Hatzegopteryx
| Hatzegopteryx | |
|---|---|
| |
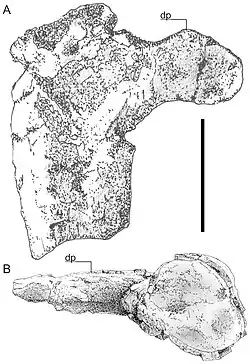
| Úmero esquerdo do espécime holótipo em vista ventral (A) e distal (B) | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Classe: | Reptilia |
| Ordem: | †Pterosauria |
| Subordem: | †Pterodactyloidea |
| Clado: | †Azhdarchoidea |
| Família: | †Azhdarchidae |
| Clado: | †Hatzegopterygia |
| Gênero: | †Hatzegopteryx Buffetaut et al., 2002 |
| Espécie-tipo | |
| †Hatzegopteryx thambema Buffetaut et al., 2002
| |
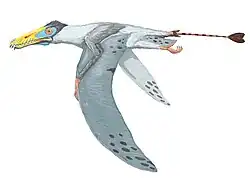
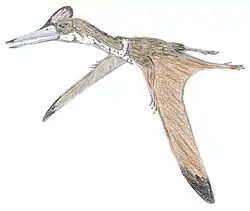
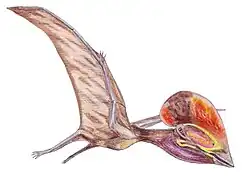
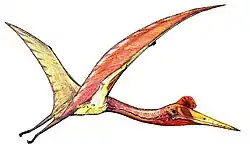
Hatzegopteryx ("asa da bacia de Hațeg") é um gênero de pterossauro da família Azhdarchidae encontrado nos depósitos do final do Maastrichtiano da Formação Densuş Ciula [en], um afloramento na Transilvânia, Romênia.[1] É conhecido apenas pela espécie-tipo, Hatzegopteryx thambema, nomeada em 2002 pelos paleontólogos Eric Buffetaut, Dan Grigorescu e Zoltan Csiki com base em partes do crânio e do úmero. Outros espécimes, incluindo uma vértebra cervical, foram posteriormente atribuídos ao gênero, representando uma variedade de tamanhos. Os maiores restos indicam que foi um dos maiores pterossauros, com uma envergadura estimada de 10 a 12 metros.
Diferentemente de outros membros gigantes de sua família, Hatzegopteryx possuía um crânio muito largo com grandes pontos de fixação muscular, ossos com textura interna esponjosa em vez de ocos e um pescoço curto, robusto e fortemente musculoso, medindo cerca de 1,5 metro de comprimento, aproximadamente metade do comprimento de outros membros de sua família com envergaduras comparáveis, capaz de resistir a fortes forças de flexão. Hatzegopteryx habitava a Ilha de Hațeg, uma ilha situada nos subtrópicos do Cretáceo no antigo Mar de Tétis. Na ausência de grandes terópodes, Hatzegopteryx era provavelmente o superpredador da Ilha de Hațeg, enfrentando presas proporcionalmente maiores (incluindo titanossauros anões e membros da infraordem Ornithopoda) do que outros indivíduos de sua família.
