Asteroideae
Asteroideae
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Classificação científica | |||||||||||||
| |||||||||||||
| Tribos | |||||||||||||
| |||||||||||||
| Sinónimos[1] | |||||||||||||

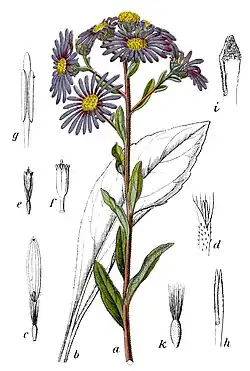


Asteroideae é uma subfamília da família de plantas com flor Asteraceae,[2] que agrupa cerca de 70% das espécies que integram aquela família.[3] A subfamília está repartida por várias tribos, incluindo os grandes agrupamentos taxonómicos Astereae, Calenduleae, Eupatorieae, Gnaphalieae, Heliantheae, Senecioneae e Tageteae. Asteroideae contém plantas com distribuição natural por todo o mundo, muitas das quais arbustivas. Para esta subfamília estão descritos cerca de 1 135 géneros com pelo menos 17 200 espécies. Os maiores géneros em número de espécies são Helichrysum (500–600 espécies) e Artemisia (550 espécies), mas engloba ainda alguns géneros muito conhecidos, entre os quais Santolina, Cynara, Matricaria (ex-Chamomilla), Helianthus e Dahlia. A análise filogenética e o registo fóssil indicam que Asteroideae se diferenciou há cerca 46–36,5 milhões de anos.[4] Asterodae contém muitas plantas economicamente importantes, como a margarida-comum e os ásteres.
Descrição
Os membros da subfamília Asteroideae são, na sua maioria, plantas herbáceas anuais a perenes, existindo também algumas espécies arbustivas relativamente lenhosas. Alguns táxons são suculentos. Os órgãos vegetativos são atravessados por canais de óleo e resina, que se formam por processos intercelulares esquizogénicos, revestidos por epitélio. Não há tubos laticíferos. As folhas são geralmente alternadas ou, por exemplo, em muitas espécies da tribo Heliantheae, opostas,[5] podendo ser simples ou compostas.
Morfologia
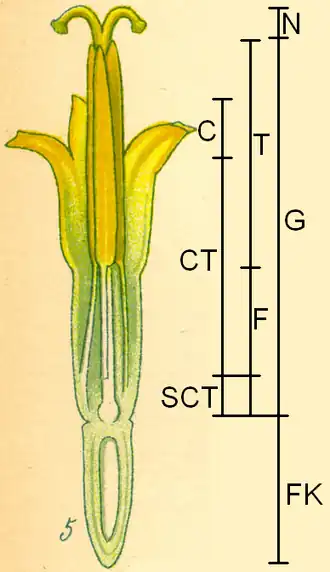
As espécies que integram a subfamília Asteroideae apresentam frequentemente inflorescências em capítulo do tipo radiado, mas algumas podem ter capítulos do tipo discóide ou disciforme. A inflorescência é composta por flósculos raiados com três lóbulos, consideradas flores perfeitas, o que implica que são bissexuais. Muitas contêm superfícies estigmáticas separadas por duas faixas marginais e apêndices terminais estéreis com tricomas longos.[6][7] As inflorescências em forma de capítulo podem ser isoladas ou agrupadas em inflorescências compostas de diferentes estruturas. O capítulo floral é rodeado por bractéolas. O receptáculo pode ter forma de disco ou de cone. Muitas vezes, há folhas modificadas na base do receptáculo semelhantes a páleas (ou praganas) que funcionam como as folhas de cobertura (brácteas) da inflorescência.
Uma característica típica da subfamília Asteroideae é que, muitas vezes, nos capítulos florais, na parte externa, há flores zigomórficas, com três pontas (flores liguladas ou flores radiais), e na parte interna há flores radiariamente simétricas (flores tubulares ou flores discóides).[5] Em alguns táxons faltam as flores liguladas, mas as flores tubulares estão sempre presentes.
As flores têm cinco pétalas (são pentâmeras) com invólucro duplo. As sépalas transformaram-se no chamado pappus. As cinco pétalas estão fundidas numa corola tubular. Nas flores tubulares, os cinco dentes relativamente curtos da corola têm aproximadamente o mesmo comprimento e largura.[5] Nas flores liguladas, as línguas terminam em três dentes na corola,[5] o que constitui uma diferença em relação a outras subfamílias. Nos cinco estames, as anteras estão fundidas formando um tubo. O estilete varre com pelos o pólen do tubo da antera e a abertura dos lóbulos do estigma e a libertação do interior fértil só ocorre em seguida, para evitar a autopolinização. As anteras não possuem apêndices.[5] As espínulas (ou spinuli) estão distribuídas de forma regular ou irregular no grão de pólen.[5] Dois carpelos estão fundidos num ovário ínfero. O estilete termina em duas ramificações. Em cada uma das duas ramificações do estilete, no lado interno, há duas linhas com tecido estigmático.[5]
Os frutos são aquénios, que muitas vezes possuem um pappus para facilitar a dispersão das sementes por anemocoria ou zoocoria (funcionando o papus como um velcro).
O número cromossómico básico é x = 9, 10 ou, na aliança Heliantheae, x = 19.[5]
As espécies desta subfamília apresentam inulina como hidratos de carbono de reserva. Muitas espécies contêm óleos essenciais. Também são importantes os elevados conteúdos em polinas (ou seja, poliacetilenos) e em sesquiterpenlactonas.
Etnobotânica
A subfamília Asteroideae agrupa muitos géneros dentro das suas tribos que têm utilizações económicas relevantes, especialmente como plantas ornamentais e na produção de flores de corte, mas também como oleaginosas, como o girassol, e para alimentação humana.
