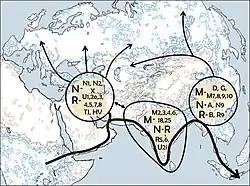
Rotas de migração propostas de haplogrupos do cromossomo Y da Eurásia Oriental (C, D, N, O e Q), durante o povoamento da Ásia Oriental.
Todos os humanos fora da África descendem de uma onda migratória que deixou o continente em que a humanidade surgiu entre 70 e 65 mil anos, tendo alcançado a Península Arábica. Dali, pelos milênios seguintes, no que ficou conhecida como dispersão austral, os humanos se dispersaram pelo Sul da Ásia, alcançando a Indochina. Foi durante essa dispersão pela Ásia às margens do Oceano Índico que ocorreu a diferenciação entre os eurasiáticos ocidentais (povos da Europa e Oriente Médio) e orientais.[1]
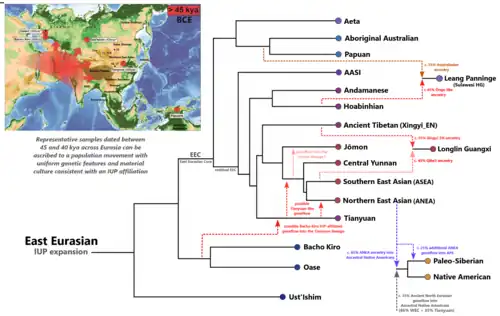
Ao longo da dispersão austral, a linhagem dos eurasiáticos orientais se trifurcou, dando origem às linhagens dos Antigos indianos ancestrais do sul (AASI em inglês, originária do Sul da Ásia), do Leste e do Sudeste Asiático (ESEA, surgida na Indochina e que migrou e povoou a China entre 55 e 50 mil anos atrás) e à dos australasianos (AA, correspondente aos aborígenes australianos, papuas e melanésios, que chegaram à região de Sahul há 60 mil anos).[1][2][3][4]
Entre 35 e 30 mil anos atrás, surgiu a variante 370A do gene EDAR, na região central da China, que é responsável por características como maior quantidade de glândulas sudoríparas, menos tecido mamário e cabelos mais grossos e está presente hoje na maioria dos habitantes da Ásia Oriental e Sudeste Asiático e também na maioria dos povos ameríndios. Essa variante genética parece ter se disseminado por causa da seleção sexual.[5][6][7]
O ramo da Ásia Oriental e Sudeste Asiático se tornaria amplamente ancestral dos modernos asiáticos orientais, povos do Sudeste Asiático (austroasiáticos e austronésios), siberianos e ameríndios. Estima-se que a população de origem da linhagem ESEA tenha se expandido a partir do Sudeste Asiático continental por volta de 42 mil anosa atrás. A divisão mais profunda dentro da linhagem ESEA deu origem aos caçadores-coletores hoabinhianos, na Indochina; à linhagem Xingyi, no sul da China; e à linhagem Tianyuan, com cerca de 39 mil anos, no norte da China e na região do rio Amur. Isso foi seguido pela divergência dos linhagem Jōmon no arquipélago japonês há cerca de 22 a 25 mil anos.[8] Então, finalmente, a divisão das ancestralidades de Yunnan central, dos antigos asiáticos orientais do sul (ASEA) e dos antigos asiáticos orientais do norte (ANEA) em algum lugar da Ásia Oriental há mais de 19 mil anos.[2] A ancestralidade ANEA substituiu as ancestralidades anterior, semelhante à de Tianyuan, na metade norte da China, na região do rio Amur e partes da Sibéria, há pelo menos 19 mil anos. A ancestralidade ASEA misturou-se com a ancestralidade Xingyi para formar a linhagem Longlin há pelo menos 11 mil anos, antes de a ancestralidade ASEA substituir a ancestralidade Longlin há cerca de seis mil anos no sul da China.[1][9]
Existem atualmente sete subancestrais identificadas no ramo do Leste e Sudeste Asiático:[1][2][3]
Antigos Asiáticos Orientais do Norte (ANEA) – Associados a populações da região do rio Amur, Mongólia, Sibéria, bem como da região do rio Amarelo até a China central. A ancestralidade pode ser amplamente dividida em três subgrupos: os antigos Asiáticos do Nordeste (ANA), também chamada de "ancestralidade do Amur", os neossiberianos" e os agricultores do Rio Amarelo. Linhagem irmã da ASEA.
Antigos Asiáticos Orientais do Sul (ASEA) – Associados a amostras antigas da região de Fujian e Guangxi, no sul da China.
Ancestralidade de Yunnan central – Ancestralidade encontrada em populações do centro de Yunnan com idades entre 5,5 mil e 1,4 mil anos. É mais relacionada às linhagens ANEA e ASEA do que às linhagens asiáticas basais, mas também distinta delas.
Ancestralidade Hoabinhiana – Ancestralidade da linhagem ESEA associada a caçadores-coletores do Laos e da Malásia com idades até 4.000 anos.
Ancestralidade Jōmon – Ancestralidade associada a indivíduos de até 3.000 anos atrás no arquipélago japonês.
Ancestralidade Tianyuan – Ancestralidade da linhagem ESEA associada a um indivíduo do Paleolítico Superior datado de aproximadamente 39 mil anos atrás no norte da China.
Árvore filogenética dos eurasiáticos orientais.Ancestralidade Xingyi – Ancestralidade associada a um indivíduo de 7,1 mil anos do sítio arqueológico de Xingyi, em Yunnan. Essa linhagem era uma população "fantasma" previamente não identificada que contribuiu para a ancestralidade Tibetana Antiga Inicial (EAT), associada a um indivíduo de Zongri de aproximadamente 6.000 anos do Planalto Tibetano.
A composição genética dos asiáticos orientais contemporâneos, como os chineses Han, é caracterizada principalmente pela presença da ancestralidade dos agricultores do rio Amarelo", um subgrupo dos antigos asiáticos orientais do norte (ANEA).[2][11] A ancestralidade dos agricultores Rio Amarelo está associada a uma amostra de um indivíduo de 9,5 mil anos da região baixa do rio Amarelo, em Shandong, formada como resultado da mistura entre um componente principal do ANEA e um componente minoritário do ASEA, ou como uma mistura anterior entre um grupo interiorano profundo (90%) e um grupo costeiro profundo (10%). A ancestralidade do Rio Amarelo é distinta da ancestralidade dos Antigos Asiáticos do Norte (ANA), mas apresenta ramificações profundas dentro do grupo dos Antigos Asiáticos Orientais do Norte (ANEA).[1][2]
Os povos contemporâneos do Nordeste da Ásia, como os tungúsicos, mongóis e túrquicos, derivam a maior parte de sua ancestralidade do subgrupo Amur do ANEA, que se expandiram massivamente com o cultivo do milhete e o pastoreio. Os povos tungúsicos exibem a maior afinidade genética com os antigos povos do nordeste asiático (ANA), representada por espécimes com cerca de 7 mil e 13 mil anos, enquanto os povos de línguas túrquicas apresentam quantidades variáveis, porém significativas, de ancestralidade da Eurásia Ocidental.[9][12]
Esquema representando as principais ancestralidades e subancestralidades na Ásia.