Descoberta e nomenclatura

Os primeiros restos de pterossauros na Romênia foram identificados por Franz Nopcsa von Felső-Szilvás em 1899, e os primeiros restos de Hatzegopteryx foram encontrados durante uma escavação estudantil no final da década de 1970, na parte superior da Formação Densuş Ciula média, em Răchitova, noroeste da Bacia de Hațeg, Transilvânia, oeste da Romênia, datada dos estágios Campaniano tardio e Maastrichtiano inicial do Cretáceo Superior, cerca de 72 milhões de anos atrás.[1] O holótipo de Hatzegopteryx, FGGUB R 1083A, consiste em dois fragmentos da parte posterior do crânio e a porção proximal danificada de um úmero esquerdo.[2] Um desses fragmentos, a região occipital, foi inicialmente atribuído a um dinossauro terópode quando anunciado em 1991.[3][4] Um segmento médio de fêmur de 38,5 cm encontrado nas proximidades, FGGUB R1625, pode também pertencer ao Hatzegopteryx.[5] O FGGUB R1625 pertenceria a um indivíduo menor de Hatzegopteryx (caso pertença ao gênero), com uma envergadura de 5 a 6 metros. Outros espécimes relatados incluem uma mandíbula não publicada, também de um indivíduo grande.[6][7]
Hatzegopteryx foi nomeado em 2002 pelo paleontólogo francês Eric Buffetaut e pelos paleontólogos romenos Dan Grigorescu e Zoltan Csiki. O nome genérico deriva da bacia de Hatzeg (ou Hațeg), na Transilvânia, onde os ossos foram encontrados, e da palavra grega pteryx (πτέρυξ), que significa “asa”. O nome específico thambema vem do grego thambema (θάμβημα), “terror, monstro”, em referência ao seu tamanho enorme.[2]
Novos espécimes de Hatzegopteryx foram recuperados de outras localidades. Na Formação Sânpetru [en], na localidade de Vadu, Sântămăria-Orlea, foi encontrado um escápulo-coracoide de tamanho médio, provavelmente de um indivíduo com envergadura de 4,5 a 5 metros. Na localidade de Râpa Roșie, da Formação Sebeș [en], contemporânea e adjacente à Formação Densuş Ciula, uma única vértebra cervical grande, o “espécime RR” ou EME 215, foi descoberta.[6][7] Embora a ausência de elementos sobrepostos impeça a atribuição definitiva deste espécime ao Hatzegopteryx thambema, sua estrutura óssea interna distinta e a falta de evidências de um segundo membro da família Azhdarchidae na área justificam sua atribuição ao menos a H. sp.[8]
Descrição
Tamanho
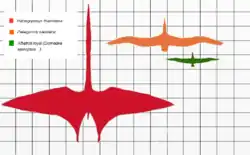
O tamanho de Hatzegopteryx foi inicialmente estimado comparando o fragmento de úmero de 236 mm com o de Quetzalcoatlus northropi, que possui um úmero de 544 mm. Observando que o fragmento de Hatzegopteryx representava menos da metade do osso original, Buffetaut e colegas estabeleceram que ele poderia ser “ligeiramente mais longo” que o de Quetzalcoatlus. A envergadura deste último foi estimada em 11 a 12 metros em 1981, embora estimativas anteriores chegassem a 15 a 20 metros. Eles concluíram que uma estimativa de 12 metros para a envergadura de Hatzegopteryx era conservadora, “desde que seu úmero fosse mais longo que o de Q. northropi”.[2][5] Em 2010, Mark P. Witton [en] e Michael Habib concluíram que Hatzegopteryx provavelmente não era maior que Q. northropi em envergadura, geralmente estimada em 10 a 11 metros.[9]
Sugere-se, com base na vértebra cervical larga e robusta atribuída ao Hatzegopteryx, que toda a coluna vertebral do animal era similarmente expandida, aumentando seu tamanho geral.[6] Contudo, isso provavelmente não é verdade, já que as vértebras cervicais de grandes pterodactiloides tendem a ser mais largas e maiores que as demais. Embora estimativas de tamanho baseadas apenas em vértebras não sejam particularmente confiáveis, o tamanho dessa vértebra é consistente com um animal de envergadura de 10 a 12 metros.[8]
Crânio
O crânio de Hatzegopteryx era gigantesco, com um comprimento estimado de 2,5 metros com base em comparações com Nyctosaurus [en] e Anhanguera, tornando-o um dos maiores crânios entre animais não marinhos.[5] O crânio era alargado na parte posterior, com 0,5 metro de largura nos ossos quadrados.[5] Enquanto a maioria dos crânios de pterossauros é composta por placas e suportes frágeis, em Hatzegopteryx, os ossos cranianos são robustos, com grandes cristas que indicam fortes pontos de fixação muscular.[2] Em 2018, Mátyás Vremir [en] concluiu que Hatzegopteryx provavelmente tinha um crânio mais curto e largo, com um comprimento estimado de 1,6 metro, e também estimou uma envergadura menor, de 8 metros.[10]
