland e na Nova Zelândia.[34]
- Tecomella Seem.: Contém apenas uma espécie:
- Zeyheria Mart.: As duas únicas espécies estão distribuídas desde a Bolívia até ao Brasil.[34]
- Tribo Tourrettieae G.Don: Continha apenas um género e, desde 2009, também contém o único género da antiga tribo Eccremocarpeae Hogg: Os dois géneros contêm apenas cerca de quatro espécies:[36]
Distribuição no Brasil
Espécies desta família possui distribuição predominantemente tropical, principalmente na América do Sul. O Brasil apresenta 117 espécies, sendo considerado o centro de diversidade desta família, com 32 gêneros e 391 espécies. Ocorre em quase todos os estados do Brasil, distribuindo-se de Norte a Sul. Esta família não é endêmica do Brasil, ou seja, ainda existem alguns gêneros com ocorrência em regiões temperadas.
O seguintes estados brasileiros têm ocorrências confirmadas:
- Norte (Acre, Amazonas, Amapá, Pará, Rondônia, Roraima, Tocantins);
- Nordeste (Alagoas, Bahia, Ceará, Maranhão, Paraíba, Pernambuco, Piauí, Rio Grande do Norte, Sergipe);
- Centro-Oeste (Distrito Federal, Goiás, Mato Grosso do Sul, Mato Grosso);
- Sudeste (Espírito Santo, Minas Gerais, Rio de Janeiro, São Paulo);
- Sul (Paraná, Rio Grande do Sul, Santa Catarina).
O seguintes estados brasileiros têm possíveis ocorrências:
- Norte (Acre, Amazonas, Pará, Rondônia);
- Nordeste (Bahia, Paraíba);
- Centro-Oeste (Distrito Federal, Goiás, Mato Grosso do Sul, Mato Grosso);
- Sudeste (Espírito Santo, Minas Gerais, Rio de Janeiro, São Paulo);
- Sul (Santa Catarina).
A família ocorre nos seguintes domínios fitogeográficos:
- Amazônia, Caatinga, Cerrado, Mata Atlântica, Pampa, Pantanal.
As espécies pertencentes à família Bignoniaceae estão associadas aos seguintes tipos de vegetação:
- Área Antrópica, Caatinga (stricto sensu), Campinarana, Campo de Altitude, Campo de Várzea, Campo Limpo, Campo Rupestre, Carrasco, Cerrado (lato sensu), Floresta Ciliar ou Galeria, Floresta de Igapó, Floresta de Terra Firme, Floresta de Várzea, Floresta Estacional Decidual, Floresta Estacional Perenifólia, Floresta Estacional Semidecidual, Floresta Ombrófila (= Floresta Pluvial), Floresta Ombrófila Mista, Manguezal, Restinga, Savana Amazônica, Vegetação Sobre Afloramentos Rochosos.
Conservação no Brasil
Segundo a lista oficial das espécies da flora brasileira ameaçadas de extinção, a família Bignoniaceae apresenta 10 espécies ameaçadas de extinção, sendo elas:
- Adenocalymma magnoalatum
- Adenocalymma ubatubense
- Digomphia densicoma
- Jacaranda carajasensis
- Jacaranda crassifolia
- Jacaranda intricata
- Jacaranda rugosa
- Jacaranda subalpina
- Tabebuia botelhensis
- Tabebuia selachidentata.
Listas de gêneros brasileiros
- Adenocalymma Mart. ex Meisn. emend L.G. Lohmann
- Amphilophium Kunth
- Anemopaegma Mart. ex Meisn.
- Bignonia L., Callichlamys Miq.
- Crescentia L.
- Cuspidaria DC.
- Cybistax Mart. ex Meisn.
- Digomphia Benth.
- Dolichandra Cham.
- Fridericia Mart.
- Godmania Hemsl.
- Handroanthus Mattos
- Jacaranda Juss.
- Lundia DC.
- Manaosella J.C. Gomes
- Mansoa DC.
- Martinella Baill.
- Neojobertia Baill.
- Pachyptera DC., ex Meisn.
- Paratecoma Kuhlm.