Um ramo inicial dos antigos asiáticos orientais do norte (ANEA) absorveu uma população dos Antigos eurasiáticos do Norte (ANE) ao norte, dando origem aos antigos paleossiberianos, que por sua vez se tornaram ancestrais tanto dos "paleossiberianos modernos" (como os falantes de línguas chukotko-kamchatkanas, ienisseianas e nivkh) quanto dos ameríndios. A ancestralidade paleossiberiana (APS) já foi disseminada por toda a Sibéria, mas foi amplamente substituída por ondas posteriores de ancestralidade neossiberiana devido a uma grande mudança populacional vinda do sul, possivelmente envolvendo falantes de línguas urálicas e yukaghir. Isso foi seguido posteriormente por outra expansão vinda do sul em tempos relativamente recentes, associada à ancestralidade antiga do nordeste da Ásia (ANA), envolvendo falantes de línguas tungúsicas, mongólicas e túrquicas.[13][14]
Os austronésios e os falantes de línguas kra–dai no Sudeste Asiático carregam principalmente a ancestralidade neolítica de Fujian, um subgrupo dos antigos asiáticos do sul (ASEA), que está associado à disseminação do cultivo de arroz.[15] Uma mistura de ancestralidade Xingyi (48,8 a 71,5%) e ancestralidade ASEA provavelmente deu origem à ancestralidade "Longlin", associada a um indivíduo de 11 mil anos de Longlin, Guangxi.[16]
Em contraste com os austronésios, a maioria dos grupos étnicos austroasiáticos demonstra maior afinidade genética com a linhagem do Yunnan Central do que com os ASEA). A ancestralidade do Yunnan central é comum entre os falantes de línguas austroasiáticas no Sudeste Asiático e na província de Yunnan, incluindo os mlabri do Laos e Tailândia, mas é baixa ou inexistente entre os falantes de línguas austroasiáticas do sul da Ásia.[17] Os caçadores-coletores isolados do Sudeste Asiático, como os semang e maniq, derivam a maior parte de sua ancestralidade das linhagens hoabinhiana.[16][18]
Populações antigas
Hoabinhianos
Os hoabinhianos representam uma sociedade tecnologicamente avançada de caçadores-coletores, que viviam principalmente na Indochina, mas também em regiões adjacentes do sul da China, entre a transição do Pleistoceno para o Holoceno e a chegada da agricultura a essa parte da Ásia. Eles estão relacionados à linhagem genética dos povos modernos da Ásia Oriental e Sudeste Asiático, embora profundamente divergente deles, sendo classificados, desse modo, como uma ancestralidade basal do Leste Asiático.[2][19]
Jōmon
O povo Jōmon representa os primeiros habitantes do arquipélago japonês e análises genéticas feitas em fósseis desse povo revelaram que eles representam uma linhagem do Leste Asiático profundamente divergente, sendo presumido que essa linhagem tenha se divergido dos antigos povos da Ásia Oriental entre 22 e 25 mil anos atrás, antes da divergência entre os Antigos Asiáticos Orientais do Norte (ANEA) e os Antigos Asiáticos Orientais do Sul (ASEA), mas depois da divergência entre a linhagem de Tianyuan, Xinyi e os hoabinhianos. Também apresentam uma fraca conexão com os Antigos Eurasiáticos do Norte.[20][21][22]
De acordo com dados compilados de vários estudos genéticos, descobriu-se que as amostras de Xiongnu têm aproximadamente 58% de ascendência do Leste Asiático, representada por uma população da idade do bronze de Khövsgöl (Mongólia), sendo o restante relacionado aos europeus e povos do Oriente Médio, representados por populações do Complexo Arqueológico Báctria-Margiana da Ásia Central e pela cultura Sintashta da Estepe Eurasiática ocidental.[25]
Linhagens paternas
Nos últimos tempos, os arqueólogos chineses publicaram várias análises sobre os resultados das escavações em Xinjiang, resultando na composição genética da classe governante suprema de Xiongnu. Particularmente interessantes são os túmulos no cemitério de Heigouliang, a leste da bacia de Barkol, perto da cidade de Hami.
Ao digitar os resultados das amostras de DNA durante a escavação de uma das tumbas, foi determinado os seguintes haplogrupos de cromossomo Y dos 12 homens: 6 Q1a* (não Q1a1-M120, não Q1a1b-M25, não Q1a2-M3), 4 Q1b-M378, 2 Q* (não Q1a, não Q1b: incapaz de determinar subclasses).[26]
Linhagens maternas
Em um estudo de restos Xiongnu permanece pertencente à necrópole de Egiin Gol no norte da Mongólia, a grande maioria (89%) das sequências mitocondriais pertencem a haplogrupos do Leste Asiático e quase 11% pertencem a haplogrupos europeus.[27]
Cinco amostras femininas de Xiongnu pertenciam aos haplogrupos mitocondriais D4b2b4, N9a2a, G3a3, D4a6 e D4b2b2b.[28]
Xianbei
Uma análise genômica completa realizada em múltiplos restos mortais dos antigos Xianbei revelou que a população derivou principalmente do conjunto genético do Nordeste Asiático Antigo (ANA), também conhecida como ancestralidade de Amur.[29] Um estudo genético de 2018 observou que o haplogrupo C2b1a1b do cromossomo Y foi detectado entre os Xianbei e os rouranos e provavelmente era uma linhagem importante entre o povo donghu.[30] Estudos genéticos publicados em 2006 e 2015 revelaram que os haplogrupos de DNA mitocondrial dos restos mortais de Xianbei eram de origem asiática oriental. De acordo com Zhou (2006), as frequências de haplogrupos maternos dos Tuoba Xianbei foram: 43,75% haplogrupo D, 31,25% haplogrupo C, 12,5% haplogrupo B, 6,25% haplogrupo A e 6,25% outros.[31] Zhou (2014) analisou o DNA mitocondrial de 17 Tuoba Xianbei e concluiu que esses espécimes eram, da mesma forma, completamente do Leste Asiático em suas origens matrilineares, pertencentes aos haplogrupos D, C, B, A, O e G.[32][33]
Populações modernas
Japoneses
