Memória espacial

Na psicologia cognitiva e na neurociência, a memória espacial é uma forma de memória responsável pelo registro e recuperação de informações necessárias para planejar um trajeto até um local e para recordar a localização de um objeto ou a ocorrência de um evento.[1] A memória espacial é essencial para a orientação no espaço.[2][3] A memória espacial também pode ser dividida em memória espacial egocêntrica e alocêntrica.[4]
A memória espacial de uma pessoa é necessária para navegar em uma cidade familiar. A memória espacial de um rato é necessária para aprender a localização de comida no final de um labirinto. Em humanos e animais, as memórias espaciais são resumidas como um mapa cognitivo.[5]
A memória espacial possui representações dentro da memória de trabalho, da memória de curto prazo e da memória de longo prazo. Pesquisas indicam que há áreas específicas do cérebro associadas à memória espacial.[6] Diversos métodos são usados para medir a memória espacial em crianças, adultos e animais.[5]
Memória de trabalho espacial
A memória de trabalho (MT) pode ser descrita como um sistema de capacidade limitada que permite armazenar e processar temporariamente informações. Esse armazenamento temporário permite realizar ou trabalhar em tarefas complexas enquanto mantém informações na mente.[7] Por exemplo, a capacidade de resolver um problema matemático complicado utiliza a memória de trabalho. Por exemplo, a capacidade de trabalhar em um problema matemático complicado utiliza a memória de trabalho.
Uma teoria influente da MT é o modelo multicomponente de memória de trabalho de Baddeley e Hitch.[7][8][9] A versão mais recente desse modelo sugere que há quatro subcomponentes na MT: o "ciclo fonológico", o "bloco de rascunho visuoespacial", o "executivo central" e o "buffer episódico".[7] Um componente desse modelo, o bloco de rascunho visuoespacial, é provavelmente responsável pelo armazenamento temporário, manutenção e manipulação de informações visuais e espaciais.[7][8] Um componente desse modelo, o bloco de rascunho visuoespacial, é provavelmente responsável pelo armazenamento temporário, manutenção e manipulação de informações visuais e espaciais.[7][9]

Em contraste com o modelo multicomponente, alguns pesquisadores acreditam que a MCP deve ser vista como uma construção unitária.[9] A esse respeito, acredita-se que as informações visuais, espaciais e verbais sejam organizadas por níveis de representação, e não pelo tipo de armazenamento ao qual pertencem.[9] Na literatura, sugere-se que mais pesquisas sobre o fracionamento da MCP e da MT sejam exploradas.[9][10] No entanto, grande parte das pesquisas sobre a construção da memória visuoespacial foi conduzida de acordo com o paradigma avançado por Baddeley e Hitch.[7][9][10][11][12]
O papel do executivo central
Pesquisas sobre a função exata do bloco de rascunho visuoespacial indicaram que tanto a memória de curto prazo quanto a memória de trabalho espacial dependem de recursos executivos e não são totalmente distintas.[7] Por exemplo, o desempenho em uma tarefa de memória de trabalho, mas não em uma tarefa de memória de curto prazo, foi afetado pela supressão articulatória, sugerindo que a deficiência na tarefa espacial foi causada pelo desempenho concorrente em uma tarefa que exigia amplo uso de recursos executivos. Resultados também indicaram que o desempenho foi prejudicado em tarefas de MCP e MT com supressão executiva. Isso ilustra como, no domínio visuoespacial, tanto a MCP quanto a MT requerem utilidade semelhante do executivo central.[7] Por exemplo, o desempenho em uma tarefa de memória de trabalho, mas não em uma tarefa de memória de curto prazo, foi afetado pela supressão articulatória, o que sugere que o prejuízo na tarefa espacial foi causado pelo desempenho simultâneo em uma tarefa que utilizou amplamente os recursos executivos. Os resultados também revelaram que o desempenho foi prejudicado em tarefas de MCP e MT com supressão executiva.[7] Isso ilustra como, no domínio visuoespacial, tanto a MCP quanto a MT exigem uma utilidade semelhante do executivo central.
Além disso, durante uma tarefa de visualização espacial (que está relacionada ao funcionamento executivo e não à MCP ou MT), a supressão executiva concorrente prejudicou o desempenho, indicando que os efeitos foram devido a demandas comuns sobre o executivo central e não ao armazenamento de curto prazo.[7] Os pesquisadores concluíram que o executivo central emprega estratégias cognitivas que permitem aos participantes codificar e manter representações mentais durante tarefas de memória de curto prazo. Os pesquisadores concluíram com a explicação de que o executivo central emprega estratégias cognitivas que permitem aos participantes codificar e manter representações mentais durante as tarefas de memória de curto prazo.[7]
Embora os estudos sugiram que o executivo central está intimamente envolvido em várias tarefas espaciais, a maneira exata como eles estão conectados ainda não foi totalmente definida.[13]
Memória espacial de curto prazo
A memória de curto prazo (MCP) pode ser descrita como um sistema que permite armazenar e gerenciar temporariamente informações necessárias para realizar tarefas cognitivas complexas.[14] Tarefas que utilizam a memória de curto prazo incluem aprendizado, raciocínio e compreensão. A memória espacial é um processo cognitivo que permite que uma pessoa se lembre de diferentes locais, bem como das relações espaciais entre objetos. Isso permite que uma pessoa se lembre de onde um objeto está em relação a outro objeto; por exemplo, permitindo que alguém navegue em uma cidade familiar. Diz-se que as memórias espaciais se formam depois que a pessoa já reuniu e processou informações sensoriais sobre seu ambiente.[14]
Memória espacial de longo prazo
A recuperação da memória espacial é estruturada em uma hierarquia. As pessoas recordam o configuração geral de um espaço específico e, em seguida, "ativam locais-alvo" dentro desse conjunto espacial.[15] Esse paradigma inclui uma escala ordinal de características que um indivíduo deve considerar para informar seu mapa cognitivo.[16] A recordação de detalhes espaciais é um procedimento de cima para baixo que exige que o indivíduo recupere primeiro as características superordenadas de um mapa cognitivo, seguidas pelas características ordenadas e subordinadas. Duas características espaciais são proeminentes na navegação de um trajeto: o layout geral e a orientação por pontos de referência. As pessoas não apenas conseguem aprender sobre o layout espacial de seus arredores, mas também podem construir novas rotas e relações espaciais por meio de inferências.
Um mapa cognitivo é "um modelo mental da configuração espacial de objetos que permite a navegação ao longo de um caminho otimizado entre pares arbitrários de pontos".[17] Esse mapa mental é construído sobre dois fundamentos principais: o layout, também conhecido como conhecimento de rota, e a orientação por pontos de referência. O layout é potencialmente o primeiro método de navegação que as pessoas aprendem a utilizar; seu funcionamento reflete as compreensões mais básicas do mundo.
Quando as crianças começam a andar, por volta dos dezoito meses, elas navegam pelo senso do layout do mundo.[18] McNamara, Hardy e Hirtle identificaram o pertencimento a uma região como um bloco fundamental de construção do mapa cognitivo de qualquer pessoa. Especificamente, o pertencimento a uma região é definida por qualquer tipo de limite, seja físico, perceptual ou subjetivo.[19] Os limites estão entre as qualidades mais básicas e endêmicas no mundo ao nosso redor. Essas fronteiras nada mais são do que linhas axiais, uma característica para a qual as pessoas têm uma predisposição ao se relacionar com o espaço; por exemplo, um determinante de linha axial é a gravidade.[20][21] As linhas axiais ajudam a dividir nossas percepções em regiões. Essa ideia de um mundo parcelado é ainda mais apoiada pela descoberta de que itens recordados juntos têm maior probabilidade de estarem agrupados dentro da mesma região do mapa cognitivo maior de uma pessoa.[16] O agrupamento mostra que as pessoas tendem a organizar informações de acordo com layouts menores dentro de um mapa cognitivo maior. O agrupamento mostra que as pessoas tendem a juntar as informações de acordo com layouts menores em um mapa cognitivo maior.
Os limites não são os únicos determinantes do layout. O agrupamento também demonstra outra propriedade importante em relação às concepções espaciais, que é o fato de a recordação espacial ser um processo hierárquico. Quando alguém recorda um ambiente ou navega por um terreno, essa pessoa implicitamente recorda o layout geral primeiro. Então, devido à "rica estrutura correlacional" do conceito, uma série de associações é ativada.[15] Eventualmente, a cascata resultante de ativações despertará os detalhes específicos que correspondem à região sendo recordada. É assim que as pessoas codificam muitas entidades de diferentes níveis ontológicos, como a localização de um grampeador; em uma mesa; que está em um escritório. Por fim, a cascata de ativações resultante despertará os detalhes específicos que correspondem à região que está sendo lembrada. É assim que as pessoas codificam muitas entidades de vários níveis ontológicos, como a localização de um grampeador, em uma mesa, que fica no escritório.
Pode-se recordar apenas uma região de cada vez (um gargalo). Um gargalo no sistema de navegação cognitiva de uma pessoa pode ser um problema. Por exemplo, se houver a necessidade de um desvio repentino em uma longa viagem de carro. A falta de experiência em um local, ou simplesmente seu tamanho, pode desorientar o layout mental, especialmente em um lugar grande e desconhecido com muitos estímulos avassaladores. Nesses ambientes, as pessoas ainda conseguem se orientar e encontrar seu caminho usando pontos de referência. Essa capacidade de "priorizar objetos e regiões em cenas complexas para seleção (e) reconhecimento" foi rotulada por Chun e Jiang em 1998.[22] Os pontos de referência fornecem orientação ao ativar "associações aprendidas entre o contexto global e os locais-alvo".[15] Mallot e Gillner mostraram que os sujeitos aprenderam uma associação entre um ponto de referência específico e a direção de uma curva, reforçando ainda mais a relação entre associações e pontos de referência.[23] Também mostraram que os participantes aprenderam uma associação entre um ponto de referência específico e a direção de uma curva, o que promoveu a relação entre associações e pontos de referência.[23] Shelton e McNamara resumiram de forma sucinta por que os pontos de referência, como marcadores, são tão úteis: "a localização... não pode ser descrita sem fazer referência à orientação do observador".[20]
As pessoas utilizam tanto o layout de um espaço específico quanto a presença de pontos de referência para navegar. Os psicólogos ainda não explicaram se o layout afeta os pontos de referência ou se os pontos de referência determinam os limites de um layout. Por causa disso, o conceito sofre de um paradoxo do tipo o ovo ou a galinha. McNamara descobriu que os sujeitos usam "grupos de pontos de referência como quadros de referência intrínsecos", o que apenas confunde ainda mais a questão.[17]
As pessoas percebem objetos em seu ambiente em relação a outros objetos nesse mesmo ambiente. Os pontos de referência e o layout são sistemas complementares para a recordação espacial, mas não se sabe como esses dois sistemas interagem quando ambos os tipos de informação estão disponíveis. Como resultado, as pessoas precisam fazer certas suposições sobre a interação entre os dois sistemas. Por exemplo, os mapas cognitivos não são "absolutos", mas sim, como qualquer um pode atestar, são "usados para fornecer um padrão...(que) é modulado de acordo com... as demandas da tarefa".[15] Os psicólogos também acreditam que os mapas cognitivos são baseados em instâncias, o que explica a "correspondência discriminativa com experiências passadas". Os psicólogos também acreditam que os mapas cognitivos são baseados em instâncias, o que explica a "correspondência discriminatória com a experiência passada".[15]
Este campo tem sido tradicionalmente prejudicado por variáveis confusas, como custo e a possibilidade de exposição prévia a um ambiente experimental. Avanços tecnológicos, incluindo aqueles em tecnologia de realidade virtual, tornaram os resultados mais acessíveis. A realidade virtual oferece aos experimentadores o luxo de um controle extremo sobre o ambiente de teste. Qualquer variável pode ser manipulada, incluindo coisas que não seriam possíveis na realidade.