- Perianthomega Bureau ex Baill.
- Pleonotoma Miers
- Pyrostegia C. Presl
- Sparattosperma Mart. ex Meisner
- Spathodea P. Beauv.
- Stizophyllum Miers
- Tabebuia Gomes ex DC.
- Tanaecium Sw. emend L.G. Lohmann
- Tecoma Juss.
- Tynanthus Miers
- Xylophragma Sprague
- Zeyheria Mart.
Referências
- ↑ Wilf, Peter (fevereiro 2000). «Late Paleocene-early Eocene climate changes in southwestern Wyoming: Paleobotanical analysis». Geological Society of America Bulletin. 112 (2): 292–307. Bibcode:2000GSAB..112..292W. doi:10.1130/0016-7606(2000)112<0292:LPEECC>2.3.CO;2. Consultado em 15 Maio 2020
- ↑ Angiosperm Phylogeny Group (2009), «An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III», Botanical Journal of the Linnean Society, 161 (2): 105–121, doi:10.1111/j.1095-8339.2009.00996.x
 , hdl:10654/18083
, hdl:10654/18083
- ↑ a b Peter F. Stevens (2001 onwards). "Bignoniaceae" At: Angiosperm Phylogeny Website. At: Botanical Databases At: Missouri Botanical Garden Website. (ver «Links externos» abaixo).
- ↑ «Bignoniaceae». Merriam-Webster Dictionary. Consultado em 11 de junho de 2023
- ↑ a b Vernon H. Heywood, Richard K. Brummitt, Ole Seberg, and Alastair Culham. Flowering Plant Families of the World. Firefly Books: Ontario, Canada. (2007). ISBN 978-1-55407-206-4.
- ↑ a b c d e f Olmstead, Richard G.; Zjhra, Michelle L.; Lohmann, Lúcia G.; Grose, Susan O.; Eckert, Andrew J. (2009). «A molecular phylogeny and classification of Bignoniaceae». American Journal of Botany. 96 (9): 1731–1743. PMID 21622359. doi:10.3732/ajb.0900004
- ↑ a b c d e f g h i j k l m n o p q Eberhard Fischer, Inge Theisen & Lúcia G. Lohmann. 2004. "Bignoniaceae". pages 9-38. In: Klaus Kubitzki (editor) and Joachim W. Kadereit (volume editor). The Families and Genera of Vascular Plants volume VII. Springer-Verlag: Berlin; Heidelberg, Germany. ISBN 978-3-540-40593-1
- ↑ a b David J. Mabberley. 2008. Mabberley's Plant-Book third edition (2008). Cambridge University Press: UK. ISBN 978-0-521-82071-4
- ↑ a b c d e Lúcia G. Lohmann and Carmen U. Ulloa. 2007 onward. Bignoniaceae in iPlants prototype Checklist (veja «Links externos» abaixo).
- ↑ a b George W. Staples and Derral R. Herbst. 2005. "A Tropical Garden Flora" Bishop Museum Press: Honolulu, HI, USA. ISBN 978-1-58178-039-0
- ↑ a b Anthony Huxley, Mark Griffiths & Margot Levy (1992). The New Royal Horticultural Society Dictionary of Gardening. The Macmillan Press, Limited: London. The Stockton Press: New York. ISBN 978-0-333-47494-5 (set).
