P3b
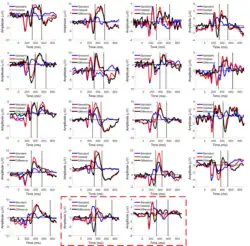
O P3b é um subcomponente do P300, um componente do potencial relacionado a eventos (ERP) que pode ser observado em gravações de atividade elétrica cerebral no couro cabeludo humano. O P3b é uma amplitude positiva (geralmente relativa a uma referência atrás da orelha ou à média de duas dessas referências) com pico em torno de 300 ms, embora o pico varie em latência (atraso entre o estímulo e a resposta) de 250 a 500 ms ou mais tarde, dependendo da tarefa[2] e da resposta individual do sujeito.[1] As amplitudes são tipicamente mais altas no couro cabeludo do que nas áreas parietais do cérebro.[1] Amplitudes are typically highest on the scalp over parietal brain areas.[2]
O P3b tem sido uma ferramenta proeminente usada para estudar processos cognitivos por várias décadas. Mais especificamente, este componente ERP desempenhou um papel fundamental na pesquisa de psicologia cognitiva sobre processamento de informações. De modo geral, eventos improváveis irão provocar um P3b, e quanto menos provável o evento, maior o P3b.[3] No entanto, para provocar um P3b, o evento improvável deve estar relacionado à tarefa em questão de alguma forma (por exemplo, o evento improvável pode ser uma letra alvo pouco frequente em um fluxo de letras, à qual um sujeito pode responder com um pressionamento de botão). O P3b também pode ser usado para medir o quão exigente uma tarefa é na carga de trabalho cognitiva.[3]
História
As primeiras observações do P3b foram relatadas em meados da década de 1960. Em 1964, os pesquisadores Chapman e Bragdon[4] descobriram que as respostas do ERP aos estímulos visuais diferiam dependendo se os estímulos tinham significado ou não. Eles mostraram aos sujeitos dois tipos de estímulos visuais: números e flashes de luz. Os sujeitos viam esses estímulos um de cada vez em uma sequência. Para cada dois números, os sujeitos eram solicitados a tomar decisões simples, como dizer qual dos dois números era numericamente menor ou maior, qual vinha primeiro ou segundo na sequência ou se eram iguais. Ao examinar os potenciais evocados para esses estímulos (ou seja, ERPs), Chapman e Bragdon descobriram que tanto os números quanto os flashes provocavam as respostas sensoriais esperadas (por exemplo, componentes visuais N1) e que a amplitude dessas respostas variava de maneira esperada com a intensidade dos estímulos. Eles também descobriram que as respostas do ERP aos números, mas não aos flashes de luz, continham uma grande positividade que atingia o pico em torno de 300 ms após o estímulo aparecer.[4] Eles também notaram que a amplitude dessa positividade não foi afetada pela intensidade do estímulo. Chapman e Bragdon especularam que essa resposta diferencial aos números, que veio a ser conhecida como resposta P300, resultou do fato de que os números eram significativos para os participantes, com base na tarefa que eles foram solicitados a realizar.
Em 1965, Sutton e colegas publicaram os resultados de dois experimentos que exploraram ainda mais essa positividade tardia. Eles apresentaram aos sujeitos uma pista que indicava se o estímulo seguinte seria um clique ou um flash, ou uma pista que exigia que os sujeitos adivinhassem se o estímulo seguinte seria um clique ou um flash. Eles descobriram que, quando os sujeitos eram solicitados a adivinhar qual seria o estímulo seguinte, a amplitude do "complexo positivo tardio"[4] era maior do que quando sabiam qual seria o estímulo. Em um segundo experimento,[5] eles apresentaram dois tipos de pistas. Para uma pista, havia uma chance de 2 em 3 de que o estímulo seguinte fosse um clique e uma chance de 1 em 3 de que o estímulo seguinte fosse um flash. O segundo tipo de pista tinha probabilidades inversas às do primeiro. Eles descobriram que a amplitude do complexo positivo era maior em resposta aos estímulos menos prováveis, ou seja, aquele que tinha apenas uma chance em 3 de aparecer. Outra descoberta importante desses estudos é que esse complexo positivo tardio foi observado tanto para os cliques quanto para os flashes, indicando que o tipo físico do estímulo (auditivo ou visual) não importava.[5]
Em estudos posteriores publicados em 1967, Sutton e colegas fizeram com que os sujeitos adivinhassem se ouviriam um clique ou dois cliques.[6] Eles observaram uma positividade em torno de 300 ms após o segundo clique ocorrer ou teria ocorrido no caso do clique único. Eles também fizeram com que os sujeitos adivinhassem qual seria a duração do intervalo entre os cliques, e a positividade tardia ocorreu 300 ms após o segundo clique.[6] Isso mostra duas descobertas importantes: primeiro, que essa positividade tardia ocorreu quando a incerteza sobre o tipo de clique foi resolvida e, segundo, que mesmo a ausência de um estímulo, quando relevante para a tarefa, provocaria o complexo positivo tardio. Esses estudos iniciais encorajaram o uso de métodos ERP para estudar a cognição e forneceram uma base para o extenso trabalho sobre o P3b nas décadas seguintes.[7]
Desde a descoberta inicial deste componente do ERP, a pesquisa mostrou que o P300 não é um fenômeno unitário. Em vez disso, podemos distinguir entre dois subcomponentes do P300: o P3 da novidade, ou P3a, e o P3 clássico, ou P3b.[8]
Características dos componentes