Realidade virtual
Durante um estudo de 2006, os pesquisadores projetaram três cidades virtuais diferentes, cada uma com seu "layout de estrada único e um conjunto único de cinco lojas".[17] No entanto, a área total dos diferentes mapas era exatamente do mesmo tamanho, 80 unidades quadradas. Neste experimento, os participantes tiveram que participar de dois conjuntos diferentes de testes.
Um estudo conduzido na Universidade de Maryland comparou o efeito de diferentes níveis de imersão na recuperação da memória espacial.[24] No estudo, 40 participantes usaram tanto um desktop tradicional quanto um dispositivo de exibição montado na cabeça para visualizar dois ambientes, uma cidade medieval e um palácio ornamentado, onde memorizaram dois conjuntos de 21 rostos apresentados como retratos 3D. Após visualizar esses 21 rostos por 5 minutos, seguido por um breve período de descanso, os rostos nos ambientes virtuais foram substituídos por números, e os participantes recordaram qual rosto estava em cada localização. O estudo constatou que, em média, aqueles que usaram o dispositivo de exibição montado na cabeça recordaram os rostos com 8,8% mais precisão e com maior confiança. Os participantes afirmaram que aproveitar seus sentidos vestíbulo-proprioceptivos inatos com o dispositivo de exibição montado na cabeça e mapear aspectos do ambiente em relação ao seu corpo, elementos ausentes no desktop, foram fundamentais para seu sucesso.
Expertise espacial
Na literatura, há evidências de que especialistas em um determinado campo conseguem realizar tarefas de memória de acordo com suas habilidades em um nível excepcional.[12] O nível de habilidade demonstrado pelos especialistas pode exceder os limites da capacidade normal do MCP e da MT. Como os especialistas têm uma enorme quantidade de conhecimento pré-aprendido e específico da tarefa, eles podem ser capazes de codificar informações de maneira mais eficiente.[12]
Um estudo interessante que investigou a memória de motoristas de táxi para ruas em Helsinque, Finlândia, examinou a função do conhecimento espacial pré-aprendido.[12] Esse estudo comparou especialistas a um grupo de controle para determinar como esse conhecimento pré-aprendido em seu domínio de habilidades permite que eles superem as limitações de capacidade da MCP e da MT. O estudo usou quatro níveis de aleatoriedade espacial:[12]
- Ordem da rota - rota espacialmente contínua;
- Rota aleatória - lista espacialmente contínua apresentada aleatoriamente;
- Ordem do mapa - nomes das ruas formando uma linha reta no mapa, mas omitindo ruas intermediárias;
- Aleatório no mapa - ruas no mapa apresentadas em ordem aleatória.

Os resultados deste estudo indicam que a recordação dos motoristas de táxi (especialistas) de ruas foi maior nas condições de ordem de rota e ordem de mapa do que nas duas condições aleatórias. Isso indica que os especialistas foram capazes de usar seu conhecimento espacial pré-aprendido para organizar as informações de maneira a superar as limitações de capacidade da MCP e da MT.[12]
A estratégia de organização que os motoristas empregaram é conhecida como "chunking" (agrupamento) Além disso, os comentários feitos pelos especialistas durante o procedimento apontam para o uso do conhecimento de rota na realização da tarefa.[12] Para garantir que era de fato informação espacial que eles estavam codificando, os pesquisadores também apresentaram listas em ordem alfabética e categorias semânticas.[12] No entanto, os pesquisadores descobriram que era de fato informação espacial que os especialistas estavam agrupando, permitindo-lhes superar as limitações de ambas as memórias visuoespaciais, MCP e MT.[12]
Pesquisas com animais
Certas espécies de parídeos e corvídeos (como o chapim-de-cabeça-preta e aves do gênero Aphelocoma) são capazes de usar a memória espacial para lembrar onde, quando e que tipo de alimento armazenaram. Estudos com ratos e esquilos também sugeriram que eles utilizam a memória espacial para localizar alimentos previamente escondidos.[25] Experimentos usando o labirinto radial permitiram que os pesquisadores controlassem variáveis como o tipo de alimento escondido, os locais onde o alimento foi armazenado, o intervalo de retenção e quaisquer pistas olfativas que poderiam interferir nos resultados das pesquisas de memória. Estudos indicaram que ratos possuem memória de onde esconderam alimentos e que tipo de alimento foi escondido.[25] Isso é demonstrado no comportamento de recuperação, de modo que os ratos são seletivos, visitando com mais frequência os braços do labirinto onde esconderam alimentos preferidos do que os braços com alimentos menos preferidos ou onde nenhum alimento foi escondido. As evidências da memória espacial de algumas espécies de animais, como ratos, indicam então que eles utilizam a memória espacial para localizar e recuperar estoques de alimentos escondidos.[25]
Um estudo que utilizou rastreamento por GPS para observar onde gatos domésticos vão quando seus donos os deixam sair relatou que os gatos possuem uma memória espacial significativa. Alguns dos gatos no estudo demonstraram uma memória espacial de longo prazo excepcional. Um deles, que normalmente não se afastava mais de 200 a 250 metros de sua casa, inesperadamente percorreu cerca de 1250 mm de distância. Inicialmente, os pesquisadores pensaram que se tratava de um mau funcionamento do GPS, mas logo descobriram que os donos do gato haviam saído da cidade naquele fim de semana, e que a casa para onde o gato foi era a antiga residência dos donos. Os donos e o gato não moravam naquela casa há mais de um ano.[26]
Distinção visuoespacial
Logie (1995) propôs que o "bloco de rascunho visuoespacial" é dividido em dois subcomponentes, um visual e outro espacial.[11] Esses são, respectivamente, o cache visual e o escriba interno. O cache visual é um armazenamento visual temporário que inclui dimensões como cor e forma. Por outro lado, o escriba interno é um mecanismo de ensaio para informações visuais e é responsável por informações relacionadas a sequências de movimento.[11] Embora tenha sido notada na literatura uma falta geral de consenso sobre essa distinção,[10][27][28] há um número crescente de evidências de que os dois componentes são separados e desempenham funções diferentes.
A memória visual é responsável por reter formas e cores visuais (ou seja, o quê), enquanto a memória espacial é responsável por informações sobre localizações e movimentos (ou seja, onde). Essa distinção nem sempre é clara, já que parte da memória visual envolve informações espaciais e vice-versa. Por exemplo, a memória para formas de objetos geralmente envolve manter informações sobre o arranjo espacial das características que definem o objeto em questão.[27]
Na prática, os dois sistemas trabalham juntos em certa medida, mas diferentes tarefas foram desenvolvidas para destacar as habilidades únicas envolvidas na memória visual ou espacial. Por exemplo, o teste de padrões visuais (TPV) mede o alcance visual, enquanto a "tarefa de blocos de Corsi" mede o alcance espacial. Estudos correlacionais das duas medidas sugerem uma separação entre habilidades visuais e espaciais, devido à falta de correlação encontrada entre elas em pacientes saudáveis e com lesões cerebrais.[10]
O suporte para a divisão dos componentes de memória visual e espacial é encontrado por meio de experimentos usando o paradigma de dupla tarefa. Vários estudos mostraram que a retenção de formas ou cores visuais (ou seja, informações visuais) é interrompida pela apresentação de imagens irrelevantes ou ruído visual dinâmico. Por outro lado, a retenção de localização (ou seja, informações espaciais) é interrompida apenas por tarefas de rastreamento espacial, tarefas de toque espacial e movimentos oculares.[27][28] Por exemplo, os participantes completaram tanto o TPV quanto a tarefa de blocos de Corsi em um experimento de interferência seletiva. Durante o intervalo de retenção do TPV, o sujeito visualizou imagens irrelevantes (por exemplo, pinturas avant-garde). A tarefa de interferência espacial exigia que os participantes seguissem, tocando os estímulos, um arranjo de pequenos pinos de madeira que estavam escondidos atrás de uma tela. Tanto o alcance visual quanto o espacial foram reduzidos pelas respectivas tarefas de interferência, confirmando que a tarefa de blocos de Corsi está principalmente relacionada à memória de trabalho espacial.[10]
Medição
Existem várias tarefas que os psicólogos utilizam para medir a memória espacial em adultos, crianças e modelos animais. Essas tarefas permitem que profissionais identifiquem irregularidades cognitivas em adultos e crianças e possibilitam que pesquisadores administrem diferentes tipos de drogas e/ou lesões nos participantes e meçam os efeitos consequentes na memória espacial.
Tarefa de toque nos blocos de Corsi
A tarefa de toque nos blocos de Corsi, também conhecida como teste de extensão de Corsi, é um teste psicológico comumente usado para determinar a extensão da memória visuoespacial e as habilidades implícitas de aprendizado visuoespacial de um indivíduo.[29][30] Os participantes sentam-se diante de nove blocos de madeira de 3x3 cm fixados em uma base de 25x30 cm em uma ordem aleatória padrão. O experimentador toca nos blocos em uma sequência que os participantes devem replicar. Os blocos são numerados do lado do experimentador para facilitar a demonstração do padrão. O comprimento da sequência aumenta a cada tentativa até que o participante não consiga mais replicar o padrão corretamente. O teste pode ser usado para medir a memória espacial de curto e longo prazo, dependendo do tempo entre o teste e a recordação.
O teste foi criado pelo canadense neuropsicólogo Phillip Corsi, que o modelou com base na tarefa de extensão de dígitos de Hebb, substituindo os itens de teste numéricos por espaciais. Em média, a maioria dos participantes alcança uma extensão de cinco itens no teste de extensão de Corsi e sete na tarefa de extensão de dígitos.
