N1 visual
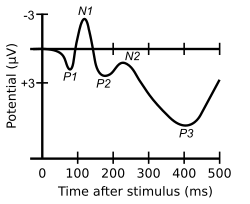
O N1 visual é um potencial evocado visual, um tipo de potencial elétrico relacionado a eventos (ERP), que é produzido no cérebro e registrado no couro cabeludo. O N1 é assim chamado para refletir a polaridade e o tempo típico do componente. O "N" indica que a polaridade do componente é negativa em relação a uma referência mastoide média. O "1" originalmente indicava que era o primeiro componente negativo, mas agora indexa melhor o pico típico deste componente, que é em torno de 150 a 200 milissegundos após o estímulo. A deflexão do N1 pode ser detectada na maioria dos locais de gravação, incluindo os locais dos eletrodos occipital, parietal, central e frontal.[1] Embora o N1 visual seja amplamente distribuído por todo o couro cabeludo, ele atinge o pico mais cedo nas regiões frontais do que nas posteriores do couro cabeludo,[1][2] sugerindo correlatos neurais e/ou cognitivos distintos.[3] O N1 é provocado por estímulos visuais e faz parte do potencial evocado visual – uma série de deflexões de voltagem observadas em resposta a inícios, deslocamentos e mudanças visuais. Tanto o hemisfério direito quanto o esquerdo geram um N1, mas a lateralidade do N1 depende se um estímulo é apresentado centralmente, lateralmente ou bilateralmente. Quando um estímulo é apresentado centralmente, o N1 é bilateral. Quando apresentado lateralmente, o N1 é maior, anterior e contralateral ao campo visual do estímulo. Quando dois estímulos visuais são apresentados, um em cada campo visual, o N1 é bilateral. No último caso, a assimetria do N1 é modulada pela atenção.[4] Além disso, sua amplitude é influenciada pela atenção seletiva e, portanto, tem sido usada para estudar uma variedade de processos atencionais.[5][6]
História
Embora o N1 seja um componente visual precoce que faz parte da resposta normal à estimulação visual, ele foi estudado mais extensivamente com relação à sua sensibilidade à atenção seletiva. Estudos iniciais com foco na modulação da amplitude do N1 com relação à atenção encontraram evidências limitadas para efeitos de atenção do N1. No entanto, a incerteza sobre a relação entre a amplitude do N1 e a atenção foi resolvida pelo estudo inovador de Haider, Spong e Lindsley (1964) no qual os níveis de atenção foram encontrados sistematicamente relacionados à variação na amplitude do N1. Especificamente, Haider et al. (1964) empregaram uma tarefa de vigilância que requer discriminação visual e resposta para garantir que os participantes prestassem atenção aos estímulos, em vez de observar passivamente as imagens visuais. Os participantes observaram uma série de flashes de luz e foram instruídos a responder com um pressionamento de botão para diminuir os flashes. Esses flashes fracos foram intercalados com flashes mais brilhantes que não exigiram uma resposta. O experimento durou aproximadamente 100 minutos e, como é típico desse tipo de tarefa de vigilância, a precisão na resposta aos flashes fracos diminuiu ao longo do tempo, o que é indicativo do declínio da atenção ao longo do experimento. É importante ressaltar que a amplitude do N1 variou sistematicamente com a resposta aos flashes fracos. À medida que a precisão e a atenção diminuíam, a amplitude do N1 diminuía, sugerindo que a amplitude do N1 está intimamente ligada aos níveis de atenção.[7]
Estudos subsequentes empregando diferentes manipulações de atenção encontraram resultados semelhantes, fornecendo suporte adicional para a ligação entre o N1 e a atenção. Em um estudo, os sujeitos direcionaram a atenção para diferentes tipos de estímulos visuais, e a amplitude do N1 para os estímulos visuais variou de acordo com se eles foram atendidos. Mais especificamente, o N1 foi maior para estímulos que foram atendidos versus aqueles que foram ignorados.[8] Um estudo posterior de Van Voorhis e Hillyard (1977)[9] examinou mudanças de amplitude no N1 durante uma tarefa na qual flashes de luz foram entregues simultaneamente ao campo visual esquerdo ou direito em sequências aleatórias independentes. Os sujeitos foram instruídos a atender à esquerda, atender à direita ou atender a ambos os campos. O aprimoramento do N1 no local occipital foi encontrado quando a atenção foi direcionada ao campo no qual os flashes de luz foram entregues. Em comparação, o N1 foi menor para flashes que ocorreram no campo oposto ao foco de atenção. Quando a atenção foi dividida entre os campos esquerdo e direito, a amplitude do N1 foi intermediária. Assim, as informações visuais em locais atendidos pareceram ser amplificadas. A modulação relacionada à atenção do N1 produziu evidências de atenção visual seletiva semelhante ao efeito de atenção descoberto na modalidade auditiva, na qual o N100 auditivo varia de acordo com a atenção seletiva dentro do campo auditivo.