Supondo que uma referência cefálica seja usada (ou seja, um eletrodo de referência colocado em algum lugar na cabeça, como a ponta do nariz ou o queixo), o P3b é um ERP positivo cuja latência na amplitude de pico é geralmente de cerca de 300 ms para estímulos sensoriais simples.[9] A amplitude foi definida como a diferença entre a voltagem média da linha de base pré-estímulo e a voltagem do maior pico (neste caso, positivo) da forma de onda do ERP em uma janela de tempo específica.[2] A amplitude do P3b é geralmente relativamente grande (10–20 microvolts), mas varia sistematicamente em função de uma série de fatores importantes. A latência foi definida como o tempo desde o início do estímulo (ou qualquer que seja o ponto de medição desejado) até o ponto de amplitude máxima.[2] A latência do P3b é geralmente em torno de 300 ms, embora possa variar dentro de uma janela de tempo de cerca de 250–500 ms (ou mais tarde), dependendo de fatores como as condições da tarefa e a idade dos sujeitos.[10]
A distribuição do P3b no couro cabeludo é geralmente maior nas áreas parietais.[2] No entanto, usando uma configuração de 15 eletrodos com uma referência de lóbulo da orelha conectado e uma tarefa excêntrica (descrita abaixo), os pesquisadores também descobriram que a positividade aumentou ao se mover de locais frontais para parietais, e que as mulheres têm um aumento maior do que os homens.[11] Outra pesquisa, usando o Sistema Internacional 10-20 com uma referência de mastoide esquerda e uma tarefa excêntrica, mostrou que com o aumento da idade, a distribuição do P3b tende a mudar mais frontalmente.[10] Portanto, a distribuição exata pode depender da tarefa, bem como do gênero e da idade dos indivíduos.
Principais paradigmas
O P3b pode ser observado em uma variedade de contextos experimentais. Os paradigmas mais comuns apresentarão estímulos infrequentes e relevantes para a tarefa como uma forma de eliciar um P3b, ou empregarão duas tarefas ao mesmo tempo para usar o P3b como uma medida da carga de trabalho cognitiva. É claro que qualquer paradigma experimental no qual os participantes são instruídos a prestar atenção e avaliar estímulos deve eliciar um componente P3b, incluindo tarefas de atenção seletiva, tarefas de memória explícita e tarefas de busca visual (para revisões completas de paradigmas experimentais que foram usados para eliciar esse componente, veja Kok, 2001[12] e Verleger, 1997).[13]
Paradigmas excêntricos
Dois paradigmas clássicos são a tarefa oddball de dois estímulos e a tarefa oddball de três estímulos, a última das quais é usada para examinar P3b e P3a.[11] Em uma tarefa oddball clássica de dois estímulos, uma sequência de estímulos visuais é apresentada. Por exemplo, os sujeitos podem ver uma sequência de letras apresentadas uma de cada vez. Um estímulo "alvo" ou "excêntrico" menos frequente, como a letra T, é apresentado junto com estímulos "padrão" mais frequentes, como a letra S. O sujeito é normalmente instruído a responder de alguma forma (como com o pressionamento de um botão) apenas aos alvos e a ignorar os padrões. O P3b é normalmente observado em torno de 300 ms após cada apresentação do estímulo alvo (excêntrico).[2]
Uma tarefa excêntrica com três estímulos é exatamente igual à tarefa excêntrica com dois estímulos, exceto que, além dos alvos e padrões, um estímulo desviante e infrequente, como a letra "D", aparecerá. Esses estímulos são frequentemente conhecidos como padrões desviantes, pois não são o alvo da tarefa, mas diferem do padrão regular. Foi demonstrado que o P3b responde apenas a estímulos relevantes para a tarefa, ou aos alvos que estão sendo ativamente procurados (neste exemplo, a letra T). Portanto, o padrão desviante "D" não eliciará um P3b forte, pois não é relevante para a tarefa. No entanto, o padrão desviante ainda eliciará um potencial positivo anterior, geralmente mais alto em sítios frontais, conhecido como P3a. Ao contrário do P3b, o P3a se habitua com apresentações repetidas.[2]
Paradigmas de dupla tarefa
Outro conjunto de paradigmas usados para estudar o P3b são os paradigmas de tarefa dupla. Existem diversas variações do paradigma de tarefa dupla, e elas podem ser usadas para estudar a carga de trabalho cognitiva (ver Kok, 2001).[12] A carga de trabalho pode ser definida como a quantidade de recursos de processamento necessários para uma tarefa específica. Em um paradigma de tarefa dupla, os participantes recebem duas tarefas para executar simultaneamente; uma tarefa primária e uma tarefa secundária. Embora a tarefa primária possa ser de praticamente qualquer tipo, a tarefa secundária deve envolver algum paradigma P300 tradicional (por exemplo, uma tarefa excêntrica). Quando essas tarefas são executadas simultaneamente, esperaríamos ver uma redução na amplitude do P3b em resposta à tarefa secundária se a tarefa primária exigir alguns recursos de avaliação de estímulo. Além disso, presume-se que a extensão dessa redução reflita a quantidade de carga de trabalho associada à tarefa primária. De fato, deve haver uma relação recíproca entre as amplitudes da resposta do P3b provocada pelas tarefas primária e secundária, respectivamente. Se a tarefa primária for mais fácil (ou seja, exigir menos recursos de avaliação de estímulos), os participantes terão mais recursos disponíveis para dedicar à tarefa secundária. Por outro lado, se a tarefa primária for mais difícil (ou seja, exigir mais recursos de avaliação de estímulos), os participantes terão menos recursos disponíveis para dedicar à tarefa secundária.