Extensão de padrão visual
A extensão de padrão visual é semelhante à tarefa de toque nos blocos de Corsi, mas é considerada um teste mais puro de recordação visual de curto prazo.[31] Os participantes recebem uma série de padrões de matriz com metade das células coloridas e a outra metade em branco. Os padrões de matriz são organizados de maneira difícil de codificar verbalmente, forçando o participante a depender da memória visuoespacial. Começando com uma matriz pequena de 2x2, os participantes copiam o padrão de matriz da memória em uma matriz vazia. Os padrões de matriz aumentam em tamanho e complexidade a uma taxa de duas células até que a capacidade do participante de replicá-los colapse. Em média, o desempenho dos participantes tende a colapsar em dezesseis células.[31]
Tarefa de extensão de caminho
Essa tarefa é projetada para medir habilidades de memória espacial em crianças.[29] O experimentador pede ao participante que visualize uma matriz em branco com um pequeno homem. Por meio de uma série de instruções direcionais, como para frente, para trás, esquerda ou direita, o experimentador guia o pequeno homem do participante por um caminho na matriz. Ao final, o participante é solicitado a indicar em uma matriz real onde o pequeno homem que ele visualizou terminou. O comprimento do caminho varia dependendo do nível de dificuldade (1–10), e as matrizes podem variar de 2x2 células a 6x6.
Labirintos dinâmicos
Os labirintos dinâmicos são destinados a medir a habilidade espacial em crianças. Nesse teste, um experimentador apresenta ao participante um desenho de um labirinto com uma imagem de um homem no centro.[29] Enquanto o participante observa, o experimentador usa o dedo para traçar um caminho desde a abertura do labirinto até o desenho do homem. O participante deve então replicar o caminho demonstrado pelo labirinto até o desenho do homem. Os labirintos variam em complexidade à medida que a dificuldade aumenta.
Labirinto de braços radiais

Criado por Olton e Samuelson em 1976,[32] o labirinto de braços radiais foi projetado para testar as capacidades de memória espacial dos ratos. Os labirintos são tipicamente desenhados com uma plataforma central e um número variável de braços[33] que se ramificam com comida colocada nas extremidades. Os braços geralmente são protegidos uns dos outros de alguma forma, mas não a ponto de impedir que pistas externas sejam usadas como pontos de referência. Na maioria dos casos, o rato é colocado no centro do labirinto e precisa explorar cada braço individualmente para recuperar comida, enquanto lembra quais braços já perseguiu. O labirinto é configurado para que o rato seja forçado a retornar ao centro antes de perseguir outro braço. Medidas são tomadas para evitar que o rato use seus sentidos olfativos para navegar, como colocar comida extra ao longo da base do labirinto.
Tarefa de navegação na água de Morris
A tarefa de navegação na água de Morris é um teste clássico para estudar aprendizado e memória espacial em ratos[34] e foi desenvolvida pela primeira vez em 1981 por Richard G. Morris, de quem o teste leva o nome. O indivíduo é colocado em um tanque redondo de água translúcida com paredes altas demais para escalar e água profunda demais para ficar em pé. As paredes do tanque são decoradas com pistas visuais para servir como pontos de referência. O rato deve nadar pelo tanque até descobrir, por acaso, logo abaixo da superfície, a plataforma escondida na qual pode subir. Tipicamente, os ratos nadam primeiro ao redor da borda do tanque antes de se aventurarem pelo centro em um padrão sinuoso até tropeçarem na plataforma escondida. No entanto, à medida que o tempo gasto no tanque aumenta a experiência, o tempo necessário para localizar a plataforma diminui, com ratos experientes nadando diretamente para a plataforma quase imediatamente após serem colocados na água. Devido à natureza da tarefa que envolve ratos nadando, a maioria dos pesquisadores acredita que a habituação é necessária para diminuir os níveis de estresse do animal. O estresse do animal pode prejudicar os resultados dos testes cognitivos.[35]
Fisiologia
Hipocampo
O hipocampo fornece aos animais um mapa espacial de seu ambiente.[36] Ele armazena informações sobre o espaço não egocêntrico (egocêntrico significa em referência à posição do corpo no espaço) e, portanto, suporta a independência de ponto de vista na memória espacial.[37] Isso significa que ele permite a manipulação de pontos de vista a partir da memória. Também é importante para a memória espacial de longo prazo do espaço alocêntrico (referente a pistas externas no espaço).[38] A manutenção e recuperação de memórias são, portanto, relacionais ou dependentes de contexto.[39] O hipocampo utiliza a memória de referência e de trabalho e tem o importante papel de processar informações sobre localizações espaciais.[40]
O bloqueio da plasticidade nesta região resulta em problemas na navegação direcionada a objetivos e prejudica a capacidade de lembrar localizações precisas.[41] Pacientes amnésicos com danos ao hipocampo não conseguem aprender ou lembrar layouts espaciais, e pacientes submetidos à remoção do hipocampo apresentam graves prejuízos na navegação espacial.[37][42]
Macacos com lesões no hipocampo não conseguem aprender associações objeto-local, e ratos também exibem déficits espaciais ao não reagirem a mudanças espaciais.[37][43] Além disso, ratos com lesões no hipocampo apresentaram amnésia retrógada temporalmente não graduada (independente do tempo) resistente ao reconhecimento de uma tarefa de plataforma aprendida apenas quando todo o hipocampo é lesionado, mas não quando é parcialmente lesionado.[44] Déficits na memória espacial também são encontrados em tarefas de discriminação espacial.[42]

Diferenças significativas nos prejuízos espaciais são encontradas entre o hipocampo dorsal e ventral. Lesões no hipocampo ventral não afetam a memória espacial, enquanto o hipocampo dorsal é necessário para a recuperação, processamento da memória de curto prazo e transferência da memória de curto prazo para períodos de atraso mais longos.[45][46][47] A infusão de anfetamina no hipocampo dorsal também foi demonstrada para melhorar a memória de locais espaciais aprendidos anteriormente.[48] Esses achados indicam que há uma dissociação funcional entre o hipocampo dorsal e ventral.
Diferenças hemisféricas dentro do hipocampo também são observadas. Um estudo com motoristas de táxi de Londres solicitou que os motoristas recordassem rotas complexas pela cidade, bem como marcos famosos dos quais não tinham conhecimento de sua localização espacial. Isso resultou na ativação apenas do hipocampo direito durante a recordação das rotas complexas, indicando que o hipocampo direito é usado para navegação em ambientes espaciais de grande escala.[49]
O hipocampo contém dois circuitos de memória distintos. Um circuito é usado para a memória de reconhecimento de lugar baseada em recordação e inclui o sistema entorrinal-CA1,[50] enquanto o outro sistema, composto pelo circuito trissináptico do hipocampo (entorrinal-dentado-CA3-CA1), é usado para a memória de recordação de lugar[51] e a facilitação da plasticidade na sinapse entorrinal-dentada em camundongos é suficiente para melhorar a recordação de lugar.[52]
As células de lugar também são encontradas no hipocampo.
Córtex parietal posterior
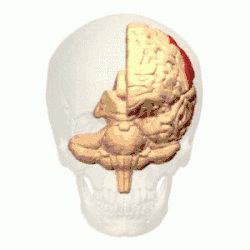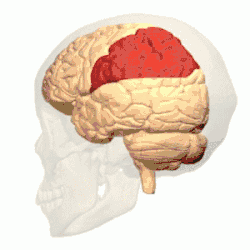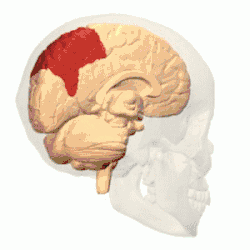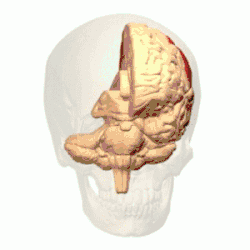
O córtex parietal codifica informações espaciais usando um quadro de referência egocêntrico. Portanto, está envolvido na transformação das coordenadas de informações sensoriais em coordenadas de ação ou efetoras, atualizando a representação espacial do corpo dentro do ambiente.[53] Como resultado, lesões no córtex parietal produzem déficits na aquisição e retenção de tarefas egocêntricas, enquanto um prejuízo menor é observado em tarefas alocêntricas.[54]
Ratos com lesões na região anterior do córtex parietal posterior reexploram objetos deslocados, enquanto ratos com lesões na região posterior do córtex parietal posterior não apresentaram reação a mudanças espaciais.[43]
Lesões no córtex parietal também são conhecidas por produzir amnésia retrógrada temporalmente não graduada.[55]
Córtex entorrinal

O córtex entorrinal dorsocaudal medial contém um mapa topograficamente organizado do ambiente espacial composto por células de Grid.[56] Esta região cerebral transforma, assim, a entrada sensorial do ambiente e a armazena como uma representação alocêntrica durável no cérebro para ser usada em integração de trajetória.[57]
O córtex entorrinal contribui para o processamento e integração de propriedades geométricas e informações no ambiente.[58] Lesões nesta região prejudicam o uso de marcos distais, mas não de marcos proximais durante a navegação e produzem um déficit dependente de atraso na memória espacial proporcional à duração do atraso.[59][60] Lesões nesta região também são conhecidas por criar déficits de retenção para tarefas aprendidas até 4 semanas, mas não 6 semanas antes das lesões.[55]
A consolidação de memória no córtex entorrinal é alcançada por meio da atividade de quinase regulada por sinal extracelular.[61]
Córtex pré-frontal
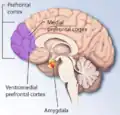
O córtex pré-frontal medial processa informações espaciais egocêntricas. Ele participa do processamento da memória espacial de curto prazo usada para guiar o comportamento de busca planejada e acredita-se que integre informações espaciais com sua significância motivacional.[47][62] A identificação de neurônios que antecipam recompensas esperadas em uma tarefa espacial apoia essa hipótese. O córtex pré-frontal medial também está implicado na organização temporal de informações.[63]
A especialização hemisférica é encontrada nesta região cerebral. O córtex pré-frontal esquerdo processa preferencialmente a memória espacial categórica, incluindo a memória de origem (referente a relações espaciais entre um lugar ou evento), enquanto o córtex pré-frontal direito processa preferencialmente a memória espacial de coordenadas, incluindo a memória de itens (referente a relações espaciais entre características de um item).[64]
Lesões no córtex pré-frontal medial prejudicam o desempenho de ratos em um labirinto de braços radiais previamente treinado, mas os ratos podem melhorar gradualmente até o nível dos controles com base na experiência.[65] Lesões nesta área também causam déficits em tarefas de não correspondência atrasada para posições e prejuízos nas tarefas de aquisição de memória espacial durante ensaios de treinamento.[66][67]
Córtex retrosplenial
O córtex retrosplenial está envolvido no processamento da memória alocêntrica e das propriedades geométricas no ambiente.[58] A inativação desta região é responsável por prejuízos na navegação no escuro e pode estar envolvida no processo de integração de trajetória.[68]