- ↑ a b c d e Gentry, Alwyn H. (1980). «"Bignoniaceae: Part I (Crescentieae and Tourrettieae)". Flora Neotropica». Monograph. 25 (1): 1–130. JSTOR 4393736
- ↑ a b c d e f g h i Gentry, Alwyn H. (1992). «A Synopsis of Bignoniaceae Ethnobotany and Economic Botany». Annals of the Missouri Botanical Garden. 79 (1): 53–64. JSTOR 2399809. doi:10.2307/2399809
- ↑ a b c d e f g Lohmann, Lúcia G. (2006). «Untangling the phylogeny of neotropical lianas (Bignonieae, Bignoniaceae)». American Journal of Botany. 93 (2): 304–318. PMID 21646191. doi:10.3732/ajb.93.2.304
- ↑ Pace, Marcelo R.; Lohmann, Lúcia G.; Angyalossy, Veronica (2011). «Evolution of disparity between the regular and variant phloem in Bignonieae (Bignoniaceae)». American Journal of Botany. 98 (4): 602–618. PMID 21613161. doi:10.3732/ajb.1000269
- ↑ Goldblatt, Peter; Gentry, Alwyn H. (1979). «Cytology of Bignoniaceae». Botaniska Notiser. 132 (4): 475–482
- ↑ Robert Hegnauer. 1989. Chemotaxonomie der Pflanzen 8:128–138. Birkhäuser Verlag: Basel, Switzerland; Boston MA, USA; Berlin, Germany. ISBN 978-3-7643-1895-6
- ↑ von Poser, Gilsane Lino; Schripsema, Jan; Henriques, Amélia T.; Rosendal Jensen, Soren (2000). «The distribution of iridoids in Bignoniaceae». Biochemical Systematics and Ecology. 28 (4): 351–366. Bibcode:2000BioSE..28..351L. PMID 10725592. doi:10.1016/s0305-1978(99)00076-9
- ↑ a b c d Susan O. Grose; Richard G. Olmstead (2007), «Evolution of a Charismatic Neotropical Clade: Molecular Phylogeny of Tabebuia s.l., Crescentieae, and Allied Genera (Bignoniaceae)», Systematic Botany, 32 (3): 650–659, JSTOR 25064274, doi:10.1600/036364407782250553
- ↑ a b Record, Samuel J.; Hess, Robert W. (1940). «American timbers of the family Bignoniaceae». Tropical Woods. 63: 9–38
- ↑ Christian Rätsch. 2005. The Encyclopedia of Psychoactive Plants (translated by John R. Baker). Park Street Press: Rochester VT, USA. ISBN 978-0-89281-978-2.
- ↑ Gentry, Alwyn H.; Cook, Kathleen (1984). «Martinella (Bignoniaceae): a widely used eye medicine if South America». Journal of Ethnopharmacology. 11 (3): 337–343. PMID 6482483. doi:10.1016/0378-8741(84)90079-5
- ↑ James L. Reveal. 2008on. "Bignoniaceae" In: A checklist of suprageneric names for extant vascular plants At: Home page of James L. Reveal & C. Rose Broome (veja «ligações externas» abaixo).
- ↑ Bignonia In: International Plant Names Index (ver «Ligaçoes externas» abaixo).
- ↑ Umberto Quattrocchi. 2000. CRC World Dictionary of Plant Names volume I. CRC Press: Boca Raton; New York; Washington, DC;, USA. London, UK. ISBN 978-0-8493-2675-2 (vol. I).
- ↑ George Bentham and Joseph D. Hooker. 1876. Genera plantarum :ad exemplaria imprimis in Herberiis Kewensibus servata definita vol. 2 part 2:1026-1053. Reeve & Co. London, England. (Veja «Ligações externas» abaixo).
- ↑ Russell E. Spangler and Richard G. Olmstead. 1999. "Phylogenetic Analysis of Bignoniaceae Based on the cpDNA Gene Sequences of rbcL and ndhF". Annals of the Missouri Botanical Garden 86(1):33-46. (ver Liagções externas abaixo).