A espécie Helianthus tuberosus (girassol-batateiro ou tupinambo) é cultivada pelos seus tubérculos comestíveis; as espécies Helianthus annuus (girassol) e Guizotia abyssinica (semente-do-níger ou ramtil) são cultivadas como oleaginosas; a espécie Artemisia dracunculus (estragão) é usada como condimento culinário; e a espécie Parthenium argentatum (guaiúle) é uma fonte de borracha. Alguns dos outros géneros são usados como ornamentais, entre os quais Dendranthema spp. (Chrysanthemum), Callistephus, Cosmos, Tagetes e muitos outros.[8]
Taxonomia e sistemática
Taxonomia
A subfamília Asteroideae foi criada em 1829 por John Lindley em texto incluído na obra de John Claudius Loudon intitulada An Encyclopedia of Plants (p. 1074).[9]
Na sua presente circunscrição taxonómica, a subfamília é composta por 21 tribos (alguns autores consideram apenas 20 tribos, não separando a tribo Doroniceae da tribo Senecioneae; outros, pelo contrário, consideram 22 tribos, separando também uma tribo Plucheeae da tribo Inuleae). A partir de 2004, as tribos foram agrupadas em três supertribos: Senecionodae, Asterodae e Helianthodae.[3][10][11] Um estudo filogenético publicado em 2019 comprova que a supertribo Asterodae com a circunscrição taxonómica que então tinha era polifilético, e em 2024 as tribos Anthemideae e Calenduleae foram recolocadas nas supertribos Anthemidodae e Calendulodae, respetivamente.[12]
As cinco supertribos e 21 tribos estão listadas abaixo. A tribo Senecioneae contém cerca de 120 géneros e mais de 3 200 espécies, maioritariamente encontradas em áreas mais temperadas.[13] A supertribo Helianthodae, que é a maior das três, contém 15 das 21 tribos.[14]
- Anthemodae
- Anthemideae (incluindo os crisântemos)
- Asterodae
- Astereae (incluindo os ásteres e a margarida-comum)
- Gnaphalieae
- Calendulodae
- Calenduleae (incluindo as calêndulas)
- Helianthodae (Ambrosiodae per Feng et al.)
- Athroismeae
- Bahieae
- Chaenactideae
- Coreopsideae (incluindo os cosmos e as dálias)
- Eupatorieae
- Feddeeae
- Helenieae (incluindo as gailárdias)
- Heliantheae (Ambrosieae per Feng et al.; incluindo o girassol e as zínias)
- Inuleae (incluindo Plucheeae)[15]
- Madieae
- Millerieae
- Neurolaeneae
- Perityleae
- Polymnieae
- Tageteae (incluindo os cravos-túnicos e os tagetes)
- Senecionodae
Sistemática




_(6256923212).jpg)


_in_Hyderabad%252C_AP_W_IMG_0010.jpg)
















A subfamília Asteroideae, na sua presente circunscrição taxonómica, contém cerca de 1 135 géneros e cerca de 16 200 espécies, com distribuição mundial.[17] Um grupo de parentesco, a chamada aliança Heliantheae, é formado pelas tribos Feddeeae, Helenieae, Coreopsideae, Neurolaeneae, Tageteae, Chaenactideae, Bahieae, Polymnieae, Heliantheae, Millerieae, Madieae, Perityleae e Eupatorieae.[5]
Na sua divisão mais consensual, a subfamília Asteroideae é dividida em 20 tribos (os géneros estão listados nos artigos de cada uma das tribos):[18]
- Tribo Anthemideae Panero: Contém entre 100 e 110 géneros com cerca de 1700 espécies.[19]
- Tribo Astereae Cass.: Contém mais de 200 géneros com cerca de 3000 espécies.[19]
- Tribo Athroismeae Panero: Desde 2017, contém mais cerca de 9 géneros,[20] com 59 a 80 espécies, distribuídas principalmente na África e na Austrália.
- Tribo Calenduleae Cass.: Anteriormente, continha 8 géneros, mas desde 2009 agrupa 12 géneros com 110 a 120 espécies. Quase sempre não possuem papus.[19]
- Tribo Chaenactideae B.G.Baldwin: Contém apenas 3 géneros, distribuídos desde o oeste da América do Norte até ao México.[21]
- Tribo Eupatorieae Cass.: Contém 167 a 181 géneros, com 2000 a 2400 espécies.[19][22]
- Tribo Gnaphalieae Cass. ex Lecoq & Juill.: Dividido em cinco a seis subtribos, com cerca de 184 a 187 géneros e 1240 a mais de 2000 espécies.[19]
- Tribo Heliantheae Cass.: No presente contém apenas 14 subtribos com apenas cerca de 112 géneros. Algumas novas tribos foram segregadas a partir deste agrupamento.[19][23]
- Tribo Inuleae Cass.: Contém entre 40 e 60 géneros com 500 a 600 espécies.[19]
- Tribo Plucheeae (Cass. ex Dum.) Anderberg: Contém cerca de 27 géneros com cerca de 219 espécies.[19]
- Tribo Senecioneae Cass.: Contém cerca de 150 géneros com cerca de 3200 espécies.[19]
- Tribo Tageteae Cass.: Contém 16 a 23 géneros, com cerca de 216 espécies. O centro da biodiversidade é o planalto seco mexicano.[24]
- Tribo Bahieae B.G.Baldwin: Foi criada em 2002 e contém 19 a 21 géneros, distribuídos principalmente no Neotropis, com maior diversidade de espécies no sudoeste dos EUA e no México. São, na sua maioria, plantas herbáceas anuais a perenes, raramente arbustos, que não contêm seiva leitosa. Não apresentam folhas caulescentes. Quando há flores liguladas, estas são exclusivamente femininas. As línguas das corolas terminam com dois ou três dentes. Nas flores tubulares, as corolas têm quatro ou cinco dentes. Os aquénios são escuros devido à presença de fitomelanina:[21][25][26]
- Achyropappus Kunth:[26] As três espécies existentes desde 2012 ocorrem no México e na Guatemala:[27]
- Apostates N.S.Lander: Contém apenas uma espécie:[26]
- Apostates rapae (F.Brown) N.S.Lander: É uma espécie endémica da Ilha da Páscoa (Rapa Nui, na língua local, daí o epíteto específico rapae).[26]
- Bahia Lag.: Continha cerca de 10 espécies,[19] mas em 2016 contém apenas uma espécie:[26]
- Bahia ambrosioides Lag.: Ocorre em áreas com climas áridos a mediterrânicos, nas costas e nas montanhas costeiras do Chile.[26]
- Bartlettia A.Gray: Contém apenas uma espécie:[19]
- Bartlettia scaposa A.Gray: Está presente nos estados do sudoeste dos Estados Unidos (Novo México e Texas) e daí até ao México.[19]
- Chaetymenia Hook. & Arn.: Contém apenas uma espécie:
- Chaetymenia peduncularis Hook. & Arn.: É comum no México.