Formação genético-demográfica dos japoneses
O povo Jōmon, os primeiros habitantes do arquipélago japonês, segundo a maioria dos historiadores, não eram um povo homogêneo, mas pelo menos dois ou três grupos distintos, .[34] As análises descobriram que o povo Jomon manteve um pequeno tamanho efetivo da população ao longo de vários milênios, tendo essa população se separado dos modernos asiáticos orientais há entre 17 e 22 mil anos, mas, após a divergência dos ancestrais dos ameríndios, por volta de 25 mil anos antes de Cristo, tornaram-se amplamente isolados do restante do mundo, recebendo fluxo genético apenas dos Antigos Eurasiáticos do Norte.[21][35][36][37]
Compondo 98% da população japonesa, os japoneses Yamato possuem, em média, 10% de sua ancestralidade oriunda dos autóctones Jōmon, 20% dos povos Yayoi, oriundos da Ásia Oriental continental e que chegaram ao arquipélago japonês nesse período da história do país asiático, e 70% dos Kofun, também oriundos do Leste Asiático continental e que chegaram ao arquipélago nipônico no período Kofun.[21][39] O jornal japonês The Nikkei publicou um artigo, que mostrava que a genética Kofun no Japão moderno estava concentrada em regiões específicas como Kinki, Hokuriku e Shikoku.[40]
Pesquisas recentes sugerem que a cultura Ainu se originou da fusão da cultura Okhotsk com a cultura Satsumon, ambas as quais acredita-se serem derivadas das diversas culturas do período Jōmon da história japonesa..[41][42] Em 2021, foi confirmado que o povo Jōmon de Hokkaido se formou por volta de 15 mil a.C., a partir das tribos Jōmon da ilha de Honshu e de um povo do final do Paleolítico Superior indígena de Hokkaido e do Norte da Eurásia. Os ainus, por sua vez, formaram-se a partir dos Jōmon de Hokkaido e do povo da cultura Okhotsk.[43] Os ainus são, do ponto de vista genético, a população moderna mais próxima dos antigos Jōmon que habitaram o arquipélago japonês ao longo dos milênios, herdando 70% de sua ancestralidade desses povos antigos.[21]
Os ryukyuanos constituem um grupo distinto, com análise genômica abrangente, dentro da população japonesa. São majoritariamente ligados aos antigos agricultores da Ásia Oriental Continental que povoaram o Japão em milênios recentes, mas também possuem uma quantidade relativamente alta de ancestralidade Jōmon, a qual varia segundo três diferentes estudos (28%, 36% e 26,1%).[44][45][46]
Japoneses modernos
Linhagens paternas
Segundo diversos estudos genéticos, os haplogrupo de cromossomo Y mais comuns entre os japoneses são: D-M55 ou D1b (c. 40%), O-SRY465 ou O2b (c. 29%) e O-M122 ou O3 (c. 20%).[47]
Linhagens maternas
De acordo com uma análise da amostra de japoneses do Projeto 1000 Genomas coletados na área metropolitana de Tóquio, os haplogrupos de DNA mitocondrial encontrados entre os japoneses modernos incluem D (35,6%, incluindo 33,1% D4 e 2,5% D5), B (13,6%, incluindo 9,3% B4 e 4,2% B5), M7 (10,2%), G (10,2%), N9 (8,5%), F (7,6%), A (6,8%), Z (3,4%), M9 (2,5%) e M8 (1,7%).[48]
Coreanos
Grande parte da ancestralidade dos coreanos é oriunda de populações autóctones da Península Coreana e da Manchúria, mas esse grupo étnico também possui alguma genética dos agricultores neolíticos oriundos do vale do Rio Yangtzé. Dentre os povos da Ásia Oriental, o mais próximo aos coreanos são os mongóis.[49][50][51]
Os coreanos são geralmente considerados um grupo do Nordeste Asiático, sendo, em geral, mais similares aos asiáticos do nordeste do que aos do Sudeste Asiático. Esta conclusão seria esperada a partir da correlação geral entre variação genética e geografia observada para populações humanas e é apoiada por um exame de haplogrupos individuais de DNA mitocondrial, distâncias genéticas entre populações derivadas de DNA mitocondrial ou dados do cromossomo Y e a distribuição da diversidade genética entre diferentes grupos de populações.[49]
Estudos de polimorfismos no cromossomo Y humano produziram até o momento evidências que sugerem que os coreanos têm uma longa história como um grupo étnico distinto, principalmente endogâmico, com ondas sucessivas de pessoas migrando para a península e três principais haplogrupos do cromossomo Y.[52] A população de referência para coreanos usada no Geno 2.0 Next Generation é 94% da Ásia Oriental e 5% do Sudeste Asiático e Oceania.[53]
Linguisticamente, a língua coreana mostra semelhanças com o ainu, nivkh e a maioria da línguas "altaicas", enquanto as similaridades com o japonês são explicadas por meio de uma convergência mais recente a partir do período Yayoi da história japonesa.[54] Esses grupos se correlacionam intimamente com os coreanos e japoneses modernos, que formam um grupo em comparações regionais, juntamente com vários povos tungúsicos, como os ulchis, nanais e oroqen. Usando o japonês moderno como um indicador, que mostra um sinal claro em relação às populações modernas de Taiwan, sul da China e Vietnã, o componente sulista dos coreanos pode ser estabelecido.[51] É possível observar um alto nível de continuidade genética na região ao longo dos últimos 7,7 mil anos, pelo menos. As populações modernas ainda dependem de uma série de práticas da caça, coleta e pesca e as condições climáticas frias provavelmente fornecem uma explicação para a aparente continuidade e falta de grande renovação genética por populações agrícolas forasteiras, como foi documentado no caso do Bálcãs e Europa Central.[51]