Principais paradigmas
- Paradigma de Filtragem
Após a constatação de que a amplitude do N1 variava de acordo com os níveis de atenção, pesquisadores se interessaram em como estímulos idênticos eram percebidos quando atendidos ou não. Um paradigma experimental, às vezes chamado de Paradigma da Filtragem, foi desenvolvido para avaliar como a atenção influencia a percepção de estímulos. No Paradigma da Filtragem, os participantes são instruídos a concentrar sua atenção no campo visual direito ou esquerdo de uma tela de computador. O campo visual é tipicamente contrabalanceado dentro dos sujeitos ao longo dos ensaios ou blocos experimentais. Assim, para o primeiro conjunto de ensaios, os participantes podem prestar atenção ao campo visual direito, mas posteriormente podem prestar atenção ao campo visual esquerdo. Dentro de cada ensaio e entre os campos visuais, os participantes são apresentados aos mesmos estímulos, por exemplo, flashes de luz com duração variável. Os participantes são informados de que, quando um estímulo específico, como um flash de luz de curta duração, chamado de alvo, aparece no campo visual que estão observando, eles devem responder pressionando um botão. O número de alvos dentro de cada campo visual é menor do que o número de não alvos, e os participantes também são instruídos a ignorar o outro campo visual e a não responder aos alvos apresentados naquele campo visual. Quando os alvos no campo visual atendido são comparados aos alvos no campo visual não atendido, os alvos não atendidos provocam um N1 menor do que os alvos atendidos, sugerindo que a atenção atua como um mecanismo de ganho sensorial que aumenta a percepção de estímulos atendidos (vs. não atendidos).[5][6][9][10]
- Paradigma de Indicação Visuoespacial
Nos Paradigmas de Indicação Visuoespacial, a atenção é direcionada para uma área da tela do computador, mas a validade da indicação em relação à apresentação do estímulo-alvo varia. Assim, esse paradigma fornece insights sobre como colocar a atenção no local correto ou incorreto influencia a amplitude do N1. Por exemplo, os participantes são apresentados a um conjunto visual no qual há quatro caixas nos cantos superior e inferior direito e esquerdo da tela do computador. No primeiro quadro da exibição visual, eles são instruídos a fixar a atenção em uma pequena linha pontilhada no centro da tela do computador. Para preparar os participantes para localizar a indicação, segue-se um quadro de advertência no qual a linha pontilhada é substituída por uma cruz. O quadro de advertência é seguido pelo quadro de indicação, no qual uma seta aponta na direção de um ou de todos os quatro quadrados. Em alguns casos, a indicação é precisa e aponta para o quadrado no qual o alvo será apresentado. Em outros casos, a indicação é imprecisa e aponta para o quadrado no qual o alvo não será apresentado. Nos casos restantes, é apresentada uma dica neutra que aponta na direção de todos os quadrados. Em seguida, é exibido um quadro-alvo no qual um pequeno ponto aparece em um dos quatro quadrados. No último quadro, uma seta aponta para um dos quatro quadrados e os participantes respondem com um pressionamento de botão se a dica apareceu no quadrado. A amplitude do N1 varia em relação aos ensaios com indicação precisa, com indicação imprecisa e com indicação neutra. Nos ensaios em que a atenção foi direcionada para o quadrado em que o alvo foi apresentado (ensaios com indicação precisa), a amplitude do N1 é maior do que em a) ensaios em que a atenção foi direcionada para todos os quadrados (ensaios com indicação neutra) e b) ensaios em que a atenção foi direcionada para o quadrado errado (ensaios com indicação imprecisa), sugerindo que a amplitude do N1 representa um benefício para colocar a atenção no local correto.[11]
Fatores que influenciam a amplitude e a latência
A amplitude, ou o tamanho, do N1 é medido tomando-se a voltagem média dentro da janela que normalmente envolve o N1 (cerca de 150 a 200 ms após o estímulo). Como o N1 é um componente negativo, amplitudes "maiores" correspondem a ser mais negativas, enquanto amplitudes "menores" correspondem a ser menos negativas.