- Chamaechaenactis Rydb.: Contém apenas uma espécie:[19]
- Espejoa DC.: Contém apenas uma espécie:
- Espejoa mexicana DC.: É comum no México.
- Florestina Cass.:[26] As cerca de 8 espécies estão distribuídas desde o Texas (apenas uma espécie) até ao México e à América Central.[19]
- Holoschkuhria H.Rob.: Foi criado em 2002 e contém apenas uma espécie:[26]
- Hymenopappus L'Hér.: As cerca de 11 espécies estão distribuídas desde a América do Norte (10 espécies) até ao México.[19]
- Hymenothrix A.Gray: Continha cerca de 5 espécies,[19] mas desde 2016, foram incluídas mais 6 espécies. Entre elas estão as espécies do antigo género Amauriopsis Rydb. que até 2010 continha apenas uma espécie,[19] mas que desde 2010 conta com 6 espécies.[28] As cerca de 11 espécies reconhecidas desde 2016 estão distribuídas nos EUA e no México.[26]
- Hypericophyllum Steetz: As cerca de 7 espécies estão distribuídas pela África tropical.
- Loxothysanus B.L.Rob.: As duas espécies estão distribuídas no México.
- Nothoschkuhria B.G.Baldwin: Foi criado em 2015 e contém apenas uma espécie:[26][29]
- Nothoschkuhria degenerica (Kuntze) B.G.Baldwin: Ocorre desde as terras altas da Bolívia até o noroeste da Argentina.[26]
- Palafoxia Lag.:[26] As cerca de 12 espécies estão distribuídas desde o sudoeste dos EUA (10 espécies) até ao México.[19]
- Peucephyllum A.Gray: Contém apenas uma espécie:
- Peucephyllum schottii A.Gray: Está presente desde os estados do sudoeste dos Estados Unidos (Arizona, Califórnia, Nevada e Utah) até ao noroeste do México (Baja California e Sonora).[19]
- Picradeniopsis Rydb.: As cerca de 8 espécies estão distribuídas desde os EUA até ao México e na América do Sul.[26]
- Platyschkuhria Rydb.:[26] Contém apenas uma espécie:[19]
- Psathyrotopsis Rydb.: As cerca de 3 espécies estão distribuídas desde o sudoeste dos EUA até ao México.[19]
- Schkuhria Roth.:[26] As 2 a 6 espécies estão distribuídas desde o sudoeste dos EUA (2 espécies) até ao México e América Central e do Sul.[19]
- Thymopsis Benth.: Das 2 espécies existentes nas ilhas das Caraíbas, uma ocorre apenas nas Bahamas e a outra apenas em Cuba.
- Tribo Coreopsideae Lindl. (sin.: Bidentideae Godr., Bidentidinae Griseb., Coreopsidaceae Link, Coreopsidinae Dumort., Coreopsidodinae C.Jeffrey):[9] Contém (21 a) cerca de 31 géneros, com cerca de 500 espécies:[30][31]
- Bidens L. (sin.: Megalodonta Greene): As 150 a mais de 250 espécies estão distribuídas por quase todo o mundo.[19] Existem muitas espécies nas regiões temperadas, subtropicais e tropicais do Novo Mundo[19] e na África.[30]
- Chrysanthellum Rich.: As cerca de 11 espécies ocorrem em muitas regiões tropicais do mundo.
- Coreocarpus Benth.: As cerca de 9 espécies são todas nativas do México, uma espécie também ocorre no Arizona.[19]
- Coreopsis L.: O âmbito deste género foi alvo de controvérsia, e as espécies africanas já não pertencem a este género. As cerca de 70 espécies estão amplamente distribuídas pelo Novo Mundo. Os centros de diversidade das espécies, com cerca de 28 espécies,[19] situam-se nas regiões temperadas do leste da América do Norte, no México e nos Andes.[30]
- Cosmos Cav.: As 26 a 28 espécies estão amplamente distribuídas na região neotropical, com maior concentração de biodiversidade no México. Uma espécie é considerada espécie invasora em muitas partes do mundo.[19]
- Cyathomone S.F.Blake: Contém apenas uma espécie:
- Cyathomone sodiroi (Hieron.) S.F.Blake: Ocorre no Equador.
- Dahlia Cav.: As 35 a 40 espécies estão distribuídas principalmente no México, mas também existem espécies na América Central e na Colômbia. Muitas variedades são utilizadas como plantas ornamentais em regiões temperadas a tropicais.[32]
- Dicranocarpus A.Gray: Contém apenas uma espécie:
- Dicranocarpus parviflorus A.Gray: Ocorre desde o sudoeste dos EUA até ao norte do México.[19]
- Diodontium F.Muell.: Contém apenas uma espécie:
- Diodontium filifolium F.Muell.: Comum no norte da Austrália.
- Ericentrodea S.F.Blake: As cerca de seis espécies ocorrem nos Andes, desde a Colômbia até à Bolívia.
- Eryngiophyllum Greenm.: As duas espécies estão distribuídas no México.
- Fitchia Hook. f.: As cerca de 7 espécies estão distribuídas pela Polinésia, 3 espécies são endémicas das Ilhas da Sociedade e uma espécie é endémica de Rapa Iti.
- Glossocardia Cass.: As cerca de 12 espécies estão distribuídas pelo Sudeste Asiático e pelas ilhas do Pacífico.
- Goldmanella Greenm.: Contém apenas uma espécie:
- Goldmanella sarmentosa (Greenm.) Greenm.: Está presente desde o México até à Guatemala, passando por Belize.
- Henricksonia B.L.Turner: Contém apenas uma espécie:
- Henricksonia mexicana B.L.Turner: Ocorre no norte do México.
- Heterosperma Cav.: As 5 a 10 espécies estão distribuídas desde o sudoeste dos EUA (uma espécie) até ao México e América Central e do Sul. São neófitas nas ilhas das Caraíbas.[19]
- Hidalgoa La Llave & Lex.: As cerca de 5 espécies estão distribuídas desde o México até à América Central.
- Isostigma Less.: As 11 a 15 espécies estão distribuídas desde a Bolívia, Brasil, Paraguai e Uruguai até à Argentina.
- Koehneola Urb.: Contém apenas uma espécie:
- Koehneola repens (Griseb. ex Urb.) Urb.: Endemismo em Cuba.
- Moonia Arn.: Contém apenas uma espécie (anteriormente até 6 espécies):
- Moonia heterophylla Arn.: Ocorre no Sri Lanka e no sul da Índia.
