Interação célula-célula
A interação célula-célula refere-se às interações diretas entre superfícies celulares que desempenham um papel crucial no desenvolvimento e na função de organismos multicelulares. Essas interações permitem que as células se comuniquem entre si em resposta a mudanças em seu microambiente. Essa capacidade de enviar e receber sinais é essencial para a sobrevivência da célula. As interações entre células podem ser estáveis, como aquelas feitas por meio de junções celulares. Essas junções estão envolvidas na comunicação e organização das células dentro de um tecido específico. Outras são transitórias ou temporárias, como aquelas entre células do sistema imunológico ou as interações envolvidas na inflamação dos tecidos. Esses tipos de interações intercelulares se distinguem de outros tipos, como aquelas entre as células e a matriz extracelular. A perda de comunicação entre as células pode resultar em crescimento celular incontrolável e câncer.
Interações estáveis
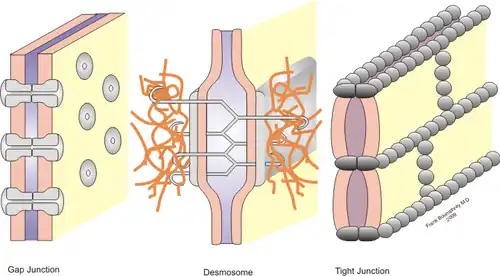
Interações célula-célula estáveis são necessárias para a adesão celular dentro de um tecido e para controlar a forma e a função das células.[1] Essas interações estáveis envolvem junções celulares, que são complexos multiproteicos que proporcionam contato entre células vizinhas. As junções celulares permitem a preservação e o funcionamento adequado das camadas de células epiteliais. Essas junções também são importantes na organização de tecidos onde células de um tipo só podem aderir a células do mesmo tecido e não a um tecido diferente.[2]
Junções estreitas
Junções estreitas são complexos multiproteicos que mantêm as células de um mesmo tecido unidas e impedem o movimento de água e moléculas solúveis em água entre as células. Nas células epiteliais, elas também funcionam para separar o fluido extracelular que envolve suas membranas apicais e basolaterais.[1] Essas junções existem como uma faixa contínua localizada logo abaixo da superfície apical, entre as membranas das células epiteliais vizinhas. As junções estreitas nas células adjacentes se alinham de modo a produzir uma vedação entre diferentes tecidos e cavidades do corpo. Por exemplo, a superfície apical das células epiteliais gastrointestinais serve como uma barreira permeável seletiva que separa o ambiente externo do corpo.[3] A permeabilidade dessas junções depende de uma variedade de fatores, incluindo a composição proteica dessa junção, o tipo de tecido e a sinalização das células.[1]
As junções estreitas são compostas de muitas proteínas diferentes. As quatro principais proteínas transmembrana são ocludina, claudina, moléculas de adesão juncional (JAMs) e tricelulinas. Os domínios extracelulares dessas proteínas formam a barreira de junção estreita, criando interações homofílicas (entre proteínas do mesmo tipo) e heterofílicas (entre diferentes tipos de proteínas) com os domínios proteicos em células adjacentes. Seus domínios citoplasmáticos interagem com o citoesqueleto celular para ancorá-los.[3]
Junções de ancoragem
Dos três tipos de junções de ancoragem, somente duas participam das interações entre células: junções aderentes e desmossomos. Ambos estão presentes em diversos tipos de células. As junções aderentes nas membranas laterais conectam as células epiteliais adjacentes. Elas estão situadas imediatamente abaixo de interseções estreitas. Sua função é fornecer forma e tensão às células e tecidos, além de serem o local onde ocorre a sinalização entre as células. As junções aderentes são compostas por moléculas de adesão celular da família das caderinas. Há mais de 100 tipos de caderinas, que correspondem a diversas células e tecidos com diferentes demandas de ancoragem.[1]
Os desmossomos também fornecem resistência e durabilidade às células e tecidos e estão localizados logo abaixo das junções aderentes. Eles são locais de adesão e não circundam a célula. Eles são feitos de duas caderinas especializadas, desmogleína e desmocolina. Essas proteínas têm domínios extracelulares que interagem entre si em células adjacentes. No lado citoplasmático, as plaquinas formam placas que ancoram os desmossomos aos filamentos intermediários compostos de proteínas de queratina. Os desmossomos também desempenham um papel na sinalização célula-célula.[4]
Junções comunicantes
As junções comunicantes são o principal local de sinalização ou comunicação célula-célula que permite que pequenas moléculas se difundam entre células adjacentes. Nos vertebrados, as junções comunicantes são compostas de proteínas transmembrana chamadas conexinas. Eles formam poros ou canais hexagonais pelos quais íons, açúcares e outras moléculas pequenas podem passar. Cada poro é feito de 12 moléculas de conexina; 6 formam um hemicanal em uma membrana celular e interagem com um hemicanal em uma membrana celular adjacente. A permeabilidade dessas junções é regulada por muitos fatores, incluindo pH e concentração de Ca2+.[1]