Pesquisas sugeriram que a amplitude do N1 é afetada por certos parâmetros visuais, incluindo a angularidade do estímulo e a luminância, ambos diretamente relacionados ao tamanho do N1.[12][13] A amplitude do N1 também é maior em resposta a estímulos em locais atendidos vs. não atendidos. Por outro lado, a amplitude diminui quando o intervalo interestímulo (ou seja, a quantidade de tempo entre apresentações sucessivas de estímulos) é aumentado para estímulos em locais atendidos.[14] Os efeitos de amplitude no N1 estão ausentes durante tarefas simples de Tempo de Reação, que exigem apenas que os sujeitos façam uma resposta rápida aos estímulos.[1] Essa descoberta sugere que o N1 está ligado a processos de discriminação visual.
Pesquisadores interessados em entender os efeitos de seleção da atenção têm se interessado especialmente na variação de amplitude do N1 porque acredita-se que as diferenças de amplitude representam um mecanismo de controle de ganho (veja Paradigma de Filtragem acima). Por exemplo, porque a amplitude do N1 para alvos em campos visuais não atendidos é menor do que para alvos em campos visuais atendidos, acredita-se que a atenção serve para amplificar o processamento de entradas sensoriais de locais atendidos e suprimir entradas sensoriais de locais não atendidos.[5][6] Assim, as diferenças de amplitude no N1 são úteis para fornecer evidências de se a atenção serve para selecionar certos tipos de estímulos sensoriais para processamento posterior.
Um dos fatores que influenciam a latência do N1 é o esforço de processamento: a latência do N1 aumenta à medida que o esforço de processamento também aumenta.[15] Especificamente, a latência parece aumentar durante tarefas que são significativamente complexas ou difíceis e, portanto, exigem maior atenção ativa ou esforço. Por exemplo, as latências de início, pico e deslocamento do N1 ocorrem significativamente mais cedo em resposta a estímulos em movimento em uma tarefa de detecção simples versus uma tarefa de identificação.[16] O N1 também é sensível à manipulação da intensidade de um estímulo visual. A latência de pico do N1 é encurtada à medida que o brilho dos flashes de estímulo aumenta.[17] Portanto, parece que a latência do N1 é afetada por fatores perceptivos, como a intensidade do flash, bem como o nível de demanda de atenção ou esforço de processamento.