- Narvalina Cass.: Contém apenas uma espécie:
- Narvalina domingensis (Cass.) Less.: Só ocorre em Hispaniola.
- Oparanthus Sherff: Desde 2011, inclui 6 espécies endémicas das ilhas da Polinésia, 4 nas Marquesas e 2 em Rapa Iti.
- Petrobium R.Br.: Contém apenas uma espécie:
- Petrobium arboreum (J.R.Forst. & G.Forst.) R.Br. ex Spreng.: Endemismo em Santa Helena.
- Pinillosia Ossa ex DC.: Contém apenas uma espécie:
- Pinillosia berteroi (Spreng.) Urb.: Ocorre em Cuba e Hispaniola.
- Selleophytum Urb. (por vezes agrupado em Coreopsis L.): Foi reativado em 2008 e contém apenas uma espécie:
- Staurochlamys Baker: Contém apenas uma espécie:
- Tetraperone Urb.: Contém apenas uma espécie:
- Tetraperone bellioides (Griseb.) Urb.: Endemismo de Cuba.
- Thelesperma Less.: As mais de 10 espécies estão distribuídas desde o oeste da América do Norte (9 espécies) até ao México e na América do Sul.[19]
- Trioncinia (F.Muell.) Veldkamp: Contém apenas uma espécie:
- Trioncinia retroflexa (F.Muell.) Veldkamp: Ocorre apenas no estado australiano de Queensland.
- Tribo Feddeeae Pruski, P.Herrera, Anderb. & Franc.-Ort.: Foi criada em 2008 e contém apenas um género:[33]
- Feddea Urb.: Contém apenas uma espécie:
- Feddea cubensis Urb.: Nativa apenas da região oriental de Cuba e está em perigo de extinção.
- Feddea Urb.: Contém apenas uma espécie:
- Tribo Helenieae Benth. & Hook.: Contém cerca de 12 géneros no Novo Mundo, com a maior diversidade de espécies concentrada no sul dos EUA e no norte do México:
- Amblyolepis DC.: Contém apenas uma espécie:
- Amblyolepis setigera DC.: A sua distribuição estende-se desde o estado norte-americano do Texas até aos estados mexicanos de Coahuila, Nuevo León e Tamaulipas.[19]
- Baileya Harv. & A.Gray ex A.Gray: As três únicas espécies distribuem-se desde o sudoeste dos EUA até ao norte do México.[19]
- Balduina Nutt.: As três únicas espécies distribuem-se no sudeste dos EUA.[19]
- Gaillardia Foug.: As 15 a 17 ou mais espécies distribuem-se na América do Norte (11 espécies), no México e na América do Sul (principalmente na Argentina).[19]
- Helenium L.: As cerca de 32 espécies encontram-se disseminadas no Novo Mundo.[19]
- Hymenoxys Cass.: As cerca de 25 espécies distribuem-se pela América do Norte (17 espécies), México e América Central e do Sul.[19]
- Marshallia Schreb.: As cerca de 7 espécies estão distribuídas no sul, centro e sudeste dos EUA.[19]
- Pelucha S.Wats.: Contém apenas uma espécie:
- Pelucha trifida S.Wats.: Só se desenvolve no deserto mexicano de Sonora e na Baixa Califórnia.
- Plateilema (A.Gray) Cockerell: Contém apenas uma espécie:
- Plateilema palmeri (A.Gray) Cockerell: Está presente desde o estado norte-americano do Texas até aos estados mexicanos de Coahuila e Nuevo León.[19]
- Psathyrotes (Nutt.) A.Gray: As 3 únicas espécies distribuem-se desde o sudoeste dos EUA até ao noroeste do México.[19]
- Psilostrophe DC.: As cerca de 7 espécies distribuem-se desde os EUA (seis espécies) até ao México.[19]
- Tetraneuris Greene: As cerca de 9 espécies distribuem-se desde a América do Norte (9 espécies) até ao México.[19]
- Trichoptilium A.Gray: Contém apenas uma espécie:
- Trichoptilium incisum (A.Gray) A.Gray: Está disseminada desde os estados norte-americanos do Arizona, Califórnia e Nevada até à Baixa Califórnia, no México.[19]
- Amblyolepis DC.: Contém apenas uma espécie:
- Tribo Madieae Jeps.: Contém cerca de 35 géneros no Novo Mundo:[19]
- Achyrachaena Schauer: Contém apenas uma espécie:
- Achyrachaena mollis Schauer: Ocorre desde Oregon via Califórnia até à Baja Califórnia no México.[19]
- Adenothamnus D.D.Keck: Contém apenas uma espécie:
- Adenothamnus validus (T.S.Brandegee) D.D.Keck: Ocorre no México.
- Argyroxiphium DC.: Das 5 espécies existentes até à data, uma, Argyroxiphium virescens Predefinição:Pessoa está extinta. Todas elas ocorrem apenas no Hawaii. Pertence a este género:
- Argyroxiphium sandwicense DC.
- Amblyopappus Hook. & Arn.: Contém apenas uma espécie:
- Amblyopappus pusillus Hook. & Arn.: Ocorre nas costas da Califórnia até à Baja Califórnia, no México e na costa ocidental da América do Sul.[19]
- Anisocarpus Nutt.: Das duas únicas espécies, uma está disseminada na parte ocidental da América do Norte e a outra ocorre apenas na Califórnia.[19]
- Arnica L.: As cerca de 29 espécies encontram-se na América do Norte (26 espécies), no México, na Europa, na parte asiática da Rússia e no Japão.[19]
- Baeriopsis J.T.Howell: Contém apenas uma espécie:
- Baeriopsis guadalupensis J.T.Howell: Ocorre apenas na Baixa Califórnia.
- Blepharipappus Hook.: Contém apenas uma espécie:
- Blepharipappus scaber Hook.: Ocorre em altitudes de 300 a 2200 metros acima do nível médio do mar no noroeste dos EUA.[19]
- Blepharizonia (A.Gray) Greene: Apenas duas espécies que ocorrem na Califórnia.[19]
- Calycadenia DC.: As cerca de 10 espécies estão disseminadas no oeste dos EUA.[19]
- Carlquistia B.G.Baldwin: Contém apenas uma espécie:
- Carlquistia muirii (A.Gray) B.G.Baldwin: Desenvolve-se em altitudes de 1100 a 2500 metros, ocorrendo apenas na Califórnia.[19]
- Centromadia Greene: As cerca de 4 espécies distribuem-se desde o oeste dos EUA (3 espécies) até ao noroeste do México.[19]
- Constancea B.G.Baldwin: Contém apenas uma espécie:
- Constancea nevinii (A.Gray) B.G.Baldwin: Endemismo das ilhas do Canal da Califórnia.