Lesões no córtex retrosplenial prejudicam consistentemente os testes de memória alocêntrica, enquanto poupam a memória egocêntrica.[69] Animais com lesões no córtex retrosplenial caudal mostram desempenho prejudicado em um labirinto de braços radiais apenas quando o labirinto é girado para remover a dependência de pistas dentro dele.[70]
Em humanos, danos ao córtex retrosplenial resultam em desorientação topográfica. A maioria dos casos envolve danos ao córtex retrosplenial direito e inclui a área de Brodmann 30. Os pacientes frequentemente apresentam dificuldades para aprender novas rotas e navegar em ambientes familiares.[71] No entanto, a maioria dos pacientes geralmente se recupera em até 8 semanas. O córtex retrosplenial processa preferencialmente informações espaciais no hemisfério direito.[71]
Córtex perirrinal
O córtex perirrinal está associado tanto à referência espacial quanto à memória de trabalho espacial.[40] Ele processa informações relacionais de pistas ambientais e localizações. Lesões no córtex perirrinal causam déficits na memória de referência e na memória de trabalho, além de aumentar a taxa de esquecimento de informações durante os ensaios de treinamento do labirinto aquático de Morris.[72] Isso explica o prejuízo na aquisição inicial da tarefa. As lesões também causam prejuízo em uma tarefa de localização de objetos e reduzem a habituação a um novo ambiente.[40]
Neuroplasticidade
As memórias espaciais são formadas após um animal coletar e processar informações sensoriais sobre seu entorno (especialmente visão e propriocepção). Em geral, mamíferos requerem um hipocampo funcional (particularmente a área CA1) para formar e processar memórias sobre espaço. Há evidências de que a memória espacial humana está fortemente ligada ao hemisfério direito do cérebro.[73][74][75]
O aprendizado espacial requer receptores NMDA e AMPA, a consolidação requer receptores NMDA, e a recuperação de memórias espaciais requer receptores AMPA.[76] Em roedores, a memória espacial mostrou-se correlacionada com o tamanho de uma parte da projeção de fibra musgosa hipocampal.[77]
A função dos receptores NMDA varia de acordo com a sub-região do hipocampo. Receptores NMDA são necessários no CA3 do hipocampo quando as informações espaciais precisam ser reorganizadas, enquanto os receptores NMDA no CA1 são necessários na aquisição e recuperação de memória após um atraso, bem como na formação de campos de lugar no CA1.[78] O bloqueio dos receptores NMDA impede a indução de potenciação de longa duração e prejudica o aprendizado espacial.[79]
O CA3 do hipocampo desempenha um papel especialmente importante na codificação e recuperação de memórias espaciais. O CA3 é inervado por dois caminhos aferentes conhecidos como o caminho perfurante (PPCA3) e as fibras musgosas (MFs) mediadas pelo giro dentado (GD). O primeiro caminho é considerado o caminho de índice de recuperação, enquanto o segundo está relacionado à codificação.[80]
Distúrbios/déficits
Desorientação topográfica
A desorientação topográfica (DT) é um distúrbio cognitivo que resulta na incapacidade do indivíduo de se orientar em ambientes reais ou virtuais. Os pacientes também enfrentam dificuldades em tarefas dependentes de informações espaciais. Esses problemas podem ser resultado de uma interrupção na capacidade de acessar o mapa cognitivo, uma representação mental do ambiente circundante, ou da incapacidade de julgar a localização de objetos em relação a si mesmo.[81]
A desorientação topográfica do desenvolvimento (DTD) é diagnosticada quando os pacientes mostram uma incapacidade de navegar mesmo em ambientes familiares desde o nascimento, sem causas neurológicas aparentes, como lesões ou danos cerebrais. A DTD é um distúrbio relativamente novo e pode ocorrer em diferentes graus de severidade.
Um estudo foi realizado para verificar se a desorientação topográfica afetava indivíduos com comprometimento cognitivo leve (CCL). O estudo recrutou 41 pacientes diagnosticados com CCL e 24 indivíduos controle saudáveis. Os padrões estabelecidos para este experimento foram:
- Queixa cognitiva subjetiva pelo paciente ou seu cuidador;
- Função cognitiva geral normal acima do percentil 16 na versão coreana do Mini-Exame do Estado Mental ("K-MMSE").
- Atividades normais de vida diária (AVD) avaliadas clinicamente e em uma escala padronizada (como descrito abaixo).
- Declínio cognitivo objetivo abaixo do percentil 16 em testes neuropsicológicos.
- Exclusão de demência.
A DT foi avaliada clinicamente em todos os participantes. Avaliações neurológicas e neuropsicológicas foram determinadas por uma varredura de imagem por ressonância magnética realizada em cada participante. A morfometria baseada em voxel foi usada para comparar padrões de atrofia de matéria cinzenta entre pacientes com e sem DT e um grupo de controles normais. O resultado do experimento foi que a DT foi encontrada em 17 dos 41 pacientes com CCL (41,4%). As habilidades funcionais foram significativamente prejudicadas em pacientes com CCL com DT em comparação com pacientes com CCL sem DT, e a presença de DT em pacientes com CCL está associada à perda de matéria cinzenta nas regiões temporais mediais, incluindo o hipocampo.[82]
Danos no hipocampo e esquizofrenia
Pesquisas com ratos indicam que a memória espacial pode ser adversamente afetada por danos neonatais ao hipocampo de uma maneira que se assemelha à esquizofrenia. A esquizofrenia é considerada decorrente de problemas de desenvolvimento neural logo após o nascimento.[83]
Ratos são comumente usados como modelos de pacientes com esquizofrenia. Os experimentadores criam lesões na área do hipocampo ventral logo após o nascimento, um procedimento conhecido como lesão hipocampal ventral neonatal (LHVN). Ratos adultos com LHVN mostram indicadores típicos de esquizofrenia, como hipersensibilidade a psicoestimulantes, interações sociais reduzidas e déficits em inibição de prepulso, memória de trabalho e mudança de foco.[84][85][86][87][88] Semelhante à esquizofrenia, ratos lesionados falham em usar o contexto ambiental em tarefas de aprendizado espacial, mostrando dificuldade em completar o labirinto de braços radiais e o labirinto aquático de Morris.[89][90][91]
NEIL1
A endonuclease VIII-1 (NEIL1) é uma enzima de reparo de DNA amplamente expressa no cérebro. NEIL1 é uma glicosilase de DNA que inicia o primeiro passo no reparo por excisão de base ao clivar bases danificadas por espécies reativas de oxigênio e, em seguida, introduzir uma quebra na fita de DNA por meio de uma reação de liase associada. Essa enzima reconhece e remove bases de DNA oxidadas, incluindo formamidopirimidina, glicol de timina, 5-hidroxiuracila e 5-hidroxicitosina. A NEIL1 promove a retenção de memória espacial de curto prazo.[92] Camundongos sem NEIL1 apresentaram retenção de memória espacial de curto prazo prejudicada em um teste de labirinto aquático.[92]
Uso de GPS e a cognição espacial

A tecnologia do GPS (sistema de posicionamento global) revolucionou a forma como navegamos e exploramos nosso ambiente. O GPS tornou-se uma ferramenta essencial em nossas vidas diárias, fornecendo informações em tempo real sobre nossa localização e as direções que precisamos tomar para chegar ao nosso destino. No entanto, alguns pesquisadores levantaram preocupações sobre o impacto do uso de GPS em nosso aprendizado e memória espacial.
O aprendizado espacial refere-se à nossa capacidade de perceber, lembrar e usar informações espaciais adquiridas no ambiente. A memória, por outro lado, envolve nossa capacidade de armazenar e recuperar informações sobre o mundo ao nosso redor. Tanto o aprendizado quanto a memória espacial são cruciais para nossa habilidade de navegar e explorar nosso ambiente de forma eficaz. O uso de GPS mostrou ter efeitos positivos e negativos no aprendizado e na memória espacial. Pesquisas indicaram que pessoas que dependem de GPS para navegação são menos propensas a desenvolver e usar mapas mentais e têm mais dificuldade em lembrar detalhes sobre o ambiente, pois o uso de GPS pode levar a um declínio dessas habilidades ao longo do tempo.[93] Além disso, os usuários de GPS tendem a confiar mais na tecnologia do que em suas próprias habilidades cognitivas, levando a uma perda de confiança em suas habilidades de navegação.[94]
No entanto, essa perda de confiança nas próprias habilidades é contrabalançada pelo conhecimento de que se perder não é mais um problema, graças ao GPS em nossos telefones, o que, por sua vez, restaura nossa confiança em nossa capacidade de orientação. Alguns resultados benéficos atribuídos à assistência do GPS são uma navegação mais eficiente e precisa, juntamente com uma redução significativa na carga cognitiva necessária para a navegação. Quando as pessoas usam dispositivos GPS, elas não precisam se preocupar em lembrar a rota, prestar atenção a marcos, ou verificar constantemente mapas. Isso pode liberar recursos cognitivos para outras tarefas, levando a um melhor desempenho nessas tarefas e níveis mais altos de concentração e foco. Isso permite liberar recursos cognitivos para facilitar o processamento de informação e o aprendizado.[95]
Para compensar os problemas decorrentes do uso de GPS, houve pesquisas substanciais que propuseram formas alternativas de navegação por GPS ou adições às existentes que demonstraram melhorar o aprendizado espacial. Um estudo de 2021 implementou um sistema de áudio espacial 3D semelhante a uma bússola auditiva, onde os usuários são direcionados ao seu destino sem instruções explícitas. Em vez de serem conduzidos passivamente por direções verbais, os usuários são incentivados a desempenhar um papel ativo em sua própria navegação espacial. Isso levou a mapas cognitivos mais precisos do espaço, uma melhoria que foi demonstrada quando os participantes do estudo desenharam mapas precisos após realizar uma tarefa de caça ao tesouro.[96] Outro estudo sugeriu destacar características locais, como marcos, ao longo da rota e em pontos de decisão; ou destacar características estruturais que fornecem orientação global (não os detalhes da rota tomada pelos participantes do estudo, mas marcos da área maior ao redor). O estudo mostrou que acentuar características locais em mapas de orientação (GPS) suporta a aquisição de conhecimento de rota, que foi medido com uma tarefa de apontamento e uma tarefa de recordação de características globais.[97]
Além disso, em pessoas com perda de visão e deficiência visual, o uso de GPS oferece vantagens no aprendizado e na memória espacial. Pessoas cegas e com deficiência visual frequentemente precisam obter informações sobre locais com antecedência e praticar uma rota específica com a ajuda de um parente, amigo ou instrutor especializado antes de viajar para o destino de forma independente. O GPS ajuda fornecendo informações úteis, permitindo que se tornem mais independentes e confiantes em suas viagens para um destino específico.