- Cor e movimento
Diferenças de amplitude no N1 forneceram evidências de que a atenção permite uma análise mais extensa de informações visuais, como cor e movimento. Por exemplo, em um Paradigma de Filtragem (veja a descrição acima), os participantes foram instruídos a identificar alvos com base na cor ou no movimento. Em alguns casos, os participantes foram instruídos a prestar atenção a um lado do campo visual, enquanto em outros casos a atenção dos participantes não estava focada em um lado do campo visual. Foi descoberto que a amplitude do N1 era maior para alvos da cor e movimento corretos quando os participantes eram instruídos a prestar atenção a um lado do campo visual em comparação quando não eram instruídos a fazê-lo. Essas descobertas sugerem que a atenção a um local específico serve para facilitar o processamento posterior de informações visuais e suprimir o processamento visual posterior em locais desacompanhados.[18]
- Objetos e localização
Embora a atenção espacial tenha se mostrado única na seleção de informações perceptuais que serão processadas posteriormente, os objetos também se mostraram importantes na filtragem de informações para processamento posterior. Por exemplo, em um Paradigma de Filtragem (veja acima), retângulos foram apresentados em ambos os lados do campo visual. Os participantes foram instruídos a prestar atenção a um lado do campo visual e aos 50% superiores do objeto dentro desse campo visual. O alvo era uma região sombreada do canto superior direito; no entanto, alvos semelhantes foram apresentados na metade inferior desassistida do objeto no campo visual assistido e nas metades superior e inferior do objeto no campo visual desassistido. Como esperado, ao comparar alvos no campo visual assistido com alvos no campo visual desassistido, descobriu-se que a amplitude do N1 era maior para objetos assistidos (vs. desassistidos). Além disso, embora a amplitude do N1 tenha sido maior para alvos no campo visual assistido e na parte assistida do objeto, a amplitude do N1 para alvos na parte não assistida do objeto assistido foi maior do que a amplitude do N1 para alvos a uma distância equivalente do locus de atenção, mas em um objeto não assistido. Esses resultados fornecem evidências de que, embora a atenção espacial sirva como um mecanismo de seleção para processamento posterior, a atenção espacial pode se espalhar entre objetos e influenciar o processamento perceptual posterior.[19]
- Estímulos emocionais
Mais recentemente, a pesquisa sobre o N1 expandiu-se para o processamento de estímulos socialmente relevantes. A atenção é especialmente relevante para o processamento de estímulos emocionais porque se acredita que estímulos emocionais (vs. estímulos não emocionais) recebem atenção preferencial e processamento perceptual. A pesquisa ERP tem sido útil para entender como a emoção se relaciona com a atenção porque o N1 fornece um meio de examinar a importância da emoção na captura de recursos atencionais. Vários estudos, usando uma variedade de paradigmas, descobriram que estímulos emocionais são influentes na captura de atenção. Por exemplo, em um estudo, tanto estímulos que foram positivamente valenciados (por exemplo, pessoa nua do sexo oposto) quanto negativamente valenciados (por exemplo, lobo rosnando) mostraram provocar maiores amplitudes de N1 do que estímulos de valência neutra (por exemplo, relógio de pulso).[20] Da mesma forma, descobriu-se que a valência de estímulos interpessoais influencia a amplitude do N1. Descobriu-se que estímulos positivos (por exemplo, rostos sorridentes) e estímulos negativos (por exemplo, rostos tristes) provocam um N1 maior do que estímulos neutros (por exemplo, rostos neutros).[21] Essas descobertas apoiam a afirmação de que os estímulos emocionais são mais eficazes na captura de recursos de atenção do que os estímulos não emocionais.
Processos de atenção
O grande corpus de estudos focados em fatores que modulam a amplitude do N1 visual forneceu uma riqueza de evidências sugerindo que, embora o N1 visual seja um componente sensorial evocado por qualquer estímulo visual, ele também reflete um benefício da alocação correta de recursos atencionais e que é uma manifestação de um importante mecanismo sensorial de controle da atenção. Quando a atenção é focada em áreas do campo visual nas quais informações relevantes são apresentadas (vs. distribuídas uniformemente pelo campo visual ou focadas em uma área na qual informações relevantes não são apresentadas), a amplitude do N1 é maior e indica um benefício da alocação correta de recursos atencionais.[22] Além disso, acredita-se que a amplitude do N1 represente um mecanismo de controle de ganho sensorial porque focar a atenção em uma área do campo visual serve para aumentar a amplitude do N1 para informações perceptuais relevantes apresentadas naquele campo (vs. o outro campo visual) e, portanto, facilita o processamento perceptual posterior de estímulos.[5][6] Esta descoberta apoia o Modelo de Seleção Precoce da Atenção, que afirma que a atenção atua (ou seja, filtra informações) em um conjunto de estímulos no início do fluxo de processamento de informações.