Para dar um exemplo, os sujeitos podem executar uma tarefa primária, como rastrear um objeto visual em uma tela com um joystick, simultaneamente com uma tarefa secundária de contar mentalmente objetos estranhos em um fluxo auditivo. A dificuldade da tarefa primária é geralmente manipulada de várias maneiras, e os impactos dessas manipulações na resposta P3b à tarefa secundária são examinados. Por exemplo, em uma condição, os sujeitos podem rastrear o movimento unidimensional de um objeto (apenas para cima e para baixo) e, na condição mais difícil, eles podem ter que rastrear o movimento bidimensional (qualquer direção na tela do computador).[14] Uma manipulação motora como essa normalmente impactará os tempos de reação na tarefa secundária, mas não impactará a resposta P3b. No entanto, se você aumentar a demanda na memória de trabalho ou outros recursos cognitivos durante a tarefa primária, por exemplo, adicionando objetos à tela ou fazendo com que os sujeitos atenda seletivamente apenas uma parte da tela, a amplitude do P3b em resposta a objetos estranhos na tarefa secundária diminuirá. A quantidade que diminui pode ser uma medida de quantos recursos de memória de trabalho ou de avaliação de estímulos estão sendo usados pela tarefa primária.[12]
Em outra variação do paradigma de tarefa dupla, os sujeitos são apresentados a um fluxo visual de itens apresentados um de cada vez. Neste fluxo, há dois alvos, cada um dos quais requer uma resposta separada. A quantidade de tempo ou itens que separam os dois alvos é variada, e a amplitude do P3b em resposta ao segundo item é examinada. A amplitude diminuída da resposta do P3b ao segundo alvo seria esperada quando o primeiro alvo exigisse mais recursos de processamento ou memória de trabalho.[15]
Significado funcional: fatores que influenciam a amplitude
Variáveis cognitivas
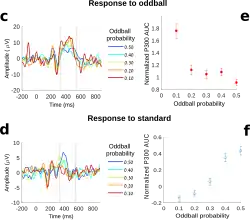
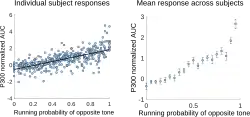
Donchin afirmou que quanto menos provável for o evento, maior será a amplitude do P3b.[3] Existem vários tipos de probabilidade que podem afetar a amplitude do P3b: probabilidade global, ou a frequência com que os alvos são relativos ao número de padrões (por exemplo, a amplitude do P3b é maior quando os alvos constituem 10 por cento dos estímulos do que quando os alvos constituem 20 por cento dos estímulos);[16] probabilidade local, ou a probabilidade dentro da sequência específica dos eventos (por exemplo, se um alvo seguiu um padrão ou outro alvo);[16] e probabilidade temporal, ou a frequência com que os alvos ocorrem dentro de um período de um minuto (se os padrões estão presentes ou não).[17]
Ray "Skip" Johnson, Jr., agora professor no Queens College, publicou um modelo triárquico de amplitude P300 em 1986.[18] Embora ele não se refira explicitamente a P3b neste artigo, a maioria de suas discussões se refere a P3b. Ele ofereceu três coisas que afetaram a amplitude - probabilidade subjetiva, significado do estímulo e transmissão de informações. Ele resumiu sua visão na seguinte fórmula: amplitude P300 = f[T x (1/P + M)], onde P é probabilidade subjetiva, M é significado do estímulo e T é informação transmitida.[18] Ele descreve a probabilidade subjetiva como probabilidade objetiva com o elemento adicional de julgamento humano de quão relevante um estímulo é para a tarefa e observa que a amplitude P300 está diretamente relacionada à quantidade de incerteza que é reduzida por um estímulo. Uma dependência intimamente relacionada entre o P300 e a probabilidade subjetiva foi mostrada por Levi-Aharoni et al.[1] que usaram uma representação difusa e comprimida da probabilidade do estímulo para explicar a variabilidade de teste único na magnitude da resposta P300. No entanto, também foi descoberto que a amplitude do P300 pode mudar na ausência de mudanças na probabilidade. O significado do estímulo, portanto, refere-se a variáveis que respondem pelo processamento de um estímulo que não estão relacionadas à probabilidade. O significado do estímulo abrange três variáveis independentes que podem ser manipuladas - complexidade da tarefa (quão difícil é uma tarefa ou quantas tarefas devem ser realizadas de uma vez), complexidade do estímulo (demanda perceptual ou quantas características relevantes do estímulo devem ser processadas - um rosto sendo mais complexo do que um ponto) e valor do estímulo (a significância ou, por exemplo, o valor monetário: quanto maior o valor, maior a amplitude do P300). A transmissão de informações é a proporção de informações de estímulo recebidas por uma pessoa em relação à quantidade de informações que o estímulo continha originalmente.[18] Existem manipulações externas e internas de transmissão de informação. Quando muita informação é perdida por razões externas, por exemplo porque o estímulo é mais difícil de discriminar ou perceber, a amplitude do P300 é menor. Manipulações internas são variações em quanta atenção os sujeitos são requeridos ou permitidos a dar ao estímulo. P3b requer atenção, e aumentar a dificuldade de manter a atenção diminuirá correspondentemente a amplitude do P3b. Para resumir, Johnson descreve que a probabilidade em muitos níveis, a relevância do estímulo para a tarefa, e a quantidade de informação que um estímulo transmite são todas variáveis que determinarão a amplitude do P3b.[18]