- Deinandra Greene: As cerca de 21 espécies distribuem-se desde o oeste dos EUA (16 espécies) até ao noroeste do México.[19]
- Dubautia Gaudich.: As cerca de 39 espécies ocorrem apenas no Hawaii.
- Eatonella A.Gray: Contém apenas uma espécie:
- Eatonella nivea (D.C.Eaton) A.Gray: Ocorre em altitudes de 800 a 3100 metros no oeste dos EUA.[19]
- Eriophyllum Lag.: As cerca de 14 espécies estão distribuídas por todo o oeste dos EUA, com um foco de biodiversidade na Califórnia.[19]
- Harmonia B.G.Baldwin: As cerca de 5 espécies ocorrem apenas na Califórnia.[19]
- Hemizonella (A.Gray) A.Gray: Contém apenas uma espécie:
- Hemizonella minima (A.Gray) A.Gray: Ocorre em altitudes de 300 a 2900 metros no oeste da América do Norte.[19]
- Hemizonia DC.: Contém apenas uma espécie:
- Hemizonia congesta DC.: Está disseminada no oeste dos EUA, com 6 subespécies.[19]
- Holocarpha Greene: As cerca de 4 espécies ocorrem apenas na Califórnia.[19]
- Holozonia Greene: Contém apenas uma espécie:
- Holozonia filipes (Hook. & Arn.) Greene: Ocorre em altitudes de 30 a 600 metros, apenas na Califórnia.[19]
- Hulsea Torr. & A.Gray: As cerca de 7 espécies distribuem-se desde o oeste dos EUA até ao noroeste do México.[19]
- Jensia B.G.Baldwin: Apenas duas espécies, com ocorrência na Califórnia.[19]
- Kyhosia B.G.Baldwin: Contém apenas uma espécie:
- Kyhosia bolanderi (A.Gray) B.G.Baldwin: Ocorre em altitudes de 1000 a 2600 metros, nos estados da Califórnia, Nevada e Oregon, no oeste dos EUA.
- Lagophylla Nutt.: As cerca de 4 espécies estão disseminadas na parte ocidental da América do Norte.[19]
- Lasthenia Cass.: Das cerca de 18 espécies, 17 distribuem-se na América do Norte, principalmente na Califórnia, e uma espécie ocorre no Chile.[19]
- Layia Hook. & Arn. ex DC.: As cerca de 14 espécies distribuem-se desde o oeste da América do Norte (todas as 14 espécies) até ao noroeste do México.[19]
- Madia Molina: As cerca de 10 espécies estão espalhadas pelo Novo Mundo. A espécie Madia sativa era anteriormente cultivada como planta oleaginosa. É nativa do Canadá, dos EUA e da Argentina.[18] Madia sativa também ocorre no Havai; não é possível comprovar com dados se se trata de uma ocorrência natural ou se é um neófito.[19]
- Monolopia DC.: As cerca de 5 espécies ocorrem apenas na Califórnia.[19]
- Osmadenia Nutt.: Contém apenas uma espécie:
- Osmadenia tenella Nutt.: Encontra-se apenas na Califórnia e na Baixa Califórnia, no México.[19]
- Pseudobahia (A.Gray) Rydb.: As 3 únicas espécies só se encontram na Califórnia.[19]
- Raillardella (A.Gray) Benth. in Benth. & Hook. f.: Apenas 3 espécies, com distribuição restrita ao oeste dos EUA.[19]
- Syntrichopappus A.Gray: As duas únicas espécies encontram-se no sudoeste dos EUA.[19]
- Wilkesia A.Gray: As duas únicas espécies são endémicas da ilha havaiana de Kauaʻi.
- Venegasia DC.: Contém apenas uma espécie:
- Venegasia carpesioides DC.: Ocorre apenas na Califórnia e na Baixa Califórnia, no México.[19]
- Achyrachaena Schauer: Contém apenas uma espécie:
- Tribo Millerieae Lindl. (sin.: subtribo Milleriinae): Contém cerca de 34 géneros:[34][35]
- Acanthospermum Schrank: As cerca de 6 espécies distribuem-se principalmente pelas regiões temperadas quentes e tropicais do Novo Mundo. Algumas espécies são neófitas em muitas zonas do mundo.[19]
- Alepidocline S.F.Blake: As cerca de 4 espécies estão disseminadas na região neotropical.
- Alloispermum Willd.: As cerca de 16 espécies estão distribuídas na região neotropical.
- Aphanactis Wedd.: As cerca de 11 espécies estão distribuídas na região neotropical.
- Axiniphyllum Benth.: As cerca de 5 espécies encontram-se no México.
- Bebbia Greene: As duas únicas espécies distribuem-se desde o sudoeste dos EUA (apenas uma espécie) até ao norte do México.[19]
- Carramboa Cuatrec. (por vezes em Espeletia): As 4 a 7 espécies ocorrem nos Andes.
- Coespeletia Cuatrec. (por vezes em Espeletia): As 7 a 8 espécies (com a circunscrição de 2013) ocorrem nos Andes da Venezuela e da Colômbia.
- Cymophora B.L.Rob.: As 4 ou 5 espécies ocorrem no México e na Venezuela.
- Desmanthodium Benth.: As cerca de 7 espécies estão distribuídas na região neotropical.
- Dyscritothamnus B.L.Rob.: As cerca de duas espécies encontram-se no México.
- Espeletia Mutis ex Humb. & Bonpl.: Este género foi dividido em sete géneros mais pequenos. As 45 a 71 espécies ocorrem nos páramos do norte da América do Sul, na Venezuela, Colômbia e Equador.
- Espeletiopsis Cuatrec. (por vezes em Espeletia): Das cerca de 23 espécies, 5 espécies crescem nos Andes centrais da Venezuela e 18 espécies na Cordilheira Oriental da Colômbia.
- Faxonia Brandegee: Contém apenas uma espécie:
- Faxonia pusilla Brandegee: Ocorre no México.