A mandíbula maciça apresentava um sulco característico no ponto de articulação (também visto em outros pterossauros, como Pteranodon), que permitia uma abertura muito ampla da boca.[2] Restos não publicados atribuídos ao Hatzegopteryx sugerem que ele tinha um bico proporcionalmente curto e profundo, agrupando-se com os membros da família Azhdarchidae de “bico rombudo” em vez dos que tinham “bico esguio”, que incluem Quetzalcoatlus sp. (agora conhecido como Q. lawsoni).[11][12]
Vértebras cervicais

Uma grande vértebra cervical atribuída ao Hatzegopteryx é curta e excepcionalmente robusta. A porção preservada mede 240 mm de comprimento, com a vértebra inteira provavelmente medindo 300 mm em vida.[6] Pterossauros tinham nove vértebras cervicais.[13] Regressões indicam que as vértebras cervicais terceira a sétima teriam medido coletivamente 1,508 metro de comprimento, com a vértebra mais longa – a quinta – medindo aproximadamente 400 mm. Em comparação, as mesmas vértebras no igualmente gigante Arambourgiania [en] mediam 2,652 metros. Isso indica que o pescoço de Hatzegopteryx tinha cerca de 50–60% do comprimento esperado para um membro da família Azhdarchidae de seu tamanho.[8]
A superfície inferior da vértebra cervical também era excepcionalmente espessa, com 4 a 6 mm. Na maioria dos outros gigantes da família, incluindo Arambourgiania, essa superfície tem menos de 2,6 mm de espessura. Embora a espinha neural da vértebra não esteja completamente preservada, a largura da porção preservada sugere que era relativamente alta e robusta em comparação com a de outros pterossauros. Outros aspectos da vértebra se assemelham mais à sétima vértebra cervical do menor Azhdarcho [en]: os soquetes articulares são muito mais rasos que os arcos neurais e quatro vezes mais largos que altos; um processo na base das vértebras, conhecido como hipapófise, está presente; os processos na frente das vértebras, as prezigapófises, são abertos; e a vértebra tem uma “cintura” afilada no centro.[8] Embora inicialmente identificada como uma terceira vértebra cervical,[6] essas características suportam sua identificação como proveniente da parte posterior do pescoço, especificamente como a sétima vértebra.[8]
Classificação
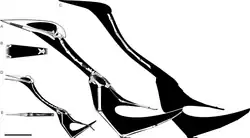
Semelhanças entre o úmero de Hatzegopteryx e de Quetzalcoatlus northropi foram notadas, pois ambos possuem uma crista deltopeitoral longa e lisa e uma cabeça umeral espessada. Essas características foram inicialmente a base para a atribuição do táxon ao clado Azhdarchidae,[2] mas também são semelhantes o suficiente para justificar a sinonímia entre Hatzegopteryx e Quetzalcoatlus. Contudo, isso provavelmente se deve à natureza relativamente não diagnóstica do úmero na taxonomia de gigantes da família e à falta de uma descrição detalhada dos elementos de Q. northropi na época da atribuição.[14] No entanto, a anatomia do pescoço e da mandíbula de Hatzegopteryx é claramente distinta do menor Q. lawsoni, o que justifica a manutenção de Hatzegopteryx como um táxon separado de Quetzalcoatlus.[2][8][15]
A vértebra cervical atribuída ao Hatzegopteryx sp. contém características que permitem identificá-la definitivamente como de um membro de Azhdarchidae. O centro é relativamente baixo, as zigapófises são grandes e achatadas, e as porções preservadas da espinha neural indicam que ela é bífida, ou dividida ao meio.[8] Uma análise filogenética conduzida pelo paleontólogo Nicholas Longrich e colegas em 2018 posicionou Hatzegopteryx em uma posição derivada (avançada) dentro de Azhdarchidae.[16] Essa posição é corroborada por análises filogenéticas subsequentes de Brian Andres em 2021 e de Rodrigo Pêgas e colegas em 2023. Ambos encontraram o Hatzegopteryx dentro da subfamília Quetzalcoatlinae, embora em posições diferentes. Andres encontrou-o em um clado com Arambourgiania e Quetzalcoatlus, enquanto Pêgas e colegas o recuperaram como grupo irmão de Albadraco [en], outro pterossauro encontrado na Bacia de Hațeg.[17][18] Seus cladogramas estão apresentados abaixo:

|
Topologia 1: Andres (2021).