Mais recentemente, Albert Kok revisou a literatura sobre carga de trabalho cognitiva e concluiu que a amplitude do P3b depende das demandas sobre a capacidade cognitiva.[12] Em paradigmas de tarefa dupla como os descritos acima, os sujeitos são obrigados a executar uma tarefa primária e uma secundária. Quando a tarefa primária é mais exigente perceptual e cognitivamente, a amplitude do P3b em resposta a eventos estranhos na tarefa secundária é diminuída. Kok também apoia aspectos da teoria de Johnson, afirmando que a quantidade de atenção alocada a uma tarefa, a relevância do estímulo para a tarefa e a probabilidade do estímulo ajudarão a determinar qual será a amplitude do P3b.[12]
Consistentemente com os modelos de Johnson e Kok, um modelo da magnitude da resposta P300 foi proposto por Levi-Aharoni, Shriki e Tishby,[1] com base na estrutura Information Bottleneck[19] e um modelo de erro de predição por Rubin et al.[20] Seus resultados mostram que a variabilidade de teste único na magnitude da resposta P300 pode ser explicada por uma surpresa subjetiva que é construída a partir de representações de probabilidade comprimidas, dependentes da capacidade de memória alocada à tarefa e das características relevantes dos estímulos precedentes para prever o estímulo seguinte. De acordo com esse modelo, quanto maior for a capacidade de memória alocada, mais precisa será a representação do estímulo e maior será a surpresa que os desviantes da resposta devem provocar. Além disso, com base nessa dependência, eles propuseram um método para utilizar a resposta P300 individual para obter uma estimativa da capacidade de memória recente individual.
Gênero, aprendizagem e assimetrias
Outras variáveis foram encontradas para influenciar a amplitude P3b. Algumas pesquisas usando tarefas excêntricas indicaram que as mulheres têm amplitudes P3b maiores do que os homens, e que a amplitude aumenta mais movendo-se de áreas frontais para parietais.[11] Outras pesquisas descobriram que a aprendizagem em uma área de tópico pode afetar a amplitude P3b em tarefas relacionadas a essa área. Um estudo pegou um grupo de indivíduos e deu a alguns deles treinamento em progressões de acordes musicais padrão enquanto os outros permaneceram sem treinamento.[21] Todos receberam então sequências de acordes que continham violações. Os pesquisadores descobriram que aqueles que receberam treinamento prévio tiveram maiores amplitudes P3b em resposta a violações harmônicas em sequências musicais. Isso provavelmente ocorre porque aqueles com treinamento tinham mais experiência com as regras que governam os harmônicos e, portanto, têm um maior grau de expectativa para progressões de acordes e são mais sensíveis ao desvio.[21] Há também algumas evidências que sugerem que em todos os indivíduos, a amplitude P3b é distribuída assimetricamente sobre o couro cabeludo. A pesquisa mostrou que as amplitudes do P3b são sistematicamente maiores nos hemisférios frontal e central direito do que no esquerdo, embora haja algum debate sobre se isso se deve a causas estruturais (como espessura do crânio ou irregularidades cranianas) ou a causas cognitivas.[22]
Variáveis clínicas
Várias variáveis clínicas, como idade, doença, doença mental e uso de substâncias, foram estudadas em relação à amplitude do P3b. Muitos desses estudos não se referem explicitamente ao P3b, mas a maioria usa tarefas excêntricas e, portanto, provavelmente teriam provocado um P3b. Como observado anteriormente, a topografia da amplitude do P3b tende a mudar mais frontalmente com a idade,[10] mas a amplitude nessas áreas frontais não parece ser afetada pela idade.[23] Outro estudo usando uma tarefa excêntrica auditiva mostrou que a amplitude do P300 foi aumentada em indivíduos não medicados com doença de Parkinson leve quando comparados a controles saudáveis. O mesmo estudo também usou grupos mais velhos e mais jovens de indivíduos com doença de Parkinson e descobriu que a idade em si não teve efeito na amplitude.[24] Outros estudos mostraram que indivíduos com esquizofrenia têm uma redução acentuada na amplitude do P3b, sugerindo deficiências na memória de trabalho ou outro processamento.[25] O uso de substâncias também demonstrou afetar o P3b. Alguns estudos mostram que os alcoólatras têm amplitudes maiores de P300 em resposta a uma tarefa auditiva excêntrica, mas que os alcoólatras que têm muitos parentes alcoólatras têm amplitudes menores de P300 quando comparados aos controles.[26] Essa tendência de a amplitude de P3b ser mais fraca para aqueles com histórico familiar de alcoolismo pode ser mais geral, visto que outros transtornos por uso de substâncias e transtornos psiquiátricos relacionados mostram a mesma redução, geralmente antes do início do uso da substância.[27] Essa redução específica na amplitude de P3b pode ser devido a fatores neuromodulatórios fracos nos cérebros daqueles propensos a desenvolver problemas de uso de substâncias.[28] A amplitude de P300 também parece ser sensível a intervenções farmacológicas. Quando doze indivíduos saudáveis receberam lorazepam (um medicamento benzodiazepínico usado para tratar ansiedade e, às vezes, depressão) e foram solicitados a realizar uma tarefa excêntrica, sua amplitude de P300 foi reduzida.[29] Esses e outros estudos demonstraram que as condições que tendem a influenciar a cognição (como idade, doença, doença mental e uso de substâncias) afetam a amplitude do P3b ou sua distribuição.