- Galinsoga Ruiz & Pav.: As 15 a mais de 33 espécies são apenas nativas do Novo Mundo. Algumas espécies são neófitas nos outros continentes.[19]
- Guardiola Cerv. ex Bonpl.: As cerca de 10 espécies são todas nativas do México, com uma espécie a ocorrer também no Arizona.[19]
- Guizotia Cass.: As cerca de 6 espécies estão distribuídas na África tropical. Guizotia abyssinica (L. f.) Cass. é uma neófita em muitas áreas do mundo.[19]
- Ichthyothere Mart.: As cerca de 20 espécies distribuem-se do Panamá à América do Sul.
- Jaegeria Kunth: As cerca de 10 espécies estão disseminadas na região neotropical.
- Lecocarpus Decne.: As 2 a 3 espécies são endemismos das Galápagos.
- Libanothamnus Cuatrec. (por vezes em Espeletia): Contém cerca de 11 espécies, com ocorrência nos Andes.
- Melampodium L.: As cerca de 36 espécies têm distribuição natural no Novo Mundo, desde os EUA (7 espécies), passando pelo México, América Central e ilhas das Caraíbas, até à América do Sul. Pelo menos uma espécie é neófita em muitas zonas do mundo.[19]
- Micractis DC.: As cerca de 4 espécies encontram-se na África Oriental e em Madagáscar.
- Milleria L.: As duas únicas espécies distribuem-se desde o México, passando pela América Central, até ao norte da América do Sul.
- Oteiza La Llave: Contém 5 espécies na Guatemala.
- Paramiflos Cuatrec. (às vezes em Espeletia): Contém apenas uma espécie:
- Paramiflos glandulosus (Cuatrec.) Cuatrec.: Ocorre nos Andes.
- Ruilopezia Cuatrec. (às vezes em Espeletia): Contém cerca de 24 espécies nos Andes.
- Rumfordia DC.: As 7 ou 8 espécies estão distribuídas do México à Guatemala.
- Sabazia Cass.: As cerca de 16 espécies estão distribuídas pela América Central.
- Schistocarpha Less.: As cerca de 13 espécies estão distribuídas na região neotropical.
- Selloa Kunth: Existem cerca de 6 espécies, distribuídas desde o México até à América Central e do Sul.
- Sigesbeckia L.: As cerca de 8 espécies estão distribuídas pelo México.
- Smallanthus Mack.: As cerca de 20 espécies estão distribuídas na região neotropical.[19]
- Stachycephalum Sch.Bip. ex Benth.: As cerca de 3 espécies estão distribuídas pelo México, Equador e Argentina.
- Tamania Cuatrec. (às vezes em Espeletia): Contém apenas uma espécie:
- Tamania chardonii (A.C.Sm.) Cuatrec.: Ocorre nos Andes.
- Tamananthus V.M.Badillo: Contém apenas uma espécie:
- Tamananthus crinitus V.M.Badillo: Ocorre na Venezuela.
- Tetragonotheca L.: As cerca de quatro espécies estão distribuídas desde o sul dos EUA (todas as 4 espécies) até ao norte do México.[19]
- Tridax L.: As cerca de 26 espécies estão distribuídas na região neotropical.[19]
- Trigonospermum Less.: As cerca de 4 espécies estão distribuídas desde o México até à América Central.
- Unxia L. f.: As 3 espécies estão distribuídas desde o Panamá até ao norte da América do Sul.
- Tribo Neurolaeneae Rydb. (sin.: subtribo Neurolaeninae) (Rydb.) Stuessy, Turner & Powell: Contém cerca de 5 géneros:
- Calea L.: As cerca de 95 espécies estão distribuídas na região neotropical. Entre elas:
- Calea ternifolia Kunth: Ocorre na América Central.
- Enydra Lour.: Contém de 3 a 10 espécies, principalmente na América do Sul.
- Greenmaniella W.M.Sharp.: Contém apenas uma espécie:
- Greenmaniella resinosa (W.Watson) W.Sharp: Ocorre no México.
- Heptanthus Griseb.: As cerca de 7 espécies só existem em Cuba.
- Neurolaena R.Br.: As cerca de 9 espécies estão distribuídas pelo México, América Central e Andes.
- Calea L.: As cerca de 95 espécies estão distribuídas na região neotropical. Entre elas:
- Tribo Perityleae B.G.Baldwin: Contém cerca de 7 géneros que têm o seu centro de biodiversidade no México:
- Amauria Benth.: As duas a três espécies ocorrem no México.
- Eutetras A.Gray: As duas espécies estão distribuídas no México.
- Galeana La Llave & Lex.: As duas espécies estão distribuídas desde o México até à Costa Rica.
- Lycapsus Phil.: Contém apenas uma espécie:
- Lycapsus tenuifolius Phil.: Esta espécie endémica ocorre apenas no arquipélago das Ilhas Desventuradas, nas ilhas San Felix e San Ambrosio.
- Pericome A.Gray: As duas espécies estão distribuídas desde o sudoeste dos EUA (uma espécie) até ao norte do México.[19]
- Perityle Benth.: As cerca de 66 espécies estão distribuídas desde os EUA (35 espécies) até ao México e à América do Sul.[19]
- Villanova Lag.: As cerca de 10 espécies estão distribuídas desde o México até à América do Sul, passando pela América Central.
- Tribo Polymnieae (H.Rob.) Panero (sin.: subtribo Polymniinae): Foi criado em 2002 e contém apenas um género:
(Algumas espécies, como os cardos, são classificadas na subfamília Carduoideae, de acordo com novos conhecimentos filogenéticos.)
Referências
- ↑ UniProt. «Asteroideae» (HTML). Consultado em 13 de junho de 2008
- ↑ «Asteroideae Lindl». Tropicos. Consultado em 21 de julho de 2019
- ↑ a b Asteraceae, Tree of Life Web Project
- ↑ Huang, C. -H; Zhang, C; Liu, M; Hu, Y; Gao, T; Qi, J; Ma, H (2016). «Multiple polyploidization events across Asteraceae with two nested events in the early history revealed by nuclear phylogenomics». Mol. Biol. Evol. 33 (11): 2820–2835. PMC 5062320
 . PMID 27604225. doi:10.1093/molbev/msw157
. PMID 27604225. doi:10.1093/molbev/msw157
- ↑ a b c d e f g h i Vicki A. Funk, Alfonso Susanna, Tod F. Stuessy, Harold Robinson: Chapter 42: Classification of Compositae. S. 171–192: In: Vicki A. Funk, A. Susanna, T. F. Stuessy, R. J. Bayer (Hrsg.): Systematics, Evolution and Biogeography of the Compositae. International Organization of Plant Taxonomy = IAPT, Wien, 2009. online
- ↑ Lindley, J., The Vegetable Kingdom
- ↑ Vicki A. Funk, Alfonso Susanna, Tod F. Stuessy, Harold Robinson: Chapter 11: Classification of Compositae. pp. 171–189. online In: Vicki A. Funk, Alfonso Susanna, Tod F. Stuessy, R. Bayer (Hrsg.): Systematics, Evolution, and Biogeography of Compositae, IAPT, Vienna 2009.