|
Topologia 2: Pêgas e colegas (2023).
|
Paleobiologia

Estrutura óssea
Embora o crânio de Hatzegopteryx fosse excepcionalmente grande e robusto, seus ossos das asas são comparáveis aos de outros pterossauros voadores, indicando que não era incapaz de voar. Buffetaut e colegas sugeriram que, para voar, o peso do crânio de Hatzegopteryx deveria ser reduzido de alguma forma. Essa redução de peso pode ter sido alcançada pela estrutura interna dos ossos cranianos, cheios de pequenos poros e cavidades (alvéolos) de até 10 mm, separados por uma matriz de finas traves ósseas (trabéculas). Os ossos das asas também apresentam uma estrutura interna semelhante. Essa construção incomum difere da de outros pterossauros e se assemelha mais à estrutura do poliestireno expandido (usado para fabricar isopor). Isso tornava o crânio resistente e robusto, mas também leve, permitindo o voo.[2] Uma estrutura interna semelhante é observada na vértebra cervical atribuída ao Hatzegopteryx.[8]
Biomecânica do pescoço
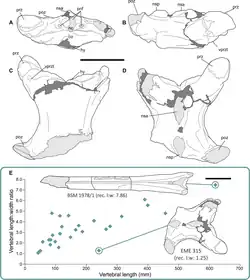
Devido às suas vértebras robustas e de paredes espessas, o pescoço de Hatzegopteryx era muito mais forte que o de Arambourgiania. Isso pode ser quantificado usando a força relativa de falha, que é a força de ruptura de um osso dividida pelo peso corporal do pterossauro, estimado em 180 a 250 kg para Arambourgiania e Hatzegopteryx. Enquanto as vértebras cervicais de Arambourgiania falham com cerca de metade de seu peso corporal, as vértebras cervicais posteriores de Hatzegopteryx podem suportar entre cinco e dez vezes seu peso corporal, dependendo da carga no osso. Mesmo as vértebras cervicais anteriores hipoteticamente mais longas de Hatzegopteryx poderiam suportar quatro a sete vezes o peso corporal.[8]

Embora o centro de Hatzegopteryx seja muito mais robusto que o da Arambourgiania, suas proporções de raio ósseo para espessura óssea (R/t)[9] são aproximadamente iguais (9,45 para Hatzegopteryx e 9,9 para Arambourgiania). Isso pode representar um compromisso entre aumentar a resistência à flexão e a resistência à flambagem. Proporções R/t mais altas aumentam a resistência à flexão, mas enfraquecem a resistência à flambagem. Para compensar, o Hatzegopteryx apresenta adaptações para melhorar a resistência à flambagem, como as estruturas internas distintas dos ossos e as grandes articulações das vértebras, que ajudam a distribuir o estresse.[8]
Para suportar a cabeça robusta, o pescoço de Hatzegopteryx era provavelmente fortemente musculoso. Nos ossos occipitais, as linhas nucais, que servem como pontos de fixação muscular, são muito desenvolvidas e apresentam cicatrizes proeminentes. Essas provavelmente sustentavam os músculos transversoespinhais, que auxiliam na extensão e flexão da cabeça e do pescoço. Da mesma forma, o processo opistótico, as espinhas neurais e as zigapófises pareciam ser grandes e robustas (com estas últimas apresentando muitos poros e bordas que provavelmente representam cicatrizes musculares), e as tuberosidades basioccipitais eram longas. Todos esses servem como pontos de fixação para vários músculos da cabeça e do pescoço. Embora não completamente desmusculado, o pescoço de Arambourgiania provavelmente não era tão musculoso quanto o de Hatzegopteryx.[8]