- ↑ Schäferhoff, Bastian; Fleischmann, Andreas; Fischer, Eberhard; Albach, Dirk C.; Borsch, Thomas; Heubl, Günther; Müller, Kai F. (2010). «Towards resolving Lamiales relationships: insights from rapidly evolving chloroplast sequences». BMC Evolutionary Biology. 10 (1): 352. Bibcode:2010BMCEE..10..352S. PMC 2992528. PMID 21073690. doi:10.1186/1471-2148-10-352
- ↑ Reveal, James L. (2011). «Summary of recent systems of angiosperm classification». Kew Bulletin. 66 (1): 5–48. Bibcode:2011KewBu..66....5R. doi:10.1007/s12225-011-9259-y
- ↑ Kron, Kathleen A. (1997). «Exploring alternative systems of classification». Aliso. 15 (2): 105–112. doi:10.5642/aliso.19961502.05
- ↑ a b Zjhra, Michelle L.; Sytsma, Kenneth J.; Olmstead, Richard G. (2004). «Delimitation of Malagasy tribe Coleeae and implications for fruit evolution in Bignoniaceae inferred from a chloroplast DNA phylogeny». Plant Systematics and Evolution. 245 (1–2): 55–67. doi:10.1007/s00606-003-0025-y
- ↑ Susan O. Grose; Richard G. Olmstead (2007), «Taxonomic Revisions in the Polyphyletic Genus Tabebuia s.l. (Bignoniaceae)», Systematic Botany, 32 (3): 660–670, doi:10.1600/036364407782250652
- ↑ a b c d e f g h i j k l m n o Lúcia G. Lohmann, Charlotte M. Taylor: A New Generic Classification of Tribe Bignonieae (Bignoniaceae). In: Annals of the Missouri Botanical Garden, vol. 99, n.º 3, 2014, pp. 348–489. doi:10.3417/2003187.
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf bg bh bi bj bk bl bm bn bo bp bq br bs bt bu bv bw bx by bz ca cb cc cd ce cf Bignoniaceae. In: POWO = Plants of the World Online von Board of Trustees of the Royal Botanic Gardens, Kew: Kew Science
- ↑ R. Udulutsch, M. A. Assis, P. Dias: Taxonomic update of Adenocalymma (Bignoniaceae) emendations, new synonyms, typifications, and status change. In: Turkish Journal of Botany, Volume 37, 2013, pp. 630–643.
- ↑ a b c d e f g h i j k Richard G. Olmstead, Michelle L. Zjhra, Lúcia G. Lohmann, Susan O. Grose, Andrew J. Eckert: A molecular phylogeny and classification of Bignoniaceae. In: American Journal of Botany, Volume 96, Issue 9, 2009, pp. 1731–1743. doi:10.3732/ajb.0900004.
- ↑ a b E. Fischer, I. Theisen, L. G. Lohmann (2004). «Flowering Plants – Dicotyledons: Lamiales (except Acanthaceae including Avicenniaceae)». In: Klaus Kubitzki, Joachim W. Kadereit. Bignoniaceae. The Families and Genera of Vascular Plants. 7. [S.l.]: Springer Science & Business Media. p. 9-38. 9 páginas. ISBN 978-3-642-18617-2}}.
- ↑ a b Martin Wilhelm Callmander, Peter B. Phillipson, Gregory Michael Plunkett, Edwards, Sven Buerki: Generic delimitations, biogeography and evolution in the tribe Coleeae (Bignoniaceae), endemic to Madagascar and the smaller islands of the western Indian Ocean. In: Molecular Phylogenetics and Evolution , Volume 96, 2015, pp. 178–186, figs: 2. doi:10.1016/j.ympev.2015.11.016.
- ↑ Andres Ernesto Ortiz-Rodriguez, Carlos Manuel Burelo Ramos, Héctor Gomez-Dominguez: A new species of Amphitecna (Bignoniaceae) endemic to Chiapas, Mexico. In: PhytoKeys, Volume 65, June 2016, pp. 15–23. doi:10.3897/phytokeys.65.8454.
Bibliografia
- Alwyn H. Gentry. 1992. "Bignoniaceae: Part II (Tecomeae)". Flora Neotropica Monograph 25(2):1-150. (See External links below).
- Araújo Ricardo de Souza 2008,Bignoniaceae Juss. do Parque Estadual do Itacolomi, Minas Gerais, Brasil: florística, similaridade e distribuição geográfica.
- Bignoniaceae em FloraSBS.
- Bignoniaceae em Lista Oficial das Espécies da Flora Brasileira Ameaçadas de Extinção.
- Bignoniaceae in Flora do Brasil 2020 em construção. Jardim Botânico do Rio de Janeiro.
- Biologia floral e polinização de Arrabidaea conjugata (Vell.) Mart. (Bignoniaceae).
- Evolução da morfologia floral e estrutura de comunidades em um clado de Lianas Tropicais (Bignoniaceae, Bignoniaceae) em Biblioteca Digital de Teses e Dissertações da USP.