Significado funcional: fatores que influenciam a latência
Em consonância com a visão de que o componente P3b reflete a resolução da incerteza, há evidências que sugerem que o tempo em que esse componente começa a aparecer no ERP (ou seja, sua latência) corresponde ao tempo em que a incerteza é resolvida. Por exemplo, Sutton et al. (1967)[6] realizaram um estudo no qual manipularam quando a incerteza poderia ser resolvida. Mais especificamente, os participantes foram apresentados a cliques simples ou duplos (auditivos) que variaram em intensidade. Em uma condição, os participantes foram solicitados a relatar o número de cliques que ouviram. Quando um clique duplo ocorreu, a resposta P3b ocorreu aproximadamente 300 ms após o segundo clique. Mais importante ainda, o tempo da resposta P3b foi quase idêntico quando apenas um único clique foi apresentado, sugerindo que esse componente foi gerado com base em quando o segundo clique pode ter ocorrido. De fato, quando a duração entre os dois cliques foi manipulada, o início do P3b foi atrasado pelo tempo exato entre eles (por exemplo, quando o segundo clique foi apresentado 500 ms após o primeiro, a resposta do P3b ocorreu em 800 ms). Em contraste, quando os participantes foram instruídos a responder com base na intensidade dos cliques, a resposta do P3b sempre ocorreu aproximadamente 300 ms após o primeiro clique. Presumivelmente, os participantes conseguiram determinar a intensidade dos cliques com base no primeiro; assim, o primeiro clique resolveu sua incerteza.
Se de fato a latência do componente P3b reflete o momento da resolução da incerteza, então pode-se esperar que a latência desse componente esteja intimamente relacionada à dificuldade da avaliação ou categorização. De fato, agora há ampla evidência para apoiar essa afirmação. Por exemplo, McCarthy e Donchin (1981)[30] apresentaram aos participantes matrizes 3 x 3, cada uma contendo a palavra 'ESQUERDA' ou a palavra 'DIREITA'. A tarefa deles era responder quando localizassem a palavra de direção na matriz, e a identidade da palavra determinava que tipo de resposta eles deveriam dar. McCarthy e Donchin descobriram que o componente P3b ocorria significativamente mais cedo quando o restante dos itens na matriz eram sinais numéricos (#), em relação a quando o restante dos itens eram letras aleatórias. Em essência, as letras aleatórias serviam como "ruído", o que fazia com que os participantes demorassem mais para identificar a palavra-alvo. Seguindo a mesma linha, descobriu-se que diversas outras manipulações de tarefas que se acredita influenciarem a dificuldade da avaliação ou categorização envolvida influenciam a latência do P3b (por exemplo, diminuindo a intensidade física dos estímulos; ver Verleger, 1997[13] para uma revisão). Coletivamente, essas descobertas sugerem que a latência do P3b reflete o tempo que os participantes levam para avaliar ou categorizar o estímulo em questão.