Uma possibilidade é a de que os caçadores-coletores locais paulatinamente adicionaram práticas de produção de alimentos ao seu modo de vida original.[51] Os asiáticos orientais, incluindo os grupos do norte e os do sul, estão mais próximos uns dos outros do que outros grupos populacionais e podem ter suas origens traçadas a um grupo ancestral comum que viveu há dezenas de milhares de anos.[55]
Há evidências de uma diversidade genética considerável na Coreia, incluindo níveis elevados de ascendência Jomon entre os habitantes antigos do extremo sul da Península Coreana. Foi levantada a hipótese de que a ascendência Jōmon dos antigos coreanos foi perdida ao longo do tempo, à medida que eles se misturavam continuamente com as populações que chegavam do norte da China, seguido por um período de isolamento durante o período dos Três Reinos, resultando no pool genético homogêneo dos coreanos modernos.[56][57]
Linhagens paternas
Jin Han-jun et al. (2003) afirmaram que a distribuição de haplogrupos do cromossomo Y mostra que os coreanos têm uma origem complexa que é resultado de contribuições genéticas de expansões de alcance, a maioria das quais são de uma região sul-norte do continente, e contribuições genéticas do assentamento do norte asiático.[58]
O haplogrupo de cromossomo Y mais comum entre os coreanos atuais é o Haplogrupo O-M122 (O2, anteriormente O3), que é muito antigo (tendo se formado entre 25 e 35 mil anos atrás, segundo Karmin et al. 2022[59]) e um haplogrupo de cromossomo Y frequentemente observado entre os asiáticos do leste e sudeste em geral.[60][61][62] O haplogrupo O2-M122 foi encontrado em mais de 40% dos homens coreanos amostrados.[47][50]
O segundo haplogrupo Y-DNA mais comum entre os coreanos atuais é o Haplogrupo O-M176 (O1b2, anteriormente O2b), um clado que provavelmente se espalhou principalmente de algum lugar da Península Coreana ou de suas proximidades.[63] Haplogrupo O1b2-M176 foi encontrado em aproximadamente 31% dos homens coreanos amostrados.[47]
Homens coreanos também exibem uma frequência moderada (aproximadamente 11%) do haplogrupo C-M217.[47]
Ele Miao et al. (2009) criou uma combinação artificial de partes iguais dos cromossomos Y das amostras HapMap de chineses Han em Pequim e japoneses em Tóquio. O estudo disse que esta combinação artificial se assemelhava a cinco populações que incluíam coreanos na Coreia do Sul e coreanos na China.[64]
Linhagens maternas
Estudos de DNA mitocondrial coreano mostraram que há uma alta frequência do Haplogrupo D4, variando de aproximadamente 23% entre coreanos étnicos em Arun Banner (Mongólia Interior)[65] a aproximadamente 32% entre coreanos da Coreia do Sul.[66][67] Haplogrupo D4 é o haplogrupo modal do DNA mitocondrial entre os asiáticos orientais do norte (japoneses, ryukyuanos, coreanos, manchus, Tuoba Xianbei, oroqens, evenks da Manchúria, daurs, mongóis, chineses Han do norte, tibetanos) em geral, com uma frequência máxima entre japoneses e ryukyuanos. O haplogrupo B, que ocorre com muita frequência em muitas populações do Sudeste Asiático, Polinésia e Américas, é encontrado em aproximadamente 10% (5/48 coreanos étnicos de Arun Banner) a 20% (21/103 coreanos da Coreia do Sul) dos coreanos.[65][67] O haplogrupo A foi detectado em aproximadamente 7% (7/103 coreanos da Coreia do Sul) a 15% (7/48 coreanos étnicos de Arun Banner) dos coreanos.[65][66][67] O haplogrupo A é o haplogrupo de DNA mitocondrial mais comum entre os grupos étnicos chukchi, esquimós, na-Dene e muitos grupos étnicos ameríndios da América do Norte e Central.
Manchus
Um estudo sobre a população manchu em Liaoning relatou que eles compartilham uma estreita relação genética com os chineses han do norte. Sua ancestralidade deriva, em grande parte, de um componente ancestral principal relacionado aos agricultores do Rio Amarelo e de um componente ancestral menor ligado a antigas populações da bacia do Rio Amur. Os manchus, portanto, representam uma exceção à estrutura genética típica das populações de línguas tungúsicas, provavelmente devido às migrações populacionais em larga escala e à mistura genética com os chineses han do norte nos últimos séculos.[68]
Mongóis
Os mongóis são um grupo étnico encontrado na Mongólia, Mongólia Interior e partes da Sibéria. Acredita-se que sejam descendentes dos xianbei e dos proto-mongóis. O termo anterior inclui os mongóis propriamente ditos (também conhecidos como mongóis calcas), oirates, os calmuques (encontrados na Calmúquia) e os mongóis do sul (abagas, abaganares, aohans, baarins, gorlos, jalaides, jaruuds, khishigten, khuuchids, muumyangans e onniguds). Os daurs são descendente do povo paramongólico quitai.[69] Os mongóis também estão relacionados aos manchus.
Quase todo o pool genético dos mongóis modernos é derivado de caçadores-coletores do Neolítico da região da Manchúria e da Mongólia e agricultores de painço da Ásia Oriental, com uma pequena influência genética de proto-indo-europeus e povos iranianos da Ásia Central.[70]
Linhagens paternas
A maioria dos mongóis pertence ao haplogrupo de cromossomo Y C-M217, sendo caracterizado nesse povo por uma diversidade que remonta à própria origem do haplogrupo (31 a 37 mil anos atrás[71]) e diversidade muito rasa em cada um dos subclados frequentemente observados: C-M504, C-M86, C-M407 e C-F1756.[71] Desses quatro subclados, C-M407 é filogeneticamente extremamente divergente dos outros e está mais intimamente relacionado aos subclados de C-M217 que são encontrados entre os atuais chineses, coreanos, japoneses e outros asiáticos do leste e sudeste; no entanto, entre os mongóis, C-M407 é encontrado com mais frequência no norte (entre os bargas[72] e buriates,[73] bem como nos vizinhos hamniganos e soyots[74]) e no oeste (entre Oirat Dorbet[75][76]).