Outro artigo de pesquisa afirma que o GPS pode ser usado para pacientes que sofrem de demência. Em um estudo realizado em 2014, motoristas com a doença de Alzheimer (DA) leve a muito leve realizaram três ensaios de condução com diferentes configurações de GPS (normal, apenas visual e apenas áudio). Os participantes foram solicitados a realizar várias tarefas de condução em um simulador de condução seguindo as instruções do GPS. Este estudo descobriu que o uso de instruções auditivas simples e únicas, na ausência da saída visual do GPS, poderia potencialmente ajudar pessoas com DA leve a melhorar sua habilidade de condução e chegar ao seu destino, confirmando que o uso de GPS reduz as cargas cognitivas.[98] Como o uso de GPS ajudaria os pacientes na orientação, permitiria que permanecessem seguros em público e recuperassem seu senso de autossuficiência. No geral, as evidências são mais fortes sobre o uso de tecnologias de GPS para evitar danos e promover bem-estar.[99]
O impacto do uso de GPS no aprendizado e na memória espacial ainda não é totalmente compreendido, e mais pesquisas são necessárias para explorar os efeitos de longo prazo do uso de GPS nesses processos cognitivos. No entanto, está claro que a tecnologia de GPS tem benefícios e desvantagens, e os usuários devem estar cientes do impacto potencial de sua dependência do GPS.
Em conclusão, a tecnologia de GPS revolucionou a forma como navegamos e exploramos nosso ambiente, mas seu impacto em nosso aprendizado e na memória espacial ainda é um tema de debate. Embora o uso de GPS possa ajudar as pessoas a navegar de forma mais eficiente, confiante e auxiliar populações que, de outra forma, seriam significativamente prejudicadas; seu uso pode levar a um declínio nas habilidades cognitivas espaciais ao longo do tempo. Portanto, é essencial que os usuários equilibrem os benefícios e desvantagens do uso de GPS e estejam cientes de seu impacto potencial em suas habilidades cognitivas.
Dificuldades de aprendizado
O transtorno de aprendizado não verbal (TANV) é caracterizado por habilidades verbais normais, mas habilidades visuoespaciais prejudicadas. Áreas problemáticas para crianças com transtorno de aprendizado não verbal incluem aritmética, geometria e ciências. Prejuízos na memória espacial estão ligados ao transtorno de aprendizado não verbal e outras dificuldades de aprendizado.[100]
Problemas de aritmética verbais envolvem texto escrito contendo um conjunto de dados seguido por uma ou mais perguntas e requerem o uso das quatro operações aritméticas básicas (adição, subtração, multiplicação ou divisão).[28] Pesquisadores sugerem que a conclusão bem-sucedida de problemas verbais aritméticos envolve a memória de trabalho espacial (envolvida na construção de representações esquemáticas) que facilita a criação de relações espaciais entre objetos. Criar relações espaciais entre objetos é uma parte importante da resolução de problemas verbais, porque operações e transformações mentais são necessárias.[28]
Pesquisadores investigaram o papel da memória espacial e da memória visual na capacidade de resolver problemas verbais aritméticos. As crianças no estudo completaram a tarefa de blocos de Corsi (séries para frente e para trás) e uma tarefa de matriz espacial, bem como uma tarefa de memória visual chamada "teste de reconhecimento de casas". Crianças com baixo desempenho em resolução de problemas apresentaram prejuízo nas tarefas de blocos de Corsi e na tarefa de matriz espacial, mas tiveram desempenho normal no teste de reconhecimento de casas quando comparadas a crianças com desempenho normal. O experimento demonstrou que a baixa capacidade de resolução de problemas está especificamente relacionada ao processamento deficiente de informações espaciais.[28]
Sono
O sono foi identificado como benéfico para a memória espacial, ao aprimorar a consolidação de memória dependente do hipocampo,[101] elevando diferentes vias responsáveis pela força sináptica, controlando a transcrição de genes relacionados à plasticidade e a tradução de proteínas.[102] Áreas do hipocampo ativadas durante o aprendizado de rotas são reativadas durante o sono subsequente (especialmente durante o sono NREM). Um estudo demonstrou que a extensão da reativação durante o sono correlacionou-se com a melhoria na recuperação de rotas e, portanto, no desempenho da memória no dia seguinte.[103] O estudo estabeleceu a ideia de que o sono aprimora o processo de consolidação em nível de sistema, que consequentemente melhora o desempenho comportamental. Um período de vigília não tem efeito na estabilização de traços de memória, em comparação com um período de sono. O sono após a primeira noite pós-treinamento, ou seja, na segunda noite, não beneficia mais a consolidação da memória espacial. Portanto, dormir na primeira noite pós-treinamento, por exemplo, após aprender uma rota, é o mais importante.[101]
Além disso, foi ilustrado que o sono noturno precoce e tardio têm efeitos diferentes na memória espacial. O N3 do sono NREM, também referido como sono de ondas lentas (SOL), supostamente desempenha um papel saliente na criação de memória espacial dependente de sono em humanos. Particularmente no estudo conduzido por Plihal e Born (1999),[104] o desempenho em tarefas de rotação mental foi maior entre os participantes que tiveram intervalos de sono precoce (23h00–02h00) após aprender a tarefa, em comparação com aqueles que tiveram intervalos de sono tardio (03h00–06h00). Esses resultados sugerem que o sono precoce, rico em SOL, tem certos benefícios para a formação de memória espacial. Quando os pesquisadores examinaram se o sono precoce teria tal impacto em uma tarefa de preparação do radical de palavras (tarefa verbal), os resultados foram opostos. Isso não foi surpreendente para os pesquisadores, pois essas tarefas dependem principalmente da memória processual, e, portanto, beneficiam mais o sono de retenção tardia (dominado pelo sono REM) do que o precoce.[104]
A privação de sono e o sono também foram uma associação pesquisada. A privação de sono prejudica a melhoria do desempenho da memória devido a uma interrupção ativa da consolidação da memória espacial.[101] Como resultado, a memória espacial é aprimorada por um período de sono. Resultados semelhantes foram confirmados por outro estudo examinando o impacto da privação total de sono (PTS) na memória espacial de ratos.[105] No primeiro experimento conduzido, os ratos foram treinados no labirinto aquático de Morris por 12 ensaios em 6 horas para encontrar uma plataforma escondida (transparente e não visível na água) usando pistas espaciais no ambiente. Em cada ensaio, eles começaram de um ponto diferente e tiveram permissão para nadar por até 120 segundos para alcançar a plataforma.
Após a fase de aprendizado, foi realizado um ensaio de teste para avaliar a memória espacial (após 24 horas). Neste ensaio, a plataforma escondida foi removida do labirinto, e o tempo que os animais passaram na área-alvo (que antes era ocupada pela plataforma escondida) foi uma medida da persistência da memória espacial. Os ratos controle, que tiveram sono espontâneo, passaram significativamente mais tempo no quadrante-alvo em comparação com aqueles que tiveram privação total de sono. Em termos de aprendizado espacial, indicado pela latência para encontrar a plataforma escondida, não houve diferenças. Para ratos controle e privados de sono, o tempo necessário para encontrar a plataforma diminuiu a cada novo ensaio.[105]
No segundo experimento, os ratos foram treinados para nadar até uma plataforma visível cuja localização mudava em cada ensaio. Para cada novo ensaio, os ratos começaram do lado oposto da plataforma. Após o treinamento em um único ensaio, sua memória foi testada após 24 horas. A plataforma ainda estava no labirinto. A distância e o tempo necessários para nadar até a plataforma visível foram considerados como medidas de memória não espacial. Não foi encontrada diferença significativa entre ratos privados de sono e ratos controle. Da mesma forma, em termos de aprendizado espacial, indicado pela latência para alcançar a plataforma visível, não houve diferenças significativas. A PTS não afeta o aprendizado não espacial e a memória não espacial.[105]
Em referência aos efeitos da privação de sono em humanos, Dominique Piber (2021)[102] destacou em sua revisão da literatura as observações clínicas que mostram que pessoas com graves transtornos do sono frequentemente apresentam anormalidades na memória espacial. Como visível nos estudos, pacientes com insônia, que sofrem de um transtorno do sono caracterizado por sono interrompido e não restaurador e déficits no desempenho cognitivo durante o dia, foram documentados como tendo um desempenho negativo em uma tarefa espacial, em comparação com participantes saudáveis.[106][107][108][109]
Da mesma forma, sonhar tem um papel importante na memória espacial. Um estudo conduzido por Wamsley e Stickgold (2019)[110] provou que participantes que incorporam uma experiência de aprendizado recente em seu conteúdo de sonho noturno mostram uma melhoria de desempenho noturna aumentada. Assim, apoiando a hipótese de que sonhar reflete o processamento de memória no cérebro adormecido. Além disso, segundo os autores, uma das explicações é que sonhos relacionados ao labirinto são indicadores de que componentes relevantes do desempenho da memória da tarefa estão sendo reativados no cérebro adormecido. Adicionalmente, o estudo apoia a ideia de que relatórios de sonhos podem incluir uma tarefa de aprendizado experimental durante todos os estágios do sono, incluindo REM e NREM.[110]
A realidade virtual (RV) também foi usada para estudar a conexão entre sonhos e memória espacial. Ribeiro, Gounden e Quaglino (2021)[111] propuseram elementos espacializados em um contexto de RV e descobriram que após uma noite completa de sono em um ambiente doméstico, quando o material estudado foi incorporado ao conteúdo do sonho, o desempenho de recordação desses elementos foi melhor do que o desempenho obtido após um período de vigília comparável.[111]