- Gentry, A. H. 1980. Bignoniaceae I (Crescentieae and Tourrettieae). Flora Neotropica Monograph 25(1): 1-130.
- José Magno das Chagas Junior, Douglas Antônio de Carvalho, Mariana Esteves Mansanares 2009 em A família Bignoniaceae Juss. (Ipês) no município de Lavras, Minas Gerais.
- Lima, A. C., Pace, M. R. & Angyalossy, V. 2010. Seasonality and growth rings in lianas of Bignoniaceae. Trees-Structure and Function 24(6): 1045-1060.
- Lohmann, L. G. (2006). Untangling the phylogeny of neotropical lianas (Bignonicae, Bignoniaceae). American Journal of Botany 93(2): 304-318.
- Lohmann, L.G. 2015. Bignoniaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro.
- BFG. Growing knowledge: an overview of Seed Plant diversity in Brazil. Rodriguésia, v. 66, n.º 4, pp.1085-1113. 2015 (DOI: 10.1590/2175-7860201566411).
- Maria Eugenia Carvalho do Amaral 1992 em Ecologia floral de dez espécies da tribo Bignonicae (Bignoniaceae), em uma floresta semidecídua no Município de Campinas, SP.
- Olmstead, R. G., M. L. Zjhra, Lohmann, L. G., Grose, S. O. & Eckert, A. J. (2009). A molecular phylogeny and classification of Bignoniaceae. American Journal of Botany 96(9): 1731-1743.
- Pace, M. R., L. G. Lohmann & Angyalossy, V. (2009). The rise and evolution of the cambial variant in Bignonieae (Bignoniaceae). Evolution & Development 11(5): 465-479.
- Pace, M. R., L. G. Lohmann & Angyalossy, V. (2011). Evolution of disparity between the regular and variant phloem in Bignonieae (Bignoniaceae). American Journal of Botany 98(4): 602-618.
- Veridiana Vizoni Scudeller1 em Bignoniaceae Juss. no Parque Nacional da Serra da Canastra – Minas Gerais, Brasil.
- Walter S. Judd; Christopher S. Campbell; Elizabeth A. Kellogg; Peter F. Stevens; Michael J. Donoghue. Sistemática Vegetal: um enfoque filogenético. Cap. 9. 3.ª ed., 2009.
Ligações externas
- Bignoniaceae in L. Watson and M. J. Dallwitz (1992 onwards), The families of flowering plants.
- Distribution Map[ligação inativa] And Genus List At: Bignoniaceae At: Lamiales At: Trees[ligação inativa] At: APweb At: botanical databases At: About Science & Conservation At: Missouri Botanical Garden
- Crescentieae and Tourrettieae At: Flora Neotropica 25(1) At: Flora Neotropica At: Organization for Flora Neotropica
- tribe Tecomeae At: Flora Neotropica 25(2) At: Flora Neotropica At: Organization for Flora Neotropica
- Bignoniaceae economic botany
- List of genera in family Bignoniaceae At: Dicotyledons At: List Genera within a Family At: Vascular Plant Families and Genera Arquivado em 2012-09-15 no Wayback Machine At: About the Checklist At: World Checklist of Selected Plant Families At: Data Sources Arquivado em 2012-09-10 no Wayback Machine At: ePIC Arquivado em 2012-09-10 no Wayback Machine At: Scientific Databases At: Kew Gardens
- Bignoniaceae At: Advanced Search At: Search Tool At: iplants
- section B In: A checklist of suprageneric names (Alphabetical Listing by Genera) At: Home page of James L. Reveal & C. Rose Broome
- Bignonia At: Plant Names At: IPNI
- CRC World Dictionary of Plant Names: A-C At: Botany & Plant Science At: Life Science At: CRC Press
- Bignoniaceae In: Genera Plantarum vol. 2 part 2 (Bentham & Hooker) At: View Record At: Titles beginning with "G" At: Titles At: Biodiversity Heritage Library
- Spangler & Olmstead (1999)
{kind=link}
{kind=link}
Famílias de plantas com flor (Sistema APG IV) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Grado ANA |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Mesangiospermae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||