Eichstaettisaurus
Eichstaettisaurus
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Titoniano-Albiano | |||||