Paleoecologia

Como todos os pterossauros de sua família, Hatzegopteryx era provavelmente um predador generalista que forrageava em terra.[19] Ele é significativamente maior que qualquer outro predador terrestre do Maastrichtiano na Europa. Devido ao seu grande tamanho em um ambiente dominado por dinossauros anões, na ausência de grandes terópodes hipercarnívoros na região, sugere-se que Hatzegopteryx desempenhava o papel de superpredador no ecossistema da Ilha de Hațeg. Sua anatomia robusta sugere que ele enfrentava presas maiores que outros membros de sua família, incluindo animais grandes demais para serem engolidos inteiros.[19] Enquanto isso, outros gigantes de sua família, como Arambourgiania, provavelmente se alimentavam de presas pequenas (até o tamanho de um humano), incluindo filhotes ou dinossauros pequenos e ovos.[8] Outro pterossauro, Thalassodromeus, também foi sugerido como ave de rapina.[20]
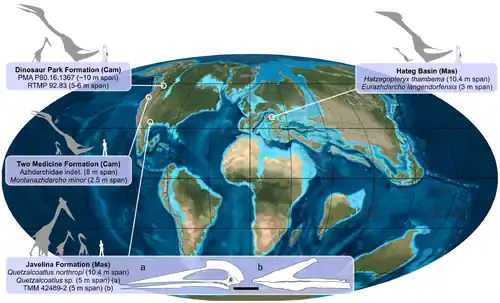
Além de Hatzegopteryx, o ecossistema da Ilha de Hațeg abrigava outros habitantes incomuns. Pterossauros coexistentes incluíam Eurazhdarcho [en], com envergadura de 3,8 metros,[7] um dinossauro de pescoço curto não nomeado, de tamanho pequeno, com envergadura de 3,5 a 4 metros, um dinossauro um pouco maior, também não nomeado, com envergadura de 5 metros, e aparentemente pequenos indivíduos da família Pteranodontidae.[21] O robusto, sem voo e possivelmente herbívoro do clado Avialae[22] ou dromeossaurídeo[23] Balaur bondoc, com duas garras enlargadas em cada pé, representa outro componente altamente especializado da fauna. O ecossistema continha vários anões insulares, como os titanossauros Magyarosaurus [en][24] e Paludititan [en],[25] o hadrossaurídeo Telmatosaurus e o iguanodontiano Zalmoxes.[24] Junto com o nodossaurídeo Struthiosaurus [en], vários indivíduos pequenos e fragmentários do clado Maniraptora estavam presentes, incluindo Bradycneme [en], Elopteryx [en] e Heptasteornis [en].[24] Restos de crocodilianos, pertencentes aos gêneros Allodaposuchus, Doratodon [en] e Acynodon, também foram encontrados.[26] Componentes não arcossaurianos incluem os mamíferos da família Kogaionidae [en] multituberculados Kogaionon [en], Barbatodon [en], Litovoi tholocephalos [en] e Hainina [en],[27][28] lagartos como o teiídeo Bicuspidon [en] e Becklesius (família Paramacellodidae [en]), uma cobra não nomeada da família Madtsoiidae, e os lissanfíbios Albanerpeton [en], Eodiscoglossus [en] e Paradiscoglossus [en].[29]
A importância dessa fauna é uma justificativa geológica significativa para a designação da área, entre 2004 e 2005, como Geoparque dos Dinossauros do País de Hațeg, um dos primeiros membros da Rede Europeia de Geoparques, e (quando a designação de Geoparques Mundiais da UNESCO foi ratificada em 2015) como Geoparque Mundial UNESCO de Hațeg.[30]