Proteínas receptoras na sinalização de contato direto
Proteínas receptoras na superfície celular têm a capacidade de se ligar a moléculas de sinalização específicas secretadas por outras células. A sinalização celular permite que as células se comuniquem com células adjacentes, células próximas (parácrinas) e até mesmo células distantes (endócrinas). Essa ligação induz uma mudança conformacional no receptor que, por sua vez, provoca uma resposta na célula correspondente. Essas respostas incluem mudanças na expressão genética e alterações na estrutura do citoesqueleto. A face extracelular da membrana plasmática tem uma variedade de proteínas, carboidratos e lipídios que se projetam para fora e agem como sinais. O contato direto entre as células permite que os receptores de uma célula se liguem às pequenas moléculas ligadas à membrana plasmática de outra célula. Nos eucariotos, muitas das células durante o desenvolvimento inicial comunicam-se através do contato direto.[5]
A sinalização sináptica, parte integrante da atividade do sistema nervoso, ocorre entre neurônios e células-alvo. Essas células-alvo também podem ser neurônios ou outros tipos de células (por exemplo, células musculares ou glandulares). As protocaderinas, um membro da família das caderinas, mediam a adesão dos neurônios às suas células-alvo nas sinapses, também conhecidas como junções sinápticas. Para que a comunicação ocorra entre um neurônio e sua célula-alvo, uma onda de despolarização percorre todo o comprimento do neurônio e faz com que neurotransmissores sejam liberados na junção sináptica. Esses neurotransmissores se ligam e ativam receptores no neurônio pós-sináptico, transmitindo assim o sinal para a célula-alvo. Assim, uma membrana pós-sináptica pertence à membrana que recebe o sinal, enquanto uma membrana pré-sináptica é a fonte do neurotransmissor. Em uma junção neuromuscular, uma sinapse é formada entre um neurônio motor e fibras musculares. Nos vertebrados, a acetilcolina liberada pelo neurônio motor atua como um neurotransmissor que despolariza a fibra muscular e causa contração muscular. A capacidade de um neurônio de receber e integrar sinais simultâneos do ambiente e de outros neurônios permite um comportamento animal complexo.[6]
Interações célula-célula vegetal
As células vegetais possuem paredes celulares ao seu redor, que atuam como barreiras para a comunicação entre as células. Junções especializadas conhecidas como plasmodesmos ultrapassam essa barreira. Elas se assemelham às junções comunicantes, ligando o citosol de células vizinhas. Plasmódios permitem a difusão livre de moléculas pequenas (menos de 1000 Da), como íons, aminoácidos e açúcares. Essas pequenas moléculas englobam moléculas de sinalização e fatores de transcrição. O tamanho do canal também é ajustado para acomodar moléculas de até 10.000 Da. A concentração de Ca2+ é um dos vários fatores que influenciam a permeabilidade desses canais. Um aumento na concentração de Ca2+ citosólico restringirá de forma reversível a passagem pelos plasmódios. Diferentemente das junções comunicantes, as membranas celulares das células vizinhas se fundem, criando um canal contínuo conhecido como anel. Além disso, dentro do canal, há uma extensão do retículo endoplasmático, chamada desmotúbulo, que se estende entre as células. As interações célula-célula facilitadas pelos plasmodesmos desempenham um papel importante no desenvolvimento de células e tecidos vegetais e na defesa contra infecções virais.[1]
Interações transitórias
Sistema imunológico
Leucócitos ou glóbulos brancos destroem células anormais e também fornecem proteção contra bactérias e outros materiais estranhos. Essas interações são transitórias por natureza, mas são cruciais como uma resposta imunológica imediata. Para combater a infecção, os leucócitos precisam se mover do sangue para os tecidos afetados. Esse movimento para dentro dos tecidos é chamado de extravasamento. Ela requer a formação e a quebra sucessivas de interações célula-célula entre os leucócitos e as células endoteliais que revestem os vasos sanguíneos. Essas interações célula-célula são mediadas principalmente por um grupo de moléculas de adesão celular (CAMs) chamadas selectinas.[1]