- ↑ Murrell, Z.E (2010). Vascular Plant Taxonomy. [S.l.]: Kendall Hunt Publishing Compant
- ↑ a b «Asteroideae». Tropicos. Missouri Botanical Garden. 100440357
- ↑ Robinson, Harold (2004). «New supertribes, Helianthodae and Senecionodae, for the subfamily Asteroideae (Asteraceae)». Phytologia. 86 (86): 116–120. ISSN 0031-9430. doi:10.5962/bhl.part.28428. Consultado em 20 de outubro de 2012
- ↑ Harold Robinson; Edward Schilling; José L. Panero. «Eupatorieae» (PDF). p. 14. Consultado em 20 de outubro de 2012
- ↑ Feng, Z.-H., Huang, Z.-J., Sun, H.-N., & Liu, S. (2024). Meeting Necessity instead of Serendipity: Miscellaneous Nomenclatural Notes on Asteraceae. Phytotaxa, 644(2), 124–134. https://doi.org/10.11646/phytotaxa.644.2.4
- ↑ (em inglês) Asteraceae tribe Senecioneae em Flora of North America
- ↑ Panero, J.L; Crozier, B.S. «Asteraceae: Sunflowers, daisies». Tree of Life. Consultado em 6 de abril de 2019
- ↑ Anderberg, A (2005), «Evolutionary relationships in the Asteraceae tribe Inuleae (incl. Plucheeae) evidenced by DNA sequences of F; with notes on the systematic positions of some aberrant genera», Organisms Diversity & Evolution, 5 (2), pp. 135–146, Bibcode:2005ODivE...5..135A, doi:10.1016/j.ode.2004.10.015
- ↑ Fernandez, I (2001), «A Phylogenetic Analysis of Doronicum (Asteraceae, Senecioneae) Based on Morphological, Nuclear Ribosomal (ITS), and Chloroplast (trnL-F) Evidence», Molecular Phylogenetics and Evolution, 20 (1), pp. 41–64, Bibcode:2001MolPE..20...41F, CiteSeerX 10.1.1.331.4339, PMID 11421647, doi:10.1006/mpev.2001.0954
- ↑ Zhu Shi, Yilin Chen, Yousheng Chen, Yourun Lin (Ling Yuou-ruen), Shangwu Liu, Xuejun Ge, Tiangang Gao, Shixin Zhu, Ying Liu, Christopher J. Humphries, Qiner Yang, Eckhard von Raab-Straube, Michael G. Gilbert, Bertil Nordenstam, Norbert Kilian, Luc Brouillet, Irina D. Illarionova, D. J. Nicholas Hind, Charles Jeffrey, Randall J. Bayer, Jan Kirschner, Werner Greuter, Arne A. Anderberg, John C. Semple, Jan Štěpánek, Susana Edith Freire, Ludwig Martins, Hiroshige Koyama, Takayuki Kawahara, Leszek Vincent, Alexander P. Sukhorukov, Evgeny V. Mavrodiev, Günter Gottschlich: Asteraceae. In: Wu Zheng-yi, Peter H. Raven, Deyuan Hong (Hrsg.): Flora of China. Volume 20–21, Science Press und Missouri Botanical Garden Press, Beijing und St. Louis 2011, ISBN 978-1-935641-07-0. ab S. 1 - textgleich online wie gedrucktes Werk.
- ↑ a b «Asteroideae». Agricultural Research Service (ARS), United States Department of Agriculture (USDA). Germplasm Resources Information Network (GRIN)
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf bg bh bi bj bk bl bm bn bo bp bq br bs bt bu bv bw bx by bz ca cb cc cd ce cf cg ch Theodore M. Barkley, Luc Brouillet, John L. Strother: Asteraceae. In: Flora of North America Editorial Committee (Hrsg.): Flora of North America North of Mexico, Volume 19, 20 und 21 - Magnoliophyta: Asteraceae, Oxford University Press, New York und Oxford, 30. Juni 2006, ISBN 0-19-530565-5. - textgleich online wie gedrucktes Werk.
- ↑ Annika Bengtson, Markus Englund, John F. Pruski, Arne Alfred Anderberg: Phylogeny of the Athroismeae (Asteraceae), with a new circumscription of the tribe. In: Taxon, Volume 66, Issue 2, Mai 2017, S. 408–420. doi:10.12705/662.8
- ↑ a b B. L. Turner: The Comps of Mexico. A Systematic Account of the Family Asteraceae. Chapter 12: Tribes Bahieae and Chaenactideae, In: Phytologia Memoirs, Volume 17, 2013.
- ↑ Harold E. Robinson, Edward E. Schilling, Jose L. Panero: Chapter 43: Eupatorieae., 731-744: In: Vicki A. Funk, A. Susanna, T. F. Stuessy, R. J. Bayer (Hrsg.): Systematics, Evolution and Biogeography of the Compositae. International Organization of Plant Taxonomy = IAPT, Wien, 2009. Volltext-PDF.
- ↑ Edward E. Schilling, Jose L. Panero: A revised classification of subtribe Helianthinae (Asteraceae: Heliantheae) II. Derived lineages. In: Botanical Journal of the Linnean Society, Volume 167, Issue 3, 2011, S. 311–331. doi:10.1111/j.1095-8339.2011.01172.x
- ↑ D. J. Loockerman, B. L. Turner, R. K Jansen: Phylogenetic relationships within the Tageteae (Asteraceae) based on nuclear ribosomal ITS and chloroplast ndhF gene sequences. In: Systematic Botany, Volume 28, Issue 1, 2003, S. 191–207. Volltext auf BioOne.