Os haplogrupos O-M175 e N-M231 são encontrados em taxas médias entre os mongóis atuais. Os subclados do Haplogrupo O-M175 que foram observados entre os mongóis tendem a ser semelhantes aos encontrados entre os chineses Han, enquanto os subclados do Haplogrupo N-M231 observados entre os mongóis tendem a ser semelhantes aos encontrados entre os nenets, nganasans, cacas e tuvanos (N-B478) por um lado ou aqueles encontrados entre chukchis, coriacos e esquimós siberianos por outro (N-B197).[77]
Linhagens maternas
Os haplogrupos mitocondriais são diversos, mas semelhantes a outras populações do norte da Ásia, incluindo D, C, B e A, que são compartilhados entre populações ameríndias e asiáticas.[78]
Chineses Han
Os chineses Han do norte descendem predominantemente de populações autóctones do nordeste e centro-norte da China, recebendo pouca influência genética do Sul da China. Já os chineses Han do sul são fruto da forte miscigenação ocorrida após o Neolítico entre as populações do nordeste e centro-norte da China e do Sul da China.[36][11][79]
A contribuição estimada dos chineses Han do norte para os Han do sul é substancial nas linhagens paterna e materna e existe uma variação clinal para o DNA mitocondrial. Consequentemente, os Han do norte são os principais contribuintes para o pool genético dos Han do sul. No entanto, é importante lembrar que o processo de expansão foi dominado por homens, como é possível perceber no fato de a contribuição genética dos Han do norte para os Han do sul ser maior no cromossomo Y do que no DNA mitocondrial. Esses dados genéticos estão conformes com os registros históricos de grandes e contínuas ondas migratórias de habitantes do norte da China, escapando de guerras e fome, para o sul da China. Além dessas grandes ondas migratórias, outras migrações menores para o sul ocorreram durante quase todos os períodos dos últimos dois mil anos.[61]
Um estudo da Academia Chinesa de Ciências sobre os dados de frequência genética das subpopulações Han e minorias étnicas na China mostrou que as subpopulações Han em diferentes regiões também são geneticamente muito próximas das minorias étnicas locais, o que indica que, em muitos casos, a genética das minorias étnicas se misturou com a dos Han, enquanto, ao mesmo tempo, a genética Han também se misturou com a das minorias étnicas locais.[80]
DNA autossômico
Um estudo de 2018 calculou o FST (uma medida da diferença genética) em pares com base em SNPs de todo o genoma, entre os chineses Han (Han do norte de Pequim e do sul das províncias de Hunan, Jiangsu e Fujian), japoneses e coreanos amostrados. Descobriu-se que o menor valor de FST estava entre os Han do Norte (Pequim) (CHB) e Han do Sul (Hunan, Fujian, etc.) (CHS) (FST[CHB-CHS] = 0,0014), enquanto CHB e coreano (KOR) (FST[CHB-KOR] = 0,0026) e entre KOR e japonês (JPT) (FST[JPT-KOR ] = 0,0033). Geralmente, o FST pareado entre os chineses Han, japoneses e coreanos (0,0026~ 0,0090) é maior do que entre os chineses Han (0,0014). Esses resultados sugerem que os chineses Han, japoneses e coreanos são diferentes em termos de composição genética, e as diferenças entre os três grupos são muito maiores do que entre os Han do norte e do sul.[81]
Outro estudo mostra que os chineses Han do norte e do sul são geneticamente mais próximos um do outro e descobre também que as características genéticas dos atuais chineses Han do norte já estavam formadas antes de 1000 a.C. na Planície Central Chinesa.[82]
Um estudo genético recente sobre os restos mortais humanos (há cerca de 4000 anos) do sítio Mogou na região Gansu-Qinghai (ou Ganqing) da China revelou mais informações sobre as contribuições genéticas desse antigo povo Di-Qiang para os ancestrais dos Han do Norte. Foi deduzido que, entre 3,3 e 3,8 mil anos atrás, algumas pessoas Mogou fundiram-se na população Han ancestral, resultando na circunstância de o povo Mogou ser semelhante a alguns Han do norte e compartilhar até aproximadamente 33% de haplogrupo paterno (O3a) e cerca de 70% dos haplogrupos materno (D, A, F, M10). A taxa de mistura foi possivelmente de 13 a 18%.[83]
Um estudo mais recente, e o mais amplo de associação do genoma da população Han até agora, mostra que a estratificação geográfica-genética de norte a sul ocorreu e as populações localizadas centralmente atuam como um canal para as periféricas.[84] Em última análise, com exceção de alguns ramos etnolinguísticos dos chineses Han, como Pinghua de Guangxi, existe "estrutura genética coerente" (homogeneidade) em todos os chineses Han.[85]
Linhagens paternas
O haplogrupo de cromossomo Y O-M122 é o mais comum entre os chineses Han, sendo encontrado em 56% dos homens dessa etnia.[47] Outros haplogrupos de Y-DNA que foram encontrados com frequência notável em amostras de chineses Han incluem O-P203 (9,1-13%), C-M217 (6,0%-12,0%), N-M231 (3,6%-10,3 %), O-M268(xM95, M176) (4,7%-7,2%) e Q-M242 (1,2%-4,2%).[86]
Testes comparando os perfis genéticos de chineses Han do norte e do sul e outras etnias do sul da China determinaram que os haplogrupos O1b-M110, O2a1-M88 e O3d-M7, que são predominantes nas minorias do sul da China, foram observados apenas em alguns Han do sul (4% em média), mas não nos Han do norte. Portanto, isso prova que a contribuição masculina das minorias étnicas do sul da China nos Han do Sul é limitada, assumindo que a distribuição de frequência dos haplogrupos de cromossomo Y nas minorias do sul da China representa aquela antes da expansão da cultura Han, que começou há dois mil anos.[61]
Linhagens maternas
O DNA mitocondrial dos chineses Han aumenta em diversidade quando se olha do norte para o sul da China, assim sugerindo que migrantes do sexo masculino do norte da China se casaram com mulheres de povos locais depois de chegarem aos atuais Guangdong, Xangai, Jiangsu, Fujian e outras regiões do sul da China.[61][87]
Os haplogrupos de DNA mitocondrial dos chineses Han podem ser classificados nos haplogrupos dominantes do norte da Ásia Oriental, incluindo A, C, D, G, M8, M9 e Z, e os haplogrupos dominantes do sul da Ásia Oriental, incluindo B, F, M7, N* e R.[61]
Tibetanos
A origem dos tibetanos remonta aos habitantes Xingyi, asiáticos orientais basais, que habitavam o Planalto Tibetano há cerca de 40 a 30 mil anos, bem como aos agricultores neolíticos com ascendência da região do rio Amarelo, que migraram do norte da China nos últimos 10 mil anos e introduziram as línguas sino-tibetanas. Os tibetanos modernos derivam aproximadamente 80% de sua ancestralidade dos agricultores do Rio Amarelo, e os 20% restantes da linhagem Xingyi. O atual conjunto genético tibetano foi amplamente estabelecido há pelo menos 5,1 mil anos.[16][88][89]
Relação com hominídeos arcaicos
Após as populações oceânicas modernas, os tibetanos modernos apresentam a maior taxa de compartilhamento de alelos com espécies arcaicas do gênero Homo (mais de 6%), mostrando afinidades genéticas com três espécies: denisovanos, neandertais e uma população arcaica não identificada.[90]
Em comparação com as populações Han modernas, os tibetanos modernos mostram maior afinidade genética com os denisovanos; no entanto, tanto os Han quanto os tibetanos possuem proporções semelhantes de afinidade genética com as populações neandertais em geral.[90]
Linhagem paterna
A distribuição do Haplogrupo D-M174 do cromossomo Y é encontrada entre quase todas as populações da Ásia Central e Nordeste da Ásia ao sul da fronteira com a Rússia, embora geralmente com uma frequência baixa, nunca maior que 2%. Um pico dramático na frequência de D-M174 acontece conforme se aproxima do Planalto do Tibete. D-M174 também é encontrado em altas frequências entre os japoneses, mas diminui em baixas frequências na Península Coreana e na China. A alegação de que navajos e tibetanos estão relacionados, embora discutida entre linguistas desde Edward Sapir, não encontrou suporte em estudos genéticos. Houve algum esclarecimento sobre suas origens, no entanto, por um estudo genético, no qual foi indicado que os cromossomos Y tibetanos tinham origens múltiplas, uma da Ásia Central e outra da Ásia Oriental.[91]
Povos túrquicos
Evidências genéticas, arqueológicas e linguísticas ligam os primeiros povos túrquicos com os agricultores de painço do Nordeste da Ásia, que mais tarde adotaram um estilo de vida nômade e se expandiram para a Mongólia e o oeste.