Dado que os índices de desempenho (como tempos de resposta) têm sido usados há muito tempo na psicologia cognitiva para estudar a duração e/ou o tempo de eventos mentais, pode-se perguntar se a latência do P3b simplesmente fornece um índice neural comparável desses mesmos processos. A pesquisa sugere que a latência do P3b é altamente correlacionada com os tempos de resposta quando os participantes são instruídos a priorizar a precisão em suas respostas, mas é menos correlacionada com os tempos de resposta quando os participantes são instruídos a priorizar a velocidade em suas respostas.[31] Esse padrão de resultados sugere que o P3b primário reflete os processos de avaliação de estímulos, enquanto os tempos de resposta são considerados como refletindo tanto a avaliação do estímulo quanto a seleção da resposta (mas para uma crítica dessa afirmação, veja Verleger, 1997).[13] Mais especificamente, quando os participantes são instruídos a priorizar a velocidade (ou seja, responder o mais rápido possível), eles podem responder antes que a avaliação do estímulo seja concluída (o que é consistente com o fato de que os participantes tendem a cometer mais erros nessas condições). Suporte adicional para esta conclusão vem da descoberta de que a latência do P3b não é afetada por manipulações experimentais que se acredita influenciarem os processos de seleção de resposta (por exemplo, compatibilidade estímulo-resposta), enquanto os tempos de reação são (por exemplo, McCarthy & Donchin, 1981).[30] Uma implicação importante desta conclusão é que o componente P3b pode ser usado para identificar o "locus de interferência" em muitos paradigmas cognitivos populares. Por exemplo, Luck (1998)[15] descobriu que a latência do P3b foi apenas ligeiramente atrasada durante o período refratário psicológico (PRP), sugerindo que a interferência no tempo de resposta neste paradigma reflete principalmente um atraso na seleção de resposta.
Outros fatores que influenciam a latência da resposta P3b incluem fatores relacionados à excitação fisiológica, como frequência cardíaca e consumo de cafeína,[9] bem como fatores relacionados à capacidade cognitiva, como idade e diferenças na rapidez com que os indivíduos podem alocar recursos de atenção.[2]
Teoria
Várias teorias foram apresentadas sobre quais processos cognitivos o P3b reflete. Donchin[3] propôs um "modelo de atualização de contexto". Este modelo postula que o cérebro gera constante e automaticamente hipóteses sobre o ambiente e o que está prestes a experimentar. Uma onda P300, incluindo um P3b, é gerada quando o cérebro recebe informações que indicam que ele precisa mudar essas hipóteses ou atualizar seu modelo mental do mundo. Em outras palavras, um P300 é eliciado sempre que há informações suficientes para indicar que o cérebro precisa atualizar a memória de trabalho.[3] Embora a explicação da "atualização de contexto" seja bem apoiada por pesquisas existentes, várias teorias alternativas foram propostas. Por exemplo, Verleger e colegas (2005)[32] propuseram que o componente P3b reflete um processo que medeia entre a análise perceptual e o início da resposta. Mais especificamente, esse processo emerge de um mecanismo cognitivo responsável por monitorar se a classificação de um estímulo é adequadamente traduzida em ação.[32] Esta teoria constitui um desafio direto à visão amplamente difundida de que o componente P3b reflete processos envolvidos na percepção, mas não na iniciação da resposta (ver Verleger, 1997).[13]
Outra teoria proposta por Kok[12] propôs que o P3b reflete mecanismos envolvidos na categorização de eventos, ou o processo que leva à decisão sobre se um estímulo externo corresponde ou não a uma representação interna de uma categoria ou estímulo específico. A categorização requer processos como atenção, percepção e memória de trabalho, todos os quais são conhecidos por afetar a amplitude do P3b (conforme revisado acima) e, portanto, este modelo integra as descobertas da pesquisa sobre o P3b. Kok também discute outro "modelo de correspondência de modelo", onde os sujeitos são obrigados a detectar um alvo e criar uma representação ou "modelo" do estímulo, e o P3b é mais forte quando o modelo é correspondido pelos estímulos apresentados. O modelo de correspondência de modelo é semelhante ao modelo de categorização de eventos e sugere que o P3b reflete processos que fundamentam a memória de reconhecimento (que também pode exigir memória de trabalho).[12] O modelo de categorização de eventos tem semelhanças com o modelo proposto por Verleger que sugere que o P3 é gerado durante o "fechamento" de um ciclo perceptual.[12] A versão cognitiva do modelo de Verleger sugere que o P3b é gerado quando é tomada uma decisão de que um estímulo pertence a uma categoria relevante para a tarefa.[33] Como Kok[12] resume, o P3b parece integrar processos que são necessários para identificar e combinar um estímulo com algum tipo de representação interna.
Origens neurais
Os geradores neurais do P3b são altamente debatidos. Estudos iniciais usando eletrodos implantados no cérebro indicaram que a formação do hipocampo pode gerar o P300.[33] No entanto, trabalhos posteriores descobriram que se o hipocampo for lesionado ou danificado, o P300 ainda é gerado e nenhuma diferença confiável em sua amplitude ou latência é observada. Pesquisas subsequentes descobriram que a lesão da junção do lobo temporal-parietal afetou significativamente a produção do P300, indicando que essa área pode conter um ou mais geradores do P3b.[33] Isso sugere que o P3b pode indicar algum tipo de via de circuito entre as áreas cerebrais frontal e temporal/parietal.[2] Um gerador temporal-parietal seria lógico, porque o P3b parece ser eliciado quando as ativações de recursos atencionais promovem a memória de trabalho e outros processos nas áreas temporais-parietais.[2] Outras pesquisas de EEG utilizando técnicas de modelagem de fonte, juntamente com pesquisas usando métodos alternativos de imagem cerebral (por exemplo, fMRI, MEG), gravações intracranianas e pacientes com lesão cerebral, também indicaram que o componente P3b se origina da ativação nos lobos parietal e temporal do córtex cerebral.[2] Há também algumas evidências de que a ativação de certas estruturas límbicas, como o córtex cingulado anterior, pode contribuir para o componente P3b.[12]
Ainda não se sabe quais sistemas neurotransmissores são responsáveis pela geração de P3b. A região temporoparietal é densamente povoada por entradas de norepinefrina,[2] e há algumas evidências de que o sistema de norepinefrina do locus coeruleus pode ser responsável pela geração de P3b.[2] Como muitos desses estudos foram conduzidos usando animais, mais pesquisas são necessárias para determinar os neurotransmissores responsáveis pela geração de P3b.