Além disso, pesquisas sobre o N1 visual sugerem que a atenção espacial e a objetos serve como um mecanismo de seleção precoce que influencia a seleção de outras características perceptivas (por exemplo, cor, movimento) para processamento posterior. A amplitude do N1 é maior para características perceptivas em locais atendidos (vs. não atendidos) e em objetos atendidos (vs. não atendidos), fornecendo evidências de que as características perceptivas são selecionadas para processamento perceptivo posterior apenas se estiverem em locais atendidos ou em objetos atendidos.[18][19]
Por fim, o N1 visual também foi interpretado como reflexo de um processo de discriminação que ocorre dentro do locus de atenção. Em comparação com condições que simplesmente exigem uma resposta, o componente N1 é intensificado em condições que exigem uma diferenciação entre classes de estímulos. Esse efeito é semelhante para discriminações baseadas em cor e forma, independentemente do nível de dificuldade da discriminação. O N1 pode, portanto, refletir um mecanismo de discriminação aplicado a uma área espacial atendida.[23]
Fontes neurais
Identificar as fontes neurológicas dos componentes do ERP com base na distribuição topográfica do N1 no couro cabeludo é especialmente difícil porque o número de fontes potenciais (chamadas de dipolos), orientações e magnitudes que podem produzir a distribuição topográfica do N1, como qualquer outro componente do ERP, é teoricamente infinito. Este problema de trabalhar a partir da distribuição topográfica dos componentes do ERP para identificar fontes neurais é chamado de problema inverso.[24] Embora os geradores neurais do N1 não sejam definitivamente conhecidos,[10] evidências sugerem que o N1 não se origina no córtex visual primário, mas sim de múltiplos geradores no córtex occipito-parietal, occipito-temporal e (possivelmente) frontal.[25]
Referências
- ↑ a b c Mangun, G.R., & Hillyard, S.A, (1991). Modulations of sensory-evoked brain potentials indicate changes in perceptual processing during visual-spatial priming. Journal of Experimental Psychology: Human perception and performance, 17(4): 1057-1074.
- ↑ Ciesielski, K.T., & French, C.N. (1989). Event-related potentials before and after training: Chronometry and lateralization of visual N1 and N2. Biological Psychology, 28: 227-238.
- ↑ Makeig, S., Westerfield, M., Townsend, J., Jung, T., Courchesne, E., & Sejnowski, T.J. (1999). Functionally independent components of early event-related potentials in a visual spatial attention task. Royal Society, 354: 1135-1144.
- ↑ Wascher, E., Hoffman, S., Sanger, J., Grosjean, M. (2009). Visuo-spatial processing and the N1 component of the ERP. Psychophysiology, 46: 1270–1277.
- ↑ a b c d Luck, S. J., Woodman, G. E., and Vogel, E. K. (2000). Event-related potential studies of attention. Trends in Cognitive Sciences, 4, 432-440.
- ↑ a b c d Rugg, M.D., Milner, A.D., Lines, C.R., & Phalp, R. (1987). Modulations of visual event-related potentials by spatial and non-spatial visual selective attention. Neuropsychologia, 25, 85-96.
- ↑ Haider, M., Spong, P., & Lindsley, D.B. (1964). Attention, vigilance, and cortical evoked-potentials in humans, Science, 145, 180-182.
- ↑ Eason, R.G., Harter, M.R., & White, T.C. (1969). Effects of attention and arousal on visually evoked cortical potentials and reaction time in man. Physiology and Behavior, 4(3): 283-289.