Os povos túrquicos são uma coleção de grupos etnolinguísticos da Ásia Central, Leste Asiático, Sibéria e Oriente Próximo, bem como partes da Europa e Norte da África. Eles falam línguas relacionadas pertencentes à família túrquica.
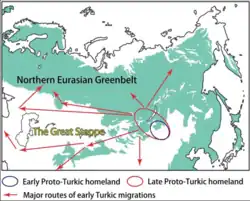
As propostas para a região de origem dos povos túrquicos e sua língua são muito diversas, indo desde a Ásia Central até a região da Manchúria.[92]
De acordo com Yunusbayev, evidências genéticas apontam para uma origem dos povos túrquicos na região dos Montes Altai e adjacências.[92]
Estudos genéticos e arqueológicos mais recentes sugerem que os povos túrquicos descendem de uma comunidade agrícola da Manchúria, que deve ser associada à cultura Xinglongwa e à cultura Hongshan que a sucedeu.[93] Por volta de 2.200 a.C., devido à desertificação da Manchúria, os ancestrais dos povos túrquicos provavelmente foram forçados a migrar para o Planalto Mongol, onde adotaram um estilo de vida pastoril.[94][95][96]
Evidências linguísticas e genéticas sugerem fortemente uma presença precoce de povos túrquicos no leste da Mongólia.[82][97] Estudos genéticos mostraram que os primeiros povos túrquicos eram de ancestralidade predominantemente do Leste Asiático. Na Alta Idade Média, os turcomanos migraram para a Ásia Central, onde se mesclaram à população local.[28][82][96][98]
No século XI, os turcos seljúcidas migraram para a Anatólia e partes do Cáucaso e começaram um processo de assimilação cultural dos nativos desses locais, levando para eles a língua e cultura túrquica.[99][100] A contribuição centro-asiática para a genética dos turcos e azeris foi pouca; os turcos, por exemplo, possuem em torno de 10% de sua genética oriunda da Ásia Central.[101]
Linhagens paternas
Os haplogrupos de cromossomo Y comuns em povos túrquicos são: o haplogrupo N-M231 (encontrado com frequência especialmente alta entre etnias turcomanas que vivem na Rússia), Haplogrupo C-M217 (especialmente na Ásia Central e, em particular, no Cazaquistão), haplogrupo Q-M242 (especialmente na região dos Montes Altai e entre turcomenos e a tribo Qangly dos cazaques) e haplogrupo O-M175 (especialmente entre os povos turcomanos que vivem na atual China, como os uigures, e a tribo Naiman dos cazaques).[82][102]
Alguns grupos também têm o haplogrupo R1b (notavelmente frequente entre os teleutes e cumandins do sul da Sibéria, os basquires e a tribo quipechaque dos cazaques), haplogrupo R1a (notavelmente frequente entre os quirguizes, altaianos e vários outros povos túrquicos que vivem na atual Rússia), haplogrupo J-M172 (especialmente frequente entre uigures, azeris e turcos) e haplogrupo D-M174 (especialmente entre os uigures amarelos, mas também observado regularmente, com baixa frequência, entre os altaianos, nogais, cazaques e uzbeques).[82][102]
Relação com outras populações da Ásia-Pacífico e ameríndios
Asiáticos centrais
Os primeiros humanos chegaram à Ásia Central a partir do Oriente Médio há mais de 40 mil anos e ali originaram a população conhecida como Antigos Eurasiáticos do Norte, geneticamente relacionados às populações da Europa e Oriente Médio. Durante a Idade do Bronze, os proto-indo-europeus (Yamnayas), pastores da Estepe Pôntica, ocuparam a Ásia Central e se mesclaram às populações que ali viviam. A genética Yamnaya passou a compor a maioria do pool genético centro-asiático.[82][103][104]
Conforme já citado, durante a Alta Idade Média, ou seja, na segunda metade do primeiro milênio depois de Cristo, os povos túrquicos migraram para a Ásia Central, aonde se mesclaram às populações locais falantes de línguas indo-europeias. As etnias túrquicas dessa parte do continente asiático, como os quirguizes, cazaques e uzbeques, possuem a maioria do seu pool genético oriundo da Ásia Oriental, sendo a exceção entre eles os turcomenos, os quais são considerados geneticamente intermediários entre os outros povos túrquicos centro-asiáticos e os povos iranianos.[82][92][105] Já os tajiques, um povo falante de uma língua da família iraniana, foram menos afetados pela chegada dos povos túrquicos à Ásia Central, de modo que mantiveram a maioria de sua genética anterior a essas ondas migratórias.[105]
Dados genéticos sugerem que a Sibéria foi povoada há aproximadamente 36 mil anos pelos Antigos Eurasiáticos do Norte.