Durante o Maastrichtiano, o sul da Europa era um arquipélago. Os membros do ecossistema da Ilha de Hațeg viviam em uma massa terrestre conhecida como Bloco Tisia–Dacia, da qual a Bacia de Hațeg era uma pequena parte. Essa massa terrestre tinha cerca de 80.000 km² e era separada de outros terrenos terrestres por trechos de oceano profundo em todas as direções por 200 a 300 km.[24] Localizada a 27°N,[31] a ilha estava mais ao sul que a latitude atual de 45°N. Assim, o clima era provavelmente subtropical, com estações seca e úmida distintas, e uma temperatura média de cerca de 25°C.[32] O ambiente consistia de várias planícies aluviais, zonas úmidas e rios,[33] cercados por bosques dominados por samambaias e angiospermas.[24] Paleossolos indicam um clima cretáceo relativamente seco, com precipitação anual inferior a 1.000 mm.[32]
Referências
- ↑ a b Albert, Gáspár; Budai, Soma; Csiki-Sava, Zoltán; Makádi, László; Ţabără, Daniel; Árvai, Valentin; Bălc, Ramona; Bindiu-Haitonic, Raluca; Ducea, Mihai N.; Botfalvai, Gábor (1 de junho de 2025). «Age and palaeoenvironmental constraints on the earliest dinosaur-bearing strata of the Densuș-Ciula Formation (Hațeg Basin, Romania): Evidence of their late Campanian-early Maastrichtian syntectonic deposition». Cretaceous Research. 170. 106095 páginas. Bibcode:2025CrRes.17006095A. ISSN 0195-6671. doi:10.1016/j.cretres.2025.106095

- ↑ a b c d e f g h Buffetaut, E.; Grigorescu, D.; Csiki, Z. (2002). «A new giant pterosaur with a robust skull from the latest Cretaceous of Romania» (PDF). Naturwissenschaften. 89 (4): 180–184. Bibcode:2002NW.....89..180B. PMID 12061403. doi:10.1007/s00114-002-0307-1
- ↑ Weishampel, D.B.; Barrett, P.M.; Coria, R.A.; Le Loeuff, J.; Xu, X.; Zhao, X.; Sahni, A.; Gomani, E.M.P.; Noto, C.M. (2004). «Dinosaur Distribution». In: Weishampel, D.B.; Dodson, P.; Osmolska, H. The Dinosauria 2nd ed. [S.l.]: University of California Press. p. 593. ISBN 9780520254084
- ↑ Witton, M.P. (2017). «New paper: when the short-necked, giant azhdarchid pterosaur Hatzegopteryx ruled Late Cretaceous Romania». Markwitton.com Blog
- ↑ a b c d Buffetaut, E.; Grigorescu, D.; Csiki, Z. (2003). «Giant azhdarchid pterosaurs from the terminal Cretaceous of Transylvania (western Romania)». Geological Society, London, Special Publications. 217 (1): 91–104. Bibcode:2003GSLSP.217...91B. doi:10.1144/GSL.SP.2003.217.01.09
- ↑ a b c d e Vremir, M.M. (2010). «New faunal elements from the Late Cretaceous (Maastrichtian) continental deposits of Sebeș area (Transylvania)» (PDF). Acta Musei Sabesiensis. 2: 635–684
- ↑ a b c Vremir, M.; Kellner, A.W.A.; Naish, D.; Dyke, G.J. (2013). «A New Azhdarchid Pterosaur from the Late Cretaceous of the Transylvanian Basin, Romania: Implications for Azhdarchid Diversity and Distribution». PLOS ONE. 8 (1): e54268. Bibcode:2013PLoSO...854268V. PMC 3559652. PMID 23382886. doi:10.1371/journal.pone.0054268
- ↑ a b c d e f g h i j k l Naish, D.; Witton, M.P. (2017). «Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators». PeerJ. 5: e2908. PMC 5248582. PMID 28133577. doi:10.7717/peerj.2908
- ↑ a b Witton, M.P.; Habib, M.B. (2010). «On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness». PLOS ONE. 5 (11): e13982. Bibcode:2010PLoSO...513982W. PMC 2981443. PMID 21085624. doi:10.1371/journal.pone.0013982