Ver também
Referências
- ↑ Burgess, Neil (2021). Spatial memory (em inglês). [S.l.]: Encyclopædia Britannica. Consultado em 12 de junho de 2025
- ↑ Morgan, Michael M.; et al. (2010). «Spatial Memory in Humans». Encyclopedia of Psychopharmacology (em inglês). [S.l.: s.n.] pp. 1262–1266. ISBN 978-3-540-68698-9. doi:10.1007/978-3-540-68706-1_355
- ↑ Pritchard, David J. (2019). «Spatial Memory». Encyclopedia of Animal Behavior (em inglês). [S.l.: s.n.] pp. 320–326. ISBN 978-0-12-813252-4. doi:10.1016/B978-0-12-809633-8.90082-7
- ↑ Kolarik, B.; Ekstrom, A. (2015). «The Neural Underpinnings of Spatial Memory and Navigation». Brain Mapping (em inglês). [S.l.: s.n.] pp. 507–514. ISBN 978-0-12-397316-0. doi:10.1016/B978-0-12-397025-1.00277-3
- ↑ a b Wang, Jin-Hu (2019). Associative Memory Cells: Basic Units of Memory Trace (em inglês). [S.l.]: Springer Nature. p. 94. ISBN 978-9811395017. Consultado em 12 de junho de 2025
- ↑ Poeppel, David; Mangun, George R.; Gazzaniga, Michael S. (2020). The Cognitive Neurosciences (em inglês). [S.l.]: The MIT Press. p. 194. ISBN 978-0262043250. Consultado em 12 de junho de 2025
- ↑ a b c d e f g h i j k Ang, S. Y.; Lee, K. (2008). «Central executive involvement in children's spatial memory». Memory (em inglês). 16 (8): 918–933. PMID 18802804. doi:10.1080/09658210802365347
- ↑ a b Jones, D.; Farrand, P.; Stuart, G.; Morris, N.; et al. (1995). «Functional equivalence of verbal and spatial information in serial short-term memory». Journal of Experimental Psychology: Learning, Memory, and Cognition (em inglês). 21 (4): 1008–1018. PMID 7673864. doi:10.1037/0278-7393.21.4.1008
- ↑ a b c d e f Jones, D.; Farrand, P.; Stuart, G.; Morris, N.; et al. (1995). «Functional equivalence of verbal and spatial information in serial short-term memory». Journal of Experimental Psychology: Learning, Memory, and Cognition (em inglês). 21 (4): 1008–1018. PMID 7673864. doi:10.1037/0278-7393.21.4.1008
- ↑ a b c d e Della Sala, S.; Gray, C.; Baddeley, A.; Allamano, N.; Wilson, L.; et al. (1999). «Pattern span: a tool for unwelding visuo-spatial memory». Neuropsychologia (em inglês). 37 (10): 1189–1199. PMID 10509840. doi:10.1016/S0028-3932(98)00159-6
- ↑ a b c Mammarella, I. C.; Pazzaglia, F.; Cornoldi, C.; et al. (2008). «Evidence for different components in children's visuospatial working memory». British Journal of Developmental Psychology (em inglês). 26 (3): 337–355. doi:10.1348/026151007X236061. hdl:11577/2440989

- ↑ a b c d e f g h i Kalakoski, V.; Saariluoma, P. (2001). «Taxi drivers' exceptional memory of street names». Memory and Cognition (em inglês). 29 (4): 634–638. PMID 11504011. doi:10.3758/BF03200464
- ↑ Fisk, J. E.; Sharp, C. A.; et al. (2003). «The role of the executive system in visuo-spatial memory functioning». Brain and Cognition (em inglês). 52 (3): 364–381. PMID 12907181. doi:10.1016/S0278-2626(03)00183-0
- ↑ a b Johnson, E.; Adamo-Villani, N. (2010). «A Study of the Effects of Immersion on Short-term Spatial Memory». Engineering and Technology (em inglês). 71: 582–587
- ↑ a b c d e Chun, M.; Jiang, Y. (1998). «Contextual Cueing: implicit learning and memory of visual context guides spatial attention». Cognitive Psychology (em inglês). 36 (1): 28–71. PMID 9679076. doi:10.1006/cogp.1998.0681
- ↑ a b McNamara, T.; Hardy, J.; Hirtle, S.; et al. (1989). «Subjective hierarchies in spatial memory». Journal of Experimental Psychology (em inglês). 15 (2): 211–227. PMID 2522511. doi:10.1037/0278-7393.15.2.211
- ↑ a b c Newman, E.L.; Caplan, J.B.; Kirschen, M.P.; Korolev, I.O.; Sekuler, R.; Kahana, M.J.; et al. (2007). «Learning Your Way Around Town: How Virtual Taxicab Drivers Learn to Use Both Layout and Landmark Information». Cognition (em inglês). 104 (2): 231–253. PMID 16879816. doi:10.1016/j.cognition.2006.05.013
- ↑ Hermer, Linda; Spelke, Elizabeth S. (julho de 1994). «A geometric process for spatial reorientation in young children». Nature (em inglês) (6484): 57–59. ISSN 1476-4687. doi:10.1038/370057a0. Consultado em 29 de junho de 2025
- ↑ McNamara, Timothy P.; Hardy, James K.; Hirtle, Stephen C. (1989). «Subjective hierarchies in spatial memory.». Journal of Experimental Psychology: Learning, Memory, and Cognition (em inglês) (2): 211–227. ISSN 1939-1285. doi:10.1037/0278-7393.15.2.211. Consultado em 29 de junho de 2025
- ↑ a b Shelton, Amy L.; McNamara, Timothy P. (1 de dezembro de 2001). «Systems of Spatial Reference in Human Memory». Cognitive Psychology (4): 274–310. ISSN 0010-0285. doi:10.1006/cogp.2001.0758. Consultado em 29 de junho de 2025
- ↑ Kim, Young Ook; Penn, Alan (julho de 2004). «Linking the Spatial Syntax of Cognitive Maps to the Spatial Syntax of the Environment». Environment and Behavior (em inglês) (4): 483–504. ISSN 0013-9165. doi:10.1177/0013916503261384. Consultado em 29 de junho de 2025
- ↑ Chun, Marvin M.; Jiang, Yuhong (1 de junho de 1998). «Contextual Cueing: Implicit Learning and Memory of Visual Context Guides Spatial Attention». Cognitive Psychology (1): 28–71. ISSN 0010-0285. doi:10.1006/cogp.1998.0681. Consultado em 29 de junho de 2025
- ↑ a b Gillner S, Mallot H (2000). «The role of global and local landmarks in virtual environment navigation». Perception (em inglês). 9 (1): 69–83. doi:10.1162/105474600566628. hdl:11858/00-001M-0000-0013-E7E9-0
- ↑ Krokos, Eric; Plaisant, Catherine; Varshney, Amitabh (2018). «Virtual Memory Palaces: Immersion Aids Recall». Virtual Reality (em inglês). 23: 1–15. doi:10.1007/s10055-018-0346-3
- ↑ a b c Bird, L. R.; Roberts, W. A.; Abroms, B.; Kit, K. A.; Crupi, C. (2003). «Spatial memory for food hidden by rats (Rattus norvegicus) on the radial maze: studies of memory for where, what, and when». Journal of Comparative Psychology (em inglês). 117 (2): 176–187. PMID 12856788. doi:10.1037/0735-7036.117.2.176
- ↑ Jennifer S. Holland (2014). «Watch: How Far Do Your Cats Roam?». National Geographic (em inglês). Consultado em 12 de junho de 2025. Arquivado do original em 2014
- ↑ a b c Klauer, K.C.; Zhao, Z.; et al. (2004). «Double dissociations in visual and spatial short-term memory». Journal of Experimental Psychology: General (em inglês). 133 (3): 355–381. PMID 15355144. doi:10.1037/0096-3445.133.3.355
- ↑ a b c d e Passolunghi, M.C.; Mammarella, I.C.; et al. (2010). «Spatial and visual working memory ability in children with difficulties in arithmetic word problem solving». European Journal of Cognitive Psychology (em inglês). 22 (6): 944–963. doi:10.1080/09541440903091127
- ↑ a b c Mammarella, I.C.; Pazzaglia, F.; Cornoldi, C. (2008). «Evidence of different components in children's visuospatial working memory». British Journal of Developmental Psychology (em inglês). 26 (3): 337–355. doi:10.1348/026151007X236061. hdl:11577/2440989. Consultado em 12 de junho de 2025
- ↑ Corsi, P. M. (1972). «Human memory and the medial temporal region of the brain». Dissertation Abstracts International (em inglês). 34 (2): 891
- ↑ a b Della Sala, S.; Gray, C.; Baddeley, A.; Wilson, L. (1997). The Visual Patterns Test: A new test of short-term visual recall (em inglês). Feltham, Suffolk: Thames Valley Test Company
- ↑ Olton, D.S.; Samuelson, R.J. (1976). «Remembrance of places past: spatial memory in rats». Journal of Experimental Psychology: Animal Behavior Processes (em inglês). 2 (2): 97–116. doi:10.1037/0097-7403.2.2.97
- ↑ Cole, M.R.; Chappell-Stephenson, Robyn (2003). «Exploring the limits of spatial memory using very large mazes». Learning & Behavior (em inglês). 31 (4): 349–368. PMID 14733483. doi:10.3758/BF03195996
- ↑ Morris, R. G. (1981). «Spatial Localization Does Not Require the Presence of Local Cues». Learning and Motivation (em inglês). 12 (2): 239–260. doi:10.1016/0023-9690(81)90020-5
- ↑ Sharma, Sunita; Rakoczy, Sharlene; Brown-Borg, Holly (2010). «Assessment of spatial memory in mice». Life Sciences (em inglês). 87 (17–18): 521–536. PMC 6457258. PMID 20837032. doi:10.1016/j.lfs.2010.09.004
- ↑ O'Keefe, J.; Dostrovsky, J. (1971). «The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat». Brain Research (em inglês). 34 (1): 171–175. PMID 5124915. doi:10.1016/0006-8993(71)90358-1
- ↑ a b c Squire, L. R. (1992). «Memory and the hippocampus: A synthesis from findings with rats, monkeys, and humans». Psychological Review (em inglês). 99 (2): 195–231. PMID 1594723. doi:10.1037/0033-295X.99.2.195
- ↑ Ramos, J. M. J. (2000). «Long-term spatial memory in rats with hippocampal lesions». European Journal of Neuroscience (em inglês). 12 (9): 3375–3384. PMID 10998120. doi:10.1046/j.1460-9568.2000.00206.x
- ↑ Winocur, G.; Moscovitch, M.; Caruana, D. A.; Binns, M. A. (2005). «Retrograde amnesia in rats with lesions to the hippocampus on a test of spatial memory». Neuropsychologia (em inglês). 43 (11): 1580–1590. PMID 16009240. doi:10.1016/j.neuropsychologia.2005.01.013
- ↑ a b c Liu, P.; Bilkey, D. K. (2001). «The effect of excitotoxic lesions centered on the hippocampus or perirhinal cortex in object recognition and spatial memory tasks». Behavioral Neuroscience (em inglês). 115 (1): 94–111. PMID 11256456. doi:10.1037/0735-7044.115.1.94
- ↑ Hebert, A. E.; Dash, P. K. (2004). «Nonredundant roles for hippocampal and entorhinal cortical plasticity in spatial memory storage». Pharmacology Biochemistry and Behavior (em inglês). 79 (1): 143–153. PMID 15388294. doi:10.1016/j.pbb.2004.06.016
- ↑ a b Morris RG, Garrud P, Rawlins JN, O'Keefe J (1982). «Place navigation impaired in rats with hippocampal lesions». Nature (em inglês). 297 (5868): 681–683. Bibcode:1982Natur.297..681M. PMID 7088155. doi:10.1038/297681a0
- ↑ a b Save, E.; Poucet, B.; Foreman, N.; Buhot, M. (1992). «Object exploration and reactions to spatial and nonspatial changes in hooded rats following damage to parietal cortex or hippocampal formation». Behavioral Neuroscience (em inglês). 106 (3): 447–456. PMID 1616611. doi:10.1037/0735-7044.106.3.447