Entre 30 e 25 mil anos atrás, no leste da Sibéria, a partir da mescla entre os Antigos Eurasiáticos do Norte e um grupo populacional oriundo da Ásia Oriental, formou-se um grupo populacional que é ancestral direto dos paleossiberianos e dos ameríndios, sendo o pool genético desse grupo ancestral de 60 a 70% oriundo dos asiáticos orientais e de 30 a 40% oriundo dos Antigos Eurasiáticos do Norte. Milênios mais tarde, os ancestrais dos indígenas da América migraram para a Beríngia, ali se isolaram e, posteriormente, povoaram as Américas. Entre 11 e 4 mil anos atrás, outros povos da Ásia Oriental migraram para a Sibéria, substituíram grande parte dos paleossiberianos e deram origem aos neossiberianos.[106][107][108][109][14][110]
Sudeste Asiático
Segundo diversos estudos genéticos, as populações modernas do Sudeste Asiático estão intimamente relacionadas às do Leste Asiático.[3][2][36][15]
Os primeiros habitantes do Sudeste Asiático são os povos genericamente conhecidos como negritos, que não são um grupo geneticamente homogêneo: os negritos das Filipinas estão geneticamente localizados entre os asiáticos orientais e os australo-papuas; já os da Indochina, conhecidos no período pré-neolítico como hoabinhianos, são relacionados à população basal dos asiáticos orientais. Atualmente, ainda habitam grupos negritos em algumas poucas regiões das Filipinas e da Península Malaia, os quais possuem parte de sua ancestralidade oriunda dos agricultores neolíticos da Ásia Oriental.[2][111][112][113][18]
Povos com origens na Ásia Oriental já habitavam o Sudeste Asiático junto com os negritos há, pelo menos, 10 mil anos.[3][114][115] Fósseis de antigos caçadores-coletores do Sudeste Asiático Marítimo, como o de uma jovem mulher que viveu há 7,2 mil anos na província indonésia de Sulawesi do Sul, tinham ascendência mista das linhagens sul-eurasiática (representada pelos papuas e aborígenes australianos) e leste-eurasiática (representada pelos asiáticos orientais e andamanesesonge). Os autores concluíram que a ancestralidade relacionada ao Leste Asiático se expandiu da Indochina para o Sudeste Asiático Marítimo muito antes do que fora sugerido anteriormente, já em 25.000 a.C.[115]
Entre cinco mil e quatro mil anos atrás, os povos agricultores austro-asiáticos e austronésios, oriundos do Sul da China, migraram para o Sudeste Asiático e substituíram as populações que anteriormente habitavam a região. Alguns grupos étnicos do Sudeste Asiático possuem alguma pouca ancestralidade das populações pré-neolíticas da região, enquanto outros, não.[36][15][114]
Na segunda metade do primeiro milênio depois de Cristo, os povos tais, sino-tibetanos e hmong-mien migraram do Sul da China para a Indochina, assim completando a diversidade étnica do Sudeste Asiático.[116]
Um estudo genético de 2022 confirmou a estreita ligação entre os povos da Ásia Oriental e do Sudeste Asiático, mas também encontrou uma proporção baixa, mas consistente, de ancestralidade associada ao Sul da Ásia (melhor amostrada pelos bengalis modernos de Dhaka, Bangladesh) entre grupos étnicos específicos da Península da Indochina (entre 2 e 16%), provavelmente como resultado da difusão cultural, principalmente de comerciantes do Sul da Ásia que espalhavam o Hinduísmo e o Budismo entre os reinos indianizados do Sudeste Asiático. Os autores, entretanto, alertam que as amostras bengalis têm ascendência substancial do Leste Asiático, o que pode afetar a estimativa de haplótipos compartilhados. No geral, estima-se que o evento de fluxo genético tenha ocorrido entre 500 e 1.000 anos atrás.[117]
Os primeiros habitantes do Sul da Ásia, apelidados de “Antigos Indianos Ancestrais do Sul” (AASI em inglês), são distantemente relacionados aos andamaneses, asiáticos orientais e aborígenes australianos. Ao longo dos milênios, povos agricultores do Irã e pastores indo-europeus da Ásia Central participaram da formação do povo sul-asiático, além de povos da Ásia Oriental, que contribuíram com a formação genética em diferentes graus em certas partes do Sul da Ásia.[104][118][119][120]
Os aborígenes australianos e papua-melanésios são geneticamente próximos aos asiáticos orientais, dos quais se divergiram há cerca de 60 mil anos, mais ou menos na época em que chegaram à Nova Guiné e Austrália. Ambos os grupos oceânicos se divergiram entre si entre 25 e 40 mil anos atrás.[2][121]
O pool genético dos povos melanésios falantes de línguas austronésias é, em grande parte, de origem papua-melanésia autóctone, com o restante oriundo da Ásia Oriental, mais especificamente das populações austronésias das Filipinas que alcançaram essa parte da Oceania entre 1.500 e 1.300 a.C.[122][123]
Os polinésios possuem entre 75 e 80% de sua ancestralidade oriunda da Ásia Oriental, ou seja, de austronésios das Filipinas, sendo o restante da ancestralidade oriunda dos papuas e melanésios.[122][123][124] Os povos do leste da Polinésia também possuem alguma ancestralidade oriunda dos povos ameríndios, dada a participação destes na colonização dessas ilhas polinésias específicas.[125]
Os micronésios também foram formados pela mistura em diferentes graus entre austronésios das Filipinas e papuas-melanésios; a ancestralidade papua-melanésia está presente nas seguintes proporções nos micronésios: quase 0% nos chamorros, quase 30% nos ilhéus de Pohnpei e Chuuk e quase 40% nos palauanos. Além disso, os chamorros possuem quase 30% de sua ancestralidade oriunda dos ameríndios e europeus, o que pode estar relacionado com a colonização espanhola desde o século XVI.[126]
↑ abcdeBennett, E. Andrew; et al. (2024). Reconstructing the Human Population History of East Asia through Ancient Genomics. [S.l.]: Cambridge University Press. 80 páginas. doi:10.1017/9781009246675 !CS1 manut: Uso explícito de et al. (link)
↑ abMao X, Zhang H, Qiao S, Liu Y, Chang F, Xie P, et al. (10 de junho de 2021). «The deep population history of northern East Asia from the Late Pleistocene to the Holocene». Cell (em inglês). 184 (12): 3256–3266.e13. PMID34048699. doi:10.1016/j.cell.2021.04.040
↑Changchun Y, Li X, Xiaolei Z, Hui Z, Hong Z (novembro de 2006). «Genetic analysis on Tuoba Xianbei remains excavated from Qilang Mountain Cemetery in Qahar Right Wing Middle Banner of Inner Mongolia». FEBS Letters (em inglês). 580 (26): 6242–6246. PMID17070809. doi:10.1016/j.febslet.2006.10.030
↑Wang H, Ge B, Mair VH, Cai D, Xie C, Zhang Q, et al. (novembro de 2007). «Molecular genetic analysis of remains from Lamadong cemetery, Liaoning, China». American Journal of Physical Anthropology (em inglês). 134 (3): 404–411. PMID17632796. doi:10.1002/ajpa.20685
↑Zhou H (março de 2014). «Genetic analyses of Xianbei populations about 1,500–1,800 years old». Human Genetics (em inglês). 50 (3): 308–314. doi:10.1134/S1022795414030119
↑ abcdYang MA, Fan X, Sun B, Chen C, Lang J, Ko YC, et al. (14 de maio de 2020). «Ancient DNA indicates human population shifts and admixture in northern and southern China». Science (em inglês). 369 (6501): 282–288. Bibcode:2020Sci...369..282Y. PMID32409524. doi:10.1126/science.aba0909
↑de Boer E, Yang MA, Kawagoe A, Barnes GL (2020). «Japan considered from the hypothesis of farmer/language spread». Evolutionary Human Sciences (em inglês). 2. ISSN2513-843X. doi:10.1017/ehs.2020.7
↑Sakitani, Mitsuru (2009). DNA, kōko, gengo no gakusai kenkyū ga shimesu shin Nihon Rettō shi: Nihonjin shūdan, Nihongo no seiritsushi. Tōkyō: Bensei Shuppan. ISBN 978-4-585-05394-1|acessodata= requer |url= (ajuda)
↑Sato T, Amano T, Ono H, Ishida H, Kodera H, Matsumura H, et al. (2007). «Origins and genetic features of the Okhotsk people, revealed by ancient mitochondrial DNA analysis». Journal of Human Genetics (em inglês). 52 (7): 618–627. PMID17568987. doi:10.1007/s10038-007-0164-z
↑Natsuki D (19 de janeiro de 2021). «Migration and adaptation of Jomon people during Pleistocene/Holocene transition period in Hokkaido, Japan». Quaternary International (em inglês). 608–609: 49–64. ISSN1040-6182. doi:10.1016/j.quaint.2021.01.009
↑Kim SH, Han MS, Kim W, Kim W (17 de agosto de 2010). «Y chromosome homogeneity in the Korean population». International Journal of Legal Medicine (em inglês). 124 (6): 653–7. PMID20714743. doi:10.1007/s00414-010-0501-1
↑Reference Populations – Geno 2.0 Next Generation. (2017). The Genographic Project. Retrieved 15 May 2017, from link.