As células T auxiliares, essenciais para o sistema imunológico, interagem com outros leucócitos liberando sinais conhecidos como citocinas, que ativam e estimulam a proliferação de células B e células T assassinas. As células T auxiliares também interagem diretamente com os macrófagos, células que englobam matéria estranha e exibem antígenos em sua superfície. As células T auxiliares que possuem os receptores apropriados podem se ligar a esses antígenos e proliferar, resultando em células T auxiliares que têm a capacidade de identificar os mesmos antígenos.[7]
Coagulação
A coagulação ou coagulação sanguínea depende, além da produção de fibrina, das interações entre as plaquetas. Quando o endotélio ou o revestimento de um vaso sanguíneo é danificado, o tecido conjuntivo, incluindo as fibras de colágeno, fica exposto localmente. Inicialmente, as plaquetas aderem ao tecido conjuntivo exposto por meio de receptores específicos da superfície celular. Isso é seguido pela ativação e agregação plaquetária, na qual as plaquetas se tornam firmemente aderidas e liberam substâncias químicas que recrutam plaquetas vizinhas para o local da lesão vascular. Uma rede de fibrina se forma em torno dessa agregação de plaquetas para aumentar a resistência do coágulo.[8]
Interações celulares entre bactérias
As populações bacterianas interagem de maneira semelhante às células dos tecidos. Eles se comunicam por meio de interações físicas e moléculas de sinalização, como lactonas homoserinas e peptídeos, como um meio de controlar o metabolismo e regular o crescimento. Um exemplo comum e uma das formas mais estudadas de interação entre células bacterianas é o biofilme. Biofilme é um agregado celular que pode estar ligado a superfícies biológicas ou abióticas. As bactérias formam biofilmes para se adaptar a vários ambientes, como mudanças na disponibilidade de substrato. Por exemplo, a formação de biofilme aumenta a resistência de uma célula bacteriana aos antibióticos em comparação com as células que não fazem parte do agregado.[9]
Implicações patológicas
Câncer
O câncer pode resultar da perda da interação célula-célula. Em células normais, o crescimento é controlado pela inibição de contato, na qual o contato com células vizinhas causa uma interrupção no crescimento celular. Acredita-se que a inibição do contato seja mediada por caderinas, proteínas que desempenham um papel importante na adesão celular. Essa inibição impede que as células se acumulem umas sobre as outras e formem montes. Entretanto, em células cancerosas onde a expressão da E-caderina é perdida, a inibição de contato é perdida e resulta em crescimento ou proliferação descontrolados, formação de tumores e metástase.[10]
Patógenos bacterianos
Para que bactérias patogênicas invadam uma célula, é necessária a comunicação com a célula hospedeira. O primeiro passo para a invasão de bactérias geralmente é a adesão às células hospedeiras. A forte ancoragem, uma característica que determina a virulência, impede que as bactérias sejam eliminadas antes que a infecção ocorra. As células bacterianas podem se ligar a muitas estruturas da superfície da célula hospedeira, como glicolipídios e glicoproteínas, que servem como receptores de ligação. Uma vez ligadas, as bactérias começam a interagir com o hospedeiro para interromper seu funcionamento normal e desorganizar ou reorganizar seu citoesqueleto. Proteínas na superfície da bactéria podem interagir com receptores de proteína no hospedeiro, afetando assim a transdução de sinal dentro da célula. Alterações na sinalização são favoráveis às bactérias porque essas alterações fornecem condições sob as quais o patógeno pode invadir. Muitos patógenos têm sistemas de secreção do Tipo III que podem injetar toxinas proteicas diretamente nas células hospedeiras. Essas toxinas levam, em última análise, ao rearranjo do citoesqueleto e à entrada da bactéria.[11]
Doença
As interações célula-célula são altamente específicas e rigorosamente reguladas. Defeitos genéticos e desregulação dessas interações podem causar muitas doenças diferentes. A desregulação que leva à migração de leucócitos para tecidos saudáveis pode causar condições como a síndrome do desconforto respiratório agudo e alguns tipos de artrite.[12] A doença autoimune pênfigo vulgar é resultado de autoanticorpos contra desmogleína e outras proteínas normais do corpo. Os autoanticorpos interrompem a adesão entre as células epiteliais. Isso causa bolhas na pele e nas mucosas. Mutações nos genes da conexina causam 8 doenças humanas, incluindo malformações cardíacas e surdez neurossensorial.[1]
Notas
- Este artigo foi inicialmente traduzido, total ou parcialmente, do artigo da Wikipédia em inglês cujo título é «Cell–cell interaction».