- ↑ Martin, S. J.; de Hozl, L.; Morris, R. G. M. (2005). «Retrograde amnesia: neither partial nor complete hippocampal lesions in rats result in preferential sparing of remote spatial memory, even after reminding». Neuropsychologia (em inglês). 43 (4): 609–624. PMID 15716151. doi:10.1016/j.neuropsychologia.2004.07.007
- ↑ Bannerman, D. M.; Deacon, R. M. J.; Offen, S.; Friswell, J.; Grubb, M.; Rawlins, J. N. P. (2002). «Double dissociation of function within the hippocampus: Spatial memory and hyponeophagia». Behavioral Neuroscience (em inglês). 116 (5): 884–901. PMID 12369808. doi:10.1037/0735-7044.116.5.884
- ↑ Moser, M.; Moser, E. I. (1998). «Distributed encoding and retrieval of spatial memory in the hippocampus». The Journal of Neuroscience (em inglês). 18 (18): 7535–7542. PMC 6793256. PMID 9736671. doi:10.1523/JNEUROSCI.18-18-07535.1998
- ↑ a b Lee, I.; Kesner, R. P. (2003). «Time-dependent relationship between the dorsal hippocampus and the prefrontal cortex in spatial memory». The Journal of Neuroscience (em inglês). 23 (4): 1517–1523. PMC 6742248. PMID 12598640. doi:10.1523/JNEUROSCI.23-04-01517.2003
- ↑ McGaugh, J. L. (2000). «Memory—a century of consolidation». Science (em inglês). 287 (5451): 248–251. Bibcode:2000Sci...287..248M. PMID 10634773. doi:10.1126/science.287.5451.248
- ↑ Maguire, E. A.; Frackowiak, R. S. J.; Frith, C. D. (1997). «Recalling routes around London: Activation of the right hippocampus in taxi drivers». The Journal of Neuroscience (em inglês). 17 (18): 7103–7110. PMC 6573257. PMID 9278544. doi:10.1523/JNEUROSCI.17-18-07103.1997
- ↑ Brun, V. H.; Otnaess, M. K.; Molden, S.; Steffenach, H.; Witter, M. P.; Moser, M.; Moser, E. I. (2002). «Place cells and place recognition maintained by direct entorhinal-hippocampal circuitry». Science (em inglês). 296 (5576): 2243–2246. Bibcode:2002Sci...296.2243B. PMID 12077421. doi:10.1126/science.1071089
- ↑ Goodrich-Hunsaker NJ, Hunsaker MR, Kesner RP (2008). «The interactions and dissociations of the dorsal hippocampus subregions: how the dentate gyrus, CA3, and CA1 process spatial information.». Behav. Neurosci. (em inglês). 122 (1): 16–26. PMID 18298245. doi:10.1037/0735-7044.122.1.16
- ↑ Saab BJ, Georgiou J, Nath A, Lee FJ, Wang M, Michalon A, Liu F, Mansuy IM, Roder JC (2009). «NCS-1 in the dentate gyrus promotes exploration, synaptic plasticity, and rapid acquisition of spatial memory». Neuron (em inglês). 63 (5): 643–56. PMID 19755107. doi:10.1016/j.neuron.2009.08.014
- ↑ Colby, C. L.; Goldberg, M. E. (1999). «Space and attention in parietal cortex». Annual Review of Neuroscience (em inglês). 22: 319–349. PMID 10202542. doi:10.1146/annurev.neuro.22.1.319
- ↑ Save, E.; Moghaddam, M. (1996). «Effects of lesions of the associative parietal cortex on the acquisition and use of spatial memory in egocentric and allocentric navigation tasks in the rat». Behavioral Neuroscience (em inglês). 110 (1): 74–85. PMID 8652075. doi:10.1037/0735-7044.110.1.74
- ↑ a b Cho, Y. H.; Kesner, R. P. (1996). «Involvement of entorhinal cortex or parietal cortex in long-term spatial discrimination memory in rats: Retrograde amnesia». Behavioral Neuroscience (em inglês). 110 (3): 436–442. PMID 8888988. doi:10.1037/0735-7044.110.3.436
- ↑ Hafting, T.; Fyhn, M.; Molden, S.; Moser, M.; Moser, E. I. (2005). «Microstructure of a spatial map in the entorhinal cortex». Nature (em inglês). 436 (7052): 801–806. Bibcode:2005Natur.436..801H. PMID 15965463. doi:10.1038/nature03721
- ↑ Fyhn, M.; Molden, S.; Witter, M. P.; Moser, E. I.; Moser, M. (2004). «Spatial representation in the entorhinal cortex». Science (em inglês). 305 (5688): 1258–1264. Bibcode:2004Sci...305.1258F. PMID 15333832. doi:10.1126/science.1099901
- ↑ a b Parron, C.; Save, E. (2004). «Comparison of the effects of entorhinal and retrosplenial cortical lesions on habituation, reaction to spatial and non-spatial changes during object exploration in the rat». Neurobiology of Learning and Memory (em inglês). 82 (1): 1–11. PMID 15183166. doi:10.1016/j.nlm.2004.03.004
- ↑ Parron, C.; Poucet, B.; Save, E. (2004). «Entorhinal cortex lesions impair the use of distal but not proximal landmarks during place navigation in the rat». Behavioural Brain Research (em inglês). 154 (2): 345–352. PMID 15313022. doi:10.1016/j.bbr.2004.03.006
- ↑ Nagahara, H. A.; Otto, T.; Gallagher, M. (1995). «Entorhinal-perirhinal lesions impair performance of rats on two versions of place learning in the Morris water maze». Behavioral Neuroscience (em inglês). 109 (1): 3–9. PMID 7734077. doi:10.1037/0735-7044.109.1.3
- ↑ Hebert, A. E.; Dash, P. K. (2002). «Extracellular signal-regulated kinase activity in the entorhinal cortex is necessary for long-term spatial memory». Learning Memory] (em inglês). 9 (4): 156–166. PMC 182586. PMID 12177229. doi:10.1101/lm.48502
- ↑ Pratt, Wayne E; Mizumori, Sheri J.Y (2001). «Neurons in rat medial prefrontal cortex show anticipatory rate changes to predictable differential rewards in a spatial memory task». Behavioural Brain Research (em inglês) (2): 165–183. doi:10.1016/S0166-4328(01)00204-2. Consultado em 12 de junho de 2025
- ↑ Kesner, R. P.; Holbrook, T. (1987). «Dissociation of item and order spatial memory in rats following medial prefrontal cortex lesions». Neuropsychologia (em inglês). 25 (4): 653–664. PMID 3658148. doi:10.1016/0028-3932(87)90056-X
- ↑ Slotnick, S. D.; Moo, L. R. (2006). «Prefrontal cortex hemispheric specialization for categorical and coordinate visual spatial memory». Neuropsychologia (em inglês). 44 (9): 1560–1568. PMID 16516248. doi:10.1016/j.neuropsychologia.2006.01.018
- ↑ Becker, J. T.; Walker, J. A.; Olton, D. S. (1980). «Neuroanatomical bases of spatial memory». Brain Research (em inglês). 200 (2): 307–320. PMID 7417818. doi:10.1016/0006-8993(80)90922-1
- ↑ Aggleton, J. P.; Neave, N.; Nagle, S.; Sahgal, A. (1995). «A comparison of the effects of medial prefrontal, cingulate cortex, and cingulum bundle lesions on tests of spatial memory: Evidence of a double dissociation between frontal and cingulum bundle contributions». The Journal of Neuroscience (em inglês). 15 (11): 7270–7281. PMC 6578066. PMID 7472481. doi:10.1523/JNEUROSCI.15-11-07270.1995
- ↑ Lacroix, L.; White, I.; Feldon, J. (2002). «Effect of excitotoxic lesions of rat medial prefrontal cortex on spatial memory». Behavioural Brain Research (em inglês). 133 (1): 69–81. PMID 12048175. doi:10.1016/S0166-4328(01)00442-9
- ↑ Cooper, B. G.; Manka, T. F.; Mizumori, S. J. Y. (2001). «Finding your way in the dark: The retrosplenial cortex contributes to spatial memory and navigation without visual cues». Behavioral Neuroscience (em inglês). 115 (5): 1012–1028. PMID 11584914. doi:10.1037/0735-7044.115.5.1012
- ↑ Vann, S. D.; Aggleton, J. P. (2002). «Extensive cytotoxic lesions of the rat retrosplenial cortex reveal consistent deficits on tasks that tax allocentric spatial memory». Behavioral Neuroscience (em inglês). 116 (1): 85–94. PMID 11895186. doi:10.1037/0735-7044.116.1.85
- ↑ Vann, S. D.; Wilton, L. A.; Muir, J. L.; Aggleton, J. P. (2003). «Testing the importance of the caudal retrosplenial cortex for spatial memory in rats». Behavioural Brain Research (em inglês). 140 (1–2): 107–118. PMID 12644284. doi:10.1016/S0166-4328(02)00274-7
- ↑ a b Maguire, E. A. (2001). «The retrosplenial contribution to human navigation: A review of lesion and neuroimaging findings». Scandinavian Journal of Psychology (em inglês). 42 (3): 225–238. PMID 11501737. doi:10.1111/1467-9450.00233
- ↑ Liu, P.; Bilkey, D. K. (1998). «Perirhinal cortex contributions to performance in the Morris water maze». Behavioral Neuroscience (em inglês). 112 (2): 304–315. PMID 9588480. doi:10.1037/0735-7044.112.2.304
- ↑ Gutbrod, K; Cohen, R; Maier, T; Meier, E (1987). «Memory for spatial and temporal order in aphasics and right hemisphere damaged patients». Cortex (em inglês). 23 (3): 463–74. PMID 3677733. doi:10.1016/s0010-9452(87)80007-2
- ↑ Nunn, JA; Graydon, FJ; Polkey, CE; Morris, RG (1999). «Differential spatial memory impairment after right temporal lobectomy demonstrated using temporal titration». Brain (em inglês). 122 (1): 47–59. PMID 10050894. doi:10.1093/brain/122.1.47
- ↑ Tucker, DM; Hartry-Speiser, A; McDougal, L; Luu, P; Degrandpre, D (1999). «Mood and spatial memory: emotion and right hemisphere contribution to spatial cognition». Biol Psychol (em inglês). 50 (2): 103–25. PMID 10403200. doi:10.1016/S0301-0511(99)00005-8
- ↑ Liang, KC; Hon, W; Tyan, YM; Liao, WL (1994). «Involvement of hippocampal NMDA and AMPA receptors in acquisition, formation and retrieval of spatial memory in the Morris water maze». Chin J Physiol (em inglês). 37 (4): 201–12. PMID 7796636
- ↑ Crusio, W. E.; Schwegler, H. (2005). «Learning spatial orientation tasks in the radial-maze and structural variation in the hippocampus in inbred mice». Behavioral and Brain Functions (em inglês). 1 (1): 3. PMC 1143776. PMID 15916698. doi:10.1186/1744-9081-1-3
- ↑ Lee, I.; Kesner, R. P. (2002). «Differential contribution of NMDA receptors in hippocampal subregions to spatial working memory». Nature Neuroscience (em inglês). 5 (2): 162–168. PMID 11780144. doi:10.1038/nn790
- ↑ Morris, R. G. M.; Anderson, E.; Lynch, G. S.; Baudry, M. (1986). «Selective impairment of learning and blockade of long-term potentiation by an N-methyl-D-aspartate receptor anatagonist, AP5». Nature (em inglês). 319 (6056): 774–776. Bibcode:1986Natur.319..774M. PMID 2869411. doi:10.1038/319774a0
- ↑ Lee, I.; Kesner, R. P. (2004). «Encoding versus retrieval of spatial memory: Double dissociation between the dentate gyrus and the perforant path inputs into CA3 in the dorsal hippocampus». Hippocampus (em inglês). 14 (1): 66–76. PMID 15058484. doi:10.1002/hipo.10167
- ↑ Stark, M; Coslett, HB; Saffran, EM (1996). Impairment of an egocentric map of locations: implications for perception and action. 13. Cogn Neuropsychol. pp. 481–523.