Referências
- ↑ a b c d e f g Levi-Aharoni H, Shriki O, Tishby N (Fevereiro de 2020). «Surprise response as a probe for compressed memory states». PLOS Computational Biology. 16 (2). Bibcode:2020PLSCB..16E7065L. PMC 7018098
 . PMID 32012146. doi:10.1371/journal.pcbi.1007065
. PMID 32012146. doi:10.1371/journal.pcbi.1007065
- ↑ a b c d e f g h i j k l m Polich J (Outubro de 2007). «Updating P300: an integrative theory of P3a and P3b». Clinical Neurophysiology. 118 (10): 2128–48. PMC 2715154. PMID 17573239. doi:10.1016/j.clinph.2007.04.019
- ↑ a b c d e Donchin E (Setembro de 1981). «Presidential address, 1980. Surprise!...Surprise?». Psychophysiology. 18 (5): 493–513. PMID 7280146. doi:10.1111/j.1469-8986.1981.tb01815.x
- ↑ a b c Chapman RM, Bragdon HR (Setembro de 1964). «Evoked Responses to Numerical and Non-Numerical Visual Stimuli while Problem Solving». Nature. 203 (4950): 1155–7. Bibcode:1964Natur.203.1155C. PMID 14213667. doi:10.1038/2031155a0
- ↑ a b Sutton S, Braren M, Zubin J, John ER (Novembro de 1965). «Evoked-potential correlates of stimulus uncertainty». Science. 150 (3700): 1187–8. Bibcode:1965Sci...150.1187S. PMID 5852977. doi:10.1126/science.150.3700.1187
- ↑ a b c Sutton S, Tueting P, Zubin J, John ER (Março de 1967). «Information delivery and the sensory evoked potential». Science. 155 (3768): 1436–9. Bibcode:1967Sci...155.1436S. PMID 6018511. doi:10.1126/science.155.3768.1436
- ↑ Bashore TR, van der Molen MW (1991). «Discovery of the P300: a tribute». Biological Psychology. 32 (2–3): 155–71. PMID 1790268. doi:10.1016/0301-0511(91)90007-4
- ↑ Squires NK, Squires KC, Hillyard SA (Abril de 1975). «Two varieties of long-latency positive waves evoked by unpredictable auditory stimuli in man». Electroencephalography and Clinical Neurophysiology. 38 (4): 387–401. CiteSeerX 10.1.1.326.332. PMID 46819. doi:10.1016/0013-4694(75)90263-1
- ↑ a b Polich J, Kok A (Outubro de 1995). «Cognitive and biological determinants of P300: an integrative review». Biological Psychology. 41 (2): 103–46. PMID 8534788. doi:10.1016/0301-0511(95)05130-9
- ↑ a b c Fjell AM, Walhovd KB, Reinvang I (Setembro de 2005). «Age-dependent changes in distribution of P3a/P3b amplitude and thickness of the cerebral cortex». NeuroReport. 16 (13): 1451–4. PMID 16110270. doi:10.1097/01.wnr.0000177011.44602.17
- ↑ a b c Conroy MA, Polich J (2007). «Normative variation of P3a and P3b from a large sample (N=120): Gender, topography, and response time». Journal of Psychophysiology. 21 (1): 22–32. doi:10.1027/0269-8803.21.1.22
- ↑ a b c d e f g h i j Kok A (Maio de 2001). «On the utility of P3 amplitude as a measure of processing capacity». Psychophysiology. 38 (3): 557–77. PMID 11352145. doi:10.1017/S0048577201990559
- ↑ a b c d Verleger R (Março de 1997). «On the utility of P3 latency as an index of mental chronometry». Psychophysiology. 34 (2): 131–56. PMID 9090263. doi:10.1111/j.1469-8986.1997.tb02125.x
- ↑ Isreal JB, Chesney GL, Wickens CD, Donchin E (Maio de 1980). «P300 and tracking difficulty: evidence for multiple resources in dual-task performance». Psychophysiology. 17 (3): 259–73. PMID 7384376. doi:10.1111/j.1469-8986.1980.tb00146.x
- ↑ a b Luck SJ (1998). «Sources of dual-task interference: Evidence from human electrophysiology». Psychological Science. 9 (3): 223–227. doi:10.1111/1467-9280.00043
- ↑ a b Duncan-Johnson CC, Donchin E (Setembro de 1977). «On quantifying surprise: the variation of event-related potentials with subjective probability». Psychophysiology. 14 (5): 456–67. PMID 905483. doi:10.1111/j.1469-8986.1977.tb01312.x
- ↑ Polich J, Margala C (Fevereiro de 1997). «P300 and probability: comparison of oddball and single-stimulus paradigms». International Journal of Psychophysiology. 25 (2): 169–76. PMID 9101341. doi:10.1016/S0167-8760(96)00742-8