- ↑ Vremir, Mátyás; Dyke, Gareth; Csiki-Sava, Zoltán; Grigorescu, Dan; Buffetaut, Eric (2018). «Partial mandible of a giant pterosaur from the uppermost Cretaceous (Maastrichtian) of the Hațeg Basin, Romania». Lethaia. 51 (4): 493–503. doi:10.1111/let.12268
- ↑ Andres, B.; Langston, W. Jr. (2021). «Morphology and taxonomy of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea)». Journal of Vertebrate Paleontology. 41 (sup1): 142. Bibcode:2021JVPal..41S..46A. ISSN 0272-4634. doi:10.1080/02724634.2021.1907587
- ↑ Witton, M.; Brusatte, S.; Dyke, G.; Naish, D.; Norell, M.; Vremir, M. (2013). Pterosaur overlords of Transylvania: short-necked giant azhdarchids in Late Cretaceous Romania. The Annual Symposium of Vertebrate Paleontology and Comparative Anatomy. Edimburgo. Consultado em 18 de janeiro de 2017. Arquivado do original em 6 de abril de 2016
- ↑ Bennett, S.C. (2014). «A new specimen of the pterosaur Scaphognathus crassirostris, with comments on constraint of cervical vertebrae number in pterosaurs». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 271 (3): 327–348. Bibcode:2014NJGPA.271..327B. doi:10.1127/0077-7749/2014/0392
- ↑ Witton, M.P.; Martill, D.M.; Loveridge, R.F. (2010). Clipping the Wings of Giant Pterosaurs: Comments on Wingspan Estimations and Diversity. Proceedings of the Third International Symposium on Pterosaurs. Acta Geoscientica Sinica. 31. Pequim. pp. 79–81. Consultado em 18 de janeiro de 2017. Arquivado do original em 31 de janeiro de 2017
- ↑ Witton, M.P. (2016). «Quetzalcoatlus: the media concept vs the science». Markwitton.com Blog
- ↑ Longrich, N.R.; Martill, D.M.; Andres, B.; Penny, D. (2018). «Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary». PLOS Biology. 16 (3): e2001663. PMC 5849296. PMID 29534059. doi:10.1371/journal.pbio.2001663
- ↑ Andres, Brian (7 de dezembro de 2021). «Phylogenetic systematics of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea)». Journal of Vertebrate Paleontology. 41 (sup1): 203–217. Bibcode:2021JVPal..41S.203A. ISSN 0272-4634. doi:10.1080/02724634.2020.1801703
- ↑ Pêgas, R. V.; Zhoi, X.; Jin, X.; Wang, K.; Ma, W. (2023). «A taxonomic revision of the Sinopterus complex (Pterosauria, Tapejaridae) from the Early Cretaceous Jehol Biota, with the new genus Huaxiadraco». PeerJ. 11. e14829. PMC 9922500. PMID 36788812. doi:10.7717/peerj.14829
- ↑ a b Witton, M.P.; Naish, D. (2015). «Azhdarchid pterosaurs: water-trawling pelican mimics or "terrestrial stalkers"?». Acta Palaeontologica Polonica. 60 (3). doi:10.4202/app.00005.2013
- ↑ Witton, M.P. (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton: Princeton University Press. p. 243. ISBN 9780691150611
- ↑ Vremir, M.; Witton, M.; Naish, D.; Dyke, G.; Brusatte, S.L.; Norell, M.; Totoianu, R. (2015). «A medium-sized robust-necked azhdarchid pterosaur (Pterodactyloidea: Azhdarchidae) from the Maastrichtian of Pui (Hațeg Basin, Transylvania, Romania)». American Museum Novitates (3827): 1–16. ISSN 0003-0082. doi:10.1206/3827.1. hdl:2246/6582
- ↑ Cau, A.; Brougham, T.; Naish, D. (2015). «The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc (Dinosauria, Maniraptora): dromaeosaurid or flightless bird?». PeerJ. 3: e1032. PMC 4476167. PMID 26157616. doi:10.7717/peerj.1032