- ↑ Tribe Bahieae (Memento vom 8. maio 2014 im Internet Archive) bei Virtual Key to the Compositae Project = VKC der The International Compositae Alliance = TICA. (Memento vom 10. dezembro 2013 im Internet Archive)
- ↑ a b c d e f g h i j k l m n o p B. G. Baldwin, K. R. Wood: Origin of the Rapa endemic genus Apostates: Revisiting major disjunctions and evolutionary conservatism in the Bahia alliance (Compositae: Bahieae). In: Taxon, Volume 65, Issue 5, 2016, S. 1064–1080. doi:10.12705/655.8 Volltext-PDF.
- ↑ Billie L. Turner: A new species of Achyropappus (Asteraceae: Bahieae) from Querétaro, Mexico. In: Phytoneuron 2012-83, 12. September 2012, S. 1–5. PDF, eingescannt bei biodiversitylibrary.org.
- ↑ Billie L. Turner: Rescension of the Mexican Amauriopsis (Asteraceae: Bahieae). In: Phytoneuron, 2010-10, 2010, pp. 1–7, Volltext-PDF.
- ↑ B. G. Baldwin: Nothoschkuhria, a new South American genus for Schkuhria degenerica (Compositae, Bahieae). In: Phytoneuron, 2015-56, 2015, pp. 1–2.
- ↑ a b c Rebecca T. Kimball, Daniel J. Crawford: Phylogeny of Coreopsideae (Asteraceae) using ITS sequences suggests lability in reproductive characters. In. Molecular Phylogenetics and Evolution, Volume 33, 2004, S. 127–139.
- ↑ a b D. J. Crawford, Mesfin Tadesse, Mark E. Mort, Rebecca T. Kimball, Christopher P. Randle: Chapter 42: Coreopsideae., S. 747–777: In: V. A. Funk, A. Susanna, T. F. Stuessy, R. J. Bayer (Hrsg.): Systematics, Evolution and Biogeography of the Compositae. International Organization of Plant Taxonomy = IAPT, Wien, 2009. Volltext-PDF.
- ↑ Dayle E. Saar, Neil O. Polans, Paul D. Sørensen: A Phylogenetic Analysis of the Genus Dahlia (Asteraceae) Based on Internal and External Transcribed Spacer Regions of Nuclear Ribosomal DNA. In: Systematic Botany, Volume 28, Issue 3, 2003, S. 627–639. doi:10.1043/01-78.1
- ↑ Kathleen A. Cariaga, John F. Pruski, Ramona Oviedo, Arne A. Anderberg, Carl E. Lewis, Javier Francisco-Ortega: Phylogeny and Systematic Position of Feddea (Asteraceae: Feddeeae): a Taxonomically Enigmatic and Critically Endangered Genus Endemic to Cuba, In: Systematic Botany, Volume 33, Issue 1, 2008, S. 193–202. doi:10.1600/036364408783887348
- ↑ Tribe Millerieae (Memento vom 8. maio 2014 im Internet Archive) bei Virtual Key to the Compositae Project = VKC der The International Compositae Alliance = TICA. (Memento vom 10. dezembro 2013 im Internet Archive)
- ↑ Mauricio Diazgranados: A nomenclator for the frailejones (Espeletiinae Cuatrec., Asteraceae). In: PhytoKeys, Volume 16, 20. August 2012, S. 1–52. doi:10.3897/phytokeys.16.3186
Bibliografia
- Peter B. Pelser, Linda E. Watson: Introduction to Asteroideae. S. 495–502: In: Vicki A. Funk, A. Susanna, T. F. Stuessy, R. J. Bayer (Hrsg.): Systematics, Evolution and Biogeography of the Compositae. International Organization of Plant Taxonomy = IAPT, Wien, 2009.
- Vicki A.Funk, Arne A.Anderberg, Bruce G. Baldwin, Randall J. Bayer, J. Mauricio Bonifacino, Use Breitwieser, Luc Brouillet, Rodrigo Carhajal, Raymund Chan, Antonio X. P. Coutinho, Daniel J. Crawford, Jorge V. Crisci, Michael O.Dillon, Susana E. Freire, Merce Galhany-Casals, Nuria Garcia-Jacas, Birgit Gemeinholzer, Michael Gruenstaeudl, Hans V.Hansen, Sven Himmelreich, Joachim W. Kadereit, Mari Kallersjo, Vesna Karaman-Castro, Per Ola Karis, Liliana Katinas, Sterling C. Keeley, Norhert Kilian, Rebecca T. Kimball, Timothy K. Lowrey, Johannes Lundberg, Robert J. McKenzie, Mesjin Tadesse, Mark E. Mort, Bertil Nordenstam, Christoph Oberprieler, Santiago Ortiz, Pieter B. Pelser, Christopher P. Randle, Harold Robinson, Nddia Roque, Gisela Sancho, John C. Semple, Miguel Serrano, Tod F. Stuessy, Alfonso Susanna, Matthew Unwin, Lowell Urbatsch, Estrella Urtubey, Joan Valles, Robert Vogt, Steve Wagstaff, Josephine Wardand, Linda E. Watson: Compositae metatrees: the next generation. 2009, S. 747–778. PDF. In: Vicki A. Funk, A. Susanna, T. F. Stuessy, R. J. Bayer (Hrsg.): Systematics, Evolution and Biogeography of the Compositae. International Organization of Plant Taxonomy = IAPT, Wien, 2009.
- J. R. Mandel, R. B. Dikow, Vicki A. Funk, R. R. Masalia, S. E. Staton, A. Kozik, R. W. Michelmore, L. H. Rieseberg, J. M. Burke: A target enrichment method for gathering phylogenetic information from hundreds of loci: An example from the Compositae. In: Applications in Plant Sciences, Volume 6, 2, 2, Februar 2014. doi:10.3732/apps.1300085
- Chien-Hsun Huang, Caifei Zhang, Mian Liu, Yi Hu, Tiangang Gao, Ji Qi, Hong Ma: Multiple polyploidization events across Asteraceae with two nested events in the early history revealed by nuclear phylogenomics. In: Molecular Biology and Evolution, Volume 33, Issue 11, 2016, S. 2820–2835. doi:10.1093/molbev/msw157
- John F. Pruski, Harold E. Robinson: Asteraceae. S. 1–608. In: G. Davidse, M. Sousa Sánchez, S. Knapp, F. Chiang Cabrera (Hrsg.): Flora Mesoamericana, Volume 5 (2): i–xix, Missouri Botanical Garden, St. Louis, 2018.