- ↑ Lim, Tae-Sung; Iaria, Giuseppe; Moon, So Young (2010). «Topographical Disorientation in Mild Cognitive Impairment: A Voxel-Based Morphometry Study». Journal of Clinical Neurology (em inglês). 6 (4): 204–211. PMC 3024525. PMID 21264201. doi:10.3988/jcn.2010.6.4.204
- ↑ Lewis, D.A.; Levitt, P. (2002). «Schizophrenia as a disorder of neurodevelopment». Annual Review of Neuroscience (em inglês). 25: 409–432. PMID 12052915. doi:10.1146/annurev.neuro.25.112701.142754
- ↑ Lipska, B.K.; Weinberger, D.R. (2000). «To model a psychiatric disorder in animals: Schizophrenia as a reality test». Neuropsychopharmacology (em inglês). 23 (3): 223–239. PMID 10942847. doi:10.1016/S0893-133X(00)00137-8
- ↑ Lipska, B.K.; Aultman, J.M.; Verma, A.; Weinberger, D.R.; Moghaddam, B. (2002). «Neonatal damage of the ventral hippocampus impairs working memory in the rat». Neuropsychopharmacology (em inglês). 27 (1): 47–54. PMID 12062906. doi:10.1016/S0893-133X(02)00282-8
- ↑ Marquis, J. P.; Goulet, S.; Dore, F. Y. (2008). «Dissociable onset of cognitive and motivational dysfunctions following neonatal lesions of the ventral hippocampus in rats». Behavioral Neuroscience (em inglês). 122 (3): 629–642. PMID 18513133. doi:10.1037/0735-7044.122.3.629
- ↑ Brady, A. M. (2009). «Neonatal ventral hippocampal lesions disrupt set-shifting ability in adult rats». Behavioural Brain Research (em inglês). 205 (1): 294–298. PMID 19646488. doi:10.1016/j.bbr.2009.07.025
- ↑ Marquis, J.P.; Goulet, S.; Dore, F.Y. (2008). «Neonatal ventral hippocampus lesions disrupt extra-dimensional shift and alter dendritic spine density in the medial prefrontal cortex of juvenile rats». Neurobiology of Learning and Memory (em inglês). 90 (2): 339–346. PMID 18490183. doi:10.1016/j.nlm.2008.04.005
- ↑ Winocur, G. & Mills, J. A. (1970). Transfer between related and unrelated problems following hippocampal lesions in rats. Journal of Comparative and Physiological Psychology
- ↑ Levin, E.D.; Christopher, N.C. (2006). «Effects of clozapine on memory function in the rat neonatal hippocampal lesion model of schizophrenia». Progress in Neuro-Psychopharmacology and Biological Psychiatry (em inglês). 30 (2): 223–229. PMID 16356617. doi:10.1016/j.pnpbp.2005.10.018
- ↑ Silva-Gomez, A.B.; Bermudez, M.; Quirion, R.; Srivastava, L.K.; Picazo, O.; Flores, G. (2003). «Comparative behavioral changes between male and female postpubertal rats following neonatal excitotoxic lesions of the ventral hippocampus». Brain Research (em inglês). 973 (2): 285–292. PMID 12738072. doi:10.1016/S0006-8993(03)02537-X
- ↑ a b Canugovi C, Yoon JS, Feldman NH, Croteau DL, Mattson MP, Bohr VA (2012). «Endonuclease VIII-like 1 (NEIL1) promotes short-term spatial memory retention and protects from ischemic stroke-induced brain dysfunction and death in mice». Proc. Natl. Acad. Sci. U.S.A. (em inglês). 109 (37): 14948–53. Bibcode:2012PNAS..10914948C. PMC 3443144. PMID 22927410. doi:10.1073/pnas.1204156109
- ↑ Ruginski, Ian T.; Creem-Regehr, Sarah H.; Stefanucci, Jeanine K.; Cashdan, Elizabeth (2019). «GPS use negatively affects environmental learning through spatial transformation abilities». Journal of Environmental Psychology (em inglês). 64: 12–20. doi:10.1016/j.jenvp.2019.05.001. Consultado em 12 de junho de 2025
- ↑ Montello, D. R; Sasinka, J. M (2006). «Human factors of navigation». In: W Karwowski, W. International Encyclopedia of Ergonomics and Human Factors (em inglês). [S.l.]: CRC Press/Taylor & Francis, Ltd. pp. 2003–2008. Consultado em 12 de junho de 2025
- ↑ Brügger, A; Richter, KF; Fabrikant, SI (2019). «How does navigation system behavior influence human behavior?». Cogn Res Princ Implic (em inglês). 4 (1). 5 páginas. PMC 6374493. PMID 30758681. doi:10.1186/s41235-019-0156-5
- ↑ Clemenson, Gregory D.; Maselli, Antonella; Fiannaca, Alexander J.; Miller, Amos; Gonzalez-Franco, Mar (2021). «Rethinking GPS navigation: creating cognitive maps through auditory clues». Scientific Reports (em inglês). 11 (1): 7764. Bibcode:2021NatSR..11.7764C. PMC 8032695. PMID 33833290. doi:10.1038/s41598-021-87148-4
- ↑ Löwen, Heinrich; Krukar, Jakub; Schwering, Angela (2019). «Spatial Learning with Orientation Maps: The Influence of Different Environmental Features on Spatial Knowledge Acquisition». ISPRS International Journal of Geo-Information (em inglês). 8 (3). 149 páginas. Bibcode:2019IJGI....8..149L. doi:10.3390/ijgi8030149
- ↑ Yi, Jewel; Lee, Hoe Chung-Yeung; Parsons, Richard; Falkmer, Torbjorn (2014). «The Effect of the Global Positioning System on the Driving Performance of People with Mild Alzheimer's disease». Gerontology (em inglês). 61 (1): 79–88. PMID 25342271. doi:10.1159/000365922
- ↑ Bartlett, Ruth; Brannelly, Tula; Topo, Päivi (2019). «Using GPS Technologies with People with Dementia». Tidsskrift for Omsorgsforskning (em inglês). 5 (3): 84–98. doi:10.18261/issn.2387-5984-2019-03-08
- ↑ Mammarella, I.C.; Lucangeli, D.; Cornoldi, C. (2010). «Spatial working memory and arithmetic deficites in children with nonverbal learning difficulties». Journal of Learning Disabilities (em inglês). 43 (5): 455–468. PMID 20375290. doi:10.1177/0022219409355482. hdl:11577/2453526
- ↑ a b c Ferrara, Michele; Iaria, Giuseppe; Tempesta, Daniela; Curcio, Giuseppe; Moroni, Fabio; Marzano, Cristina; De Gennaro, Luigi; Pacitti, Claudio (agosto de 2008). «Sleep to find your way: The role of sleep in the consolidation of memory for navigation in humans». Hippocampus (em inglês) (8): 844–851. ISSN 1050-9631. doi:10.1002/hipo.20444. Consultado em 12 de junho de 2025
- ↑ a b Piber, Dominique (2021). «The role of sleep disturbance and inflammation for spatial memory». Brain, Behavior, & Immunity - Health (em inglês). 17. 100333 páginas. PMC 8474561. PMID 34589818. doi:10.1016/j.bbih.2021.100333
- ↑ Peigneux, Philippe; Laureys, Steven; Fuchs, Sonia; Collette, Fabienne; Perrin, Fabien; Reggers, Jean; Phillips, Christophe; Degueldre, Christian; Del Fiore, Guy; Aerts, Joël; Luxen, André; Maquet, Pierre (2004). «Are Spatial Memories Strengthened in the Human Hippocampus during Slow Wave Sleep?». Neuron (em inglês). 44 (3): 535–545. PMID 15504332. doi:10.1016/j.neuron.2004.10.007
- ↑ a b Plihal, Werner; Born, Jan (1999). «Effects of early and late nocturnal sleep on priming and spatial memory». Psychophysiology (em inglês). 36 (5): 571–582. PMID 10442025. doi:10.1111/1469-8986.3650571
- ↑ a b c Guan, Zhiwei; Peng, Xuwen; Fang, Jidong (2004). «Sleep deprivation impairs spatial memory and decreases extracellular signal-regulated kinase phosphorylation in the hippocampus». Brain Research (em inglês). 1018 (1): 38–47. PMID 15262203. doi:10.1016/j.brainres.2004.05.032
- ↑ Li, Yongli; Liu, Liya; Wang, Enfeng; Zhang, Hongju; Dou, Shewei; Tong, Li; Cheng, Jingliang; Chen, Chuanliang; Shi, Dapeng (2016). «Abnormal Neural Network of Primary Insomnia: Evidence from Spatial Working Memory Task fMRI». European Neurology (em inglês). 75 (1–2): 48–57. PMID 26784899. doi:10.1159/000443372
- ↑ Chen, Gui-hai; Xia, Lan; Wang, Fang; Li, Xue-Wei; Jiao, Chuan-an (2016). «Patients with chronic insomnia have selective impairments in memory that are modulated by cortisol: Memory impairment in chronic insomniacs». Psychophysiology (em inglês). 53 (10): 1567–1576. PMID 27412857. doi:10.1111/psyp.12700
- ↑ Khassawneh, Basheer Y.; Bathgate, Christina J.; Tsai, Sheila C.; Edinger, Jack D. (2018). «Neurocognitive performance in insomnia disorder: The impact of hyperarousal and short sleep duration». Journal of Sleep Research (em inglês). 27 (6): e12747. PMID 30069961. doi:10.1111/jsr.12747
- ↑ He, Shuo; Chen, Xi-Xi; Ge, Wei; Yang, Shuai; Chen, Jun-Tao; Niu, Jing-Wen; Xia, Lan; Chen, Gui-Hai (2021). «Are Anti-Inflammatory Cytokines Associated with Cognitive Impairment in Patients with Insomnia Comorbid with Depression? A Pilot Study». Nature and Science of Sleep (em inglês). 13: 989–1000. PMC 8254552. PMID 34234602. doi:10.2147/NSS.S312272
- ↑ a b Wamsley, Erin J.; Stickgold, Robert (2019). «Dreaming of a learning task is associated with enhanced memory consolidation: Replication in an overnight sleep study». Journal of Sleep Research (em inglês). 28 (1): e12749. PMC 6338510. PMID 30091247. doi:10.1111/jsr.12749
- ↑ a b Ribeiro, Nicolas; Gounden, Yannick; Quaglino, Véronique (2021). «Enhancement of spatial memories at the associative and relational levels after a full night of sleep and likelihood of dream incorporation». International Journal of Dream Research (em inglês). 14: 67–79. doi:10.11588/ijodr.2021.1.75882
Ligações externas
- «gettinglost.ca» (em inglês)