- ↑ a b Van Voorhis, & Hillyard, S.A. (1977). Visual evoked potentials and selective attention to points in space. Perception and Psychophysics, 22(1): 54-62.
- ↑ a b Naatanen, R. & Michie, P.T. Early selective-attention effects of the evoked potential: A critical review and reinterpretation. (1979). Biological Psychology 8: 81-136.
- ↑ Luck, S. J., Hillyard, S.A., Mouloua, M., Woldorff, M.G., Clark, V.P., & Hawkins, H.L. (1994). Effects of spatial cuing on luminance detectability: Psychophysical and electrophysiological evidence for early selection. Journal of Experimental Psychology: Human Perception and Performance, 20, 887-904.
- ↑ Ito, M., Sugata, T., Kuwabara, H., Wu, C., & Kojima, K. (1999). Effects of angularity of the figures with sharp and round corners on visual evoked potentials. Japanese Psychological Research, 41(2): 91-101.
- ↑ Johannes, S., Munte, T.F., Heinze, H.J., & Mangun, G.R. (2003). Luminance and spatial attention effects on early visual processing. Cognitive Brain Research, 2(3): 189-205.
- ↑ Luck, S.J., Heinze, H.J., Mangun, G.R., & Hillyard, S.A. (1990). Visual event-related potentials index focused attention within bilateral stimulus arrays: II. Functional dissociation of P1 and N1 components. Electroencephalography & Clinical Neurophysiology, 75(6): 528-542.
- ↑ Callaway, E., & Halliday, R. (1982). The effect of attentional effort on visual evoked potential N1 latency. Psychiatry Research, 7: 299-308.
- ↑ Fort, A., Besle, J., Giard, M., & Pernier, J. (2005). Task-dependent activation latency in human visual extrastriate cortex. Neuroscience Letters, 379(2): 144-148.
- ↑ Carillo-de-la-Peña, M., Holguín, S. R., Corral, M., & Cadaveira, F. (1999). The effects of stimulus intensity and age on visual-evoked potentials (VEPs) in normal children. Psychophysiology, 36(6): 693-698.
- ↑ a b Anllo-Vento, L. & Hillyard, S.A. (1996). Selective attention to the color and direction of moving stimuli: Electrophysiological correlates of hierarchical feature selection. Perception & Psychophysics, 58, 191-206.
- ↑ a b Martinez, A., Teder-Salejarvi, W., Vasquez, M., Molholm, S., Foxe, J.J., Javitt, D.C., Di Russo, F., Worden, M.S., & Hillyard, S.A. (2006). Objects are highlighted by spatial attention. Journal of Cognitive Neuroscience, 18, 298-310.
- ↑ Carretié, L., Hinojosa, J.A., Martín-Loeches, M., Mercado, F., & Tapia, M. (2004). Automatic attention to emotional stimuli: Neural correlates, Human Brain Mapping, 22, 290-299.
- ↑ Foti, D., Hajcak, G., & Dien, J. (2009). Differentiating neural responses to emotional pictures: Evidence from temporal-spatial PCA. Psychophysiology, 46, 521-530.
- ↑ Luck, S.J, Hillyard, S.A., Mouloua, M., Woldorff, M.G., Clark, V.P., & Hawkins, H.L. (1994). Effects of spatial cuing on luminance detectability: Psychophysical and electrophysiological evidence for early selection. Journal of Experimental Psychology: Human Perception and Performance, 20, 887-904.
- ↑ Vogel, E.K., & Luck, S.J. (2000). The visual N1 component as an index of a discrimination process. Psychophysiology, 37: 190-203.
- ↑ Luck, S.J. (2005). An Introduction to the Event-Related Potential Technique. Cambridge, Mass.: The MIT Press.
- ↑ Clark, V.P., Fan, S., & Hillyard, S.A. (1995). Identification of early visual evoked potential generators by retinotopic and topographic analyses. Human Brain Mapping, 2, 170-187.