Referências
- ↑ a b c d e f g h Harvey Lodish (2008). Molecular cell biology 6th, [2nd print] ed. New York [u.a.]: Freeman. ISBN 9780716776017
- ↑ Hausman, Geoffrey M. Cooper, Robert E. (2009). The cell : a molecular approach 5th ed. Washington, D.C.: ASM Press. ISBN 978-0-87893-300-6
- ↑ a b Suzuki T (fevereiro de 2013). «Regulation of intestinal epithelial permeability by tight junctions». Cell. Mol. Life Sci. 70 (4): 631–59. PMC 11113843
 . PMID 22782113. doi:10.1007/s00018-012-1070-x
. PMID 22782113. doi:10.1007/s00018-012-1070-x
- ↑ Dubash, AD; Green, KJ (26 de julho de 2011). «Desmosomes.». Current Biology. 21 (14): R529–31. Bibcode:2011CBio...21.R529D. PMID 21783027. doi:10.1016/j.cub.2011.04.035
- ↑ Murray P. Pendarvis; Mader, Sylvia S. (2007). Biology. Boston: McGraw-Hill Higher Education. ISBN 978-0-07-246463-4
- ↑ Wu H, Xiong WC, Mei L (abril de 2010). «To build a synapse: signaling pathways in neuromuscular junction assembly». Development. 137 (7): 1017–33. PMC 2835321. PMID 20215342. doi:10.1242/dev.038711
- ↑ Bruce Alberts (2002). Molecular biology of the cell 4. ed. New York [u.a.]: Garland. ISBN 0-8153-4072-9
- ↑ Engelmann B, Massberg S (janeiro de 2013). «Thrombosis as an intravascular effector of innate immunity». Nat. Rev. Immunol. 13 (1): 34–45. PMID 23222502. doi:10.1038/nri3345
- ↑ Voloshin SA, Kaprelyants AS (novembro de 2004). «Cell-cell interactions in bacterial populations». Biochemistry Mosc. 69 (11): 1268–75. PMID 15627380. doi:10.1007/s10541-005-0072-9
- ↑ Mayor, R; Carmona-Fontaine, C (junho de 2010). «Keeping in touch with contact inhibition of locomotion.». Trends in Cell Biology. 20 (6): 319–28. PMC 2927909. PMID 20399659. doi:10.1016/j.tcb.2010.03.005
- ↑ Lu L, Walker WA (junho de 2001). «Pathologic and physiologic interactions of bacteria with the gastrointestinal epithelium». Am. J. Clin. Nutr. 73 (6): 1124S–1130S. PMID 11393190. doi:10.1093/ajcn/73.6.1124S
- ↑ Burdick MM, McCarty OJ, Jadhav S, Konstantopoulos K (2001). «Cell-cell interactions in inflammation and cancer metastasis». IEEE Eng Med Biol Mag. 20 (3): 86–91. PMID 11446216. doi:10.1109/51.932731