↑Jin HJ, Kwak KD, Hammer MF, Nakahori Y, Shinka T, Lee JW, Jin F, Jia X, Tyler-Smith C, Kim W (dezembro de 2003). «Y-chromosomal DNA haplogroups and their implications for the dual origins of the Koreans». Human Genetics (em inglês). 114 (1): 27–35. PMID14505036. doi:10.1007/s00439-003-1019-0
↑Monika Karmin, Rodrigo Flores, Lauri Saag, Georgi Hudjashov, Nicolas Brucato, Chelzie Crenna-Darusallam, Maximilian Larena, Phillip L. Endicott, Mattias Jakobsson, J. Stephen Lansing, Herawati Sudoyo, Matthew Leavesley, Mait Metspalu, François-Xavier Ricaut, and Murray P. Cox (2022), "Episodes of Diversification and Isolation in Island Southeast Asian and Near Oceanian Male Lineages." Mol. Biol. Evol. 39(3): msac045 doi:10.1093/molbev/msac045
↑ abcdeWen B, Li H, Lu D, Song X, Zhang F, He Y, et al. (setembro de 2004). «Genetic evidence supports demic diffusion of Han culture». Nature (em inglês). 431 (7006): 302–305. Bibcode:2004Natur.431..302W. PMID15372031. doi:10.1038/nature02878Erro de citação: Código <ref> inválido; o nome "Wen2004" é definido mais de uma vez com conteúdos diferentes
↑ abcKong QP, Yao YG, Liu M, Shen SP, Chen C, Zhu CL, Palanichamy MG, Zhang YP (outubro de 2003). «Mitochondrial DNA sequence polymorphisms of five ethnic populations from northern China». Human Genetics (em inglês). 113 (5): 391–405. PMID12938036. doi:10.1007/s00439-003-1004-7
↑Malyarchuk BA, Derenko M, Denisova G, Woźniak M, Rogalla U, Dambueva I, Grzybowski T (junho de 2016). «Y chromosome haplotype diversity in Mongolic-speaking populations and gene conversion at the duplicated STR DYS385a,b in haplogroup C3-M407». Journal of Human Genetics (em inglês). 61 (6): 491–496. PMID26911356. doi:10.1038/jhg.2016.14
↑Har'kov VN, Hamina KV, Medvedeva OF, Simonova KV, Eremina ER, Stepanov VA (fevereiro de 2014). «[Gene pool of Buryats: clinal variability and territorial subdivision based on data of Y-chromosome markers]». Genetika (em russo). 50 (2): 203–213. PMID25711029. doi:10.1134/S1022795413110082
↑Malyarchuk B, Derenko M, Denisova G, Wozniak M, Grzybowski T, Dambueva I, Zakharov I (novembro de 2010). «Phylogeography of the Y-chromosome haplogroup C in northern Eurasia». Annals of Human Genetics (em inglês). 74 (6): 539–546. PMID20726964. doi:10.1111/j.1469-1809.2010.00601.x
↑Malyarchuk B, Derenko M, Denisova G, Khoyt S, Woźniak M, Grzybowski T, Zakharov I (dezembro de 2013). «Y-chromosome diversity in the Kalmyks at the ethnical and tribal levels». Journal of Human Genetics (em inglês). 58 (12): 804–811. PMID24132124. doi:10.1038/jhg.2013.108
↑Du R, Xiao C, Cavalli-Sforza LL (dezembro de 1997). «Genetic distances between Chinese populations calculated on gene frequencies of 38 loci». Science in China Series C: Life Sciences (em inglês). 40 (6): 613–21. PMID18726285. doi:10.1007/BF02882691
↑Gan RJ, Pan SL, Mustavich LF, Qin ZD, Cai XY, Qian J, Liu CW, Peng JH, Li SL, Xu JS, Jin L, Li H (1 de abril de 2008). «Pinghua population as an exception of Han Chinese's coherent genetic structure». Journal of Human Genetics. 53 (4): 303–13. PMID18270655. doi:10.1007/s10038-008-0250-x
↑Lu C, Zhang J, Li Y, Xia Y, Zhang F, Wu B, et al. (março de 2009). «The b2/b3 subdeletion shows higher risk of spermatogenic failure and higher frequency of complete AZFc deletion than the gr/gr subdeletion in a Chinese population». Human Molecular Genetics (em inglês). 18 (6): 1122–1130. PMID19088127. doi:10.1093/hmg/ddn427
↑Xue F, Wang Y, Xu S, Zhang F, Wen B, Wu X, et al. (23 de janeiro de 2008). «A spatial analysis of genetic structure of human populations in China reveals distinct difference between maternal and paternal lineages». European Journal of Human Genetics (em inglês). 16 (6): 705–717. PMID18212820. doi:10.1038/sj.ejhg.5201998
↑Nelson S, Zhushchikhovskaya I, Li T, Hudson M, Robbeets M (14 de fevereiro de 2020). «Tracing population movements in ancient East Asia through the linguistics and archaeology of textile production». Evolutionary Human Sciences (em inglês). 2. ISSN2513-843X. doi:10.1017/ehs.2020.4
↑Li T, Ning C, Zhushchikhovskaya IS, Hudson MJ, Robbeets M (1 de junho de 2020). «Millet agriculture dispersed from Northeast China to the Russian Far East: Integrating archaeology, genetics, and linguistics». Archaeological Research in Asia (em inglês). 22. 100177 páginas. ISSN2352-2267. doi:10.1016/j.ara.2020.100177
↑ abUchiyama J, Gillam JC, Savelyev A, Ning C (21 de maio de 2020). «Populations dynamics in Northern Eurasian forests: a long-term perspective from Northeast Asia». Evolutionary Human Sciences (em inglês). 2. ISSN2513-843X. doi:10.1017/ehs.2020.11
↑ abGenomics and Health in the Developing World (em inglês). Oxford, England: Oxford University Press. 11 de maio de 2012. pp. 1265–1267. ISBN 978-0-19-970547-4


.png)

 Muito do texto foi copiado dessa fonte, que está disponível sob uma licença
Creative Commons Attribution 2.0 (CC BY 2.0).
Muito do texto foi copiado dessa fonte, que está disponível sob uma licença
Creative Commons Attribution 2.0 (CC BY 2.0).