- ↑ a b c d Johnson R (Julho de 1986). «A triarchic model of P300 amplitude». Psychophysiology. 23 (4): 367–84. PMID 3774922. doi:10.1111/j.1469-8986.1986.tb00649.x
- ↑ Tishby N, Pereira FC, Bialek W (Setembro de 1999). The Information Bottleneck Method (PDF). The 37th annual Allerton Conference on Communication, Control, and Computing. pp. 368–377
- ↑ Rubin J, Ulanovsky N, Nelken I, Tishby N (Agosto de 2016). Theunissen FE, ed. «The Representation of Prediction Error in Auditory Cortex». PLOS Computational Biology. 12 (8). Bibcode:2016PLSCB..12E5058R. PMC 4973877. PMID 27490251. doi:10.1371/journal.pcbi.1005058
- ↑ a b Carrión RE, Bly BM (Setembro de 2008). «The effects of learning on event-related potential correlates of musical expectancy». Psychophysiology. 45 (5): 759–75. PMID 18665861. doi:10.1111/j.1469-8986.2008.00687.x
- ↑ Alexander JE, Porjesz B, Bauer LO, Kuperman S, Morzorati S, O'Connor SJ, et al. (Setembro de 1995). «P300 hemispheric amplitude asymmetries from a visual oddball task». Psychophysiology. 32 (5): 467–75. PMID 7568641. doi:10.1111/j.1469-8986.1995.tb02098.x
- ↑ Ortiz T, Martin-Loeches M, Vila E (1990). «Frontal lobes and aging effect on the P300 component of the auditory event-related potentials». Applied Psychology: An International Review. 39 (3): 323–330. doi:10.1111/j.1464-0597.1990.tb01057.x
- ↑ Green J, Woodard JL, Sirockman BE, Zakers GO, Maier CL, Green RC, Watts RL (Janeiro de 1996). «Event-related potential P3 change in mild Parkinson's disease». Movement Disorders. 11 (1): 32–42. PMID 8771065. doi:10.1002/mds.870110108
- ↑ Friedman D, Squires-Wheeler E (1994). «Event-related potentials (ERPs) as indicators of risk for schizophrenia». Schizophrenia Bulletin. 20 (1): 63–74. PMID 8197422. doi:10.1093/schbul/20.1.63
- ↑ Cohen HL, Wang W, Porjesz B, Begleiter H (Abril de 1995). «Auditory P300 in young alcoholics: regional response characteristics». Alcoholism: Clinical and Experimental Research. 19 (2): 469–75. CiteSeerX 10.1.1.418.6561. PMID 7625584. doi:10.1111/j.1530-0277.1995.tb01533.x
- ↑ Iacono WG, Carlson SR, Malone SM, McGue M, et al. (Agosto de 2002). «P3 event-related potential amplitude and the risk for disinhibitory disorders in adolescent boys». Archives of General Psychiatry. 59 (8): 750–7. PMID 12150652. doi:10.1001/archpsyc.59.8.750
- ↑ Burwell SJ, Malone SM, Bernat EM, Iacono WG, et al. (Outubro de 2014). «Does electroencephalogram phase variability account for reduced P3 brain potential in externalizing disorders?». Clinical Neurophysiology. 125 (10): 2007–15. PMC 4156932. PMID 24656843. doi:10.1016/j.clinph.2014.02.020
- ↑ Pooviboonsuk P, Dalton JA, Curran HV, Lader MH (1996). «The effect of single doses of lorazepam on event-related potentials and cognitive function». Human Psychopharmacology. 11 (3): 241–252. doi:10.1002/(SICI)1099-1077(199605)11:3<241::AID-HUP795>3.0.CO;2-0
- ↑ a b McCarthy G, Donchin E (Janeiro de 1981). «A metric for thought: a comparison of P300 latency and reaction time». Science. 211 (4477): 77–80. Bibcode:1981Sci...211...77M. PMID 7444452. doi:10.1126/science.7444452
- ↑ Kutas M, McCarthy G, Donchin E (Agosto de 1977). «Augmenting mental chronometry: the P300 as a measure of stimulus evaluation time» (PDF). Science. 197 (4305): 792–5. Bibcode:1977Sci...197..792K. PMID 887923. doi:10.1126/science.887923
- ↑ a b Verleger R, Jaśkowski P, Wascher E (2005). «Evidence for an integrative role of P3b in linking reaction to perception». Journal of Psychophysiology. 19 (3): 165–181. doi:10.1027/0269-8803.19.3.165
- ↑ a b c Polich J, Criado JR (Maio de 2006). «Neuropsychology and neuropharmacology of P3a and P3b». International Journal of Psychophysiology. 60 (2): 172–85. PMID 16510201. doi:10.1016/j.ijpsycho.2005.12.012