- ↑ Csiki, Z.; Vremir, M.; Brusatte, S.L.; Norell, M.A. (2010). «An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania». Proceedings of the National Academy of Sciences of the United States of America. 107 (35): 15357–15361. Bibcode:2010PNAS..10715357C. PMC 2932599. PMID 20805514. doi:10.1073/pnas.1006970107
- ↑ a b c d e Benton, M.J.; Csiki, Z.; Grigorescu, D.; Redelstorff, R.; Sander, P.M.; Stein, K.; Weishampel, D.B. (2010). «Dinosaurs and the island rule: The dwarfed dinosaurs from Hațeg Island». Palaeogeography, Palaeoclimatology, Palaeoecology. 293 (3): 438–454. Bibcode:2010PPP...293..438B. doi:10.1016/j.palaeo.2010.01.026
- ↑ Cziki, Z.; Codrea, V.; Jipa-Murzea, C.; Godefroit, P. (2010). «A partial titanosaur (Sauropoda, Dinosauria) skeleton from the Maastrichtian of Nălaţ-Vad, Hațeg Basin, Romania». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 258 (3): 297–324. Bibcode:2010NJGPA.258..297C. doi:10.1127/0077-7749/2010/0098
- ↑ Martin, J.E.; Csiki, Z.; Grigorescu, D.; Buffetaut, E. (2006). «Late Cretaceous crocodilian diversity in Hațeg Basin, Romania». Hantkeniana. 5 (5): 31–37
- ↑ Csiki, Z.; Grigorescu, D. (2000). «Teeth of multituberculate mammals from the Late Cretaceous of Romania». Acta Palaeontologica Polonica. 45 (1): 85–90
- ↑ Codrea, V.A.; Solomon, A.A.; Venzcel, M.; Smith, T. (2017). «Première identification d'une espèce de mammifère du Crétacé supérieur du bassin de Rusca Montană (Transylvanie, Roumanie)» [Primeira identificação de uma espécie de mamífero do Cretáceo Superior da Bacia de Rusca Montană (Transilvânia, Romênia)]. Comptes Rendus Palevol. 16 (1): 27–38. doi:10.1016/j.crpv.2016.04.002
- ↑ Folie, A.; Codrea, V. (2005). «New lissamphibians and squamates from the Maastrichtian of Hațeg Basin, Romania». Acta Palaeontologica Polonica. 50 (1): 57–71
- ↑ «Geoparque Global UNESCO de Hațeg». Consultado em 25 de março de 2023
- ↑ Panaiotu, C.G.; Panaiotu, C.E. (2010). «Palaeomagnetism of the Upper Cretaceous Sânpetru Formation (Hațeg Basin, South Carpathians)». Palaeogeography, Palaeoclimatology, Palaeoecology. 293 (4): 343–352. Bibcode:2010PPP...293..343P. doi:10.1016/j.palaeo.2009.11.017
- ↑ a b Therrien, F. (2005). «Palaeoenvironments of the latest Cretaceous (Maastrichtian) dinosaurs of Romania: insights from fluvial deposits and paleosols of the Transylvanian and Hațeg basins» (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 218 (1): 15–56. Bibcode:2005PPP...218...15T. doi:10.1016/j.palaeo.2004.12.005
- ↑ Therrien, F.; Zelenitsky, D.K.; Weishampel, D.B. (2009). «Palaeoenvironmental reconstruction of the Late Cretaceous Sânpetru Formation (Hațeg Basin, Romania) using paleosols and implications for the "disappearance" of dinosaurs». Palaeogeography, Palaeoclimatology, Palaeoecology. 272 (1): 37–52. Bibcode:2009PPP...272...37T. doi:10.1016/j.palaeo.2008.10.023
Ligações externas
 Media relacionados com Hatzegopteryx no Wikimedia Commons
Media relacionados com Hatzegopteryx no Wikimedia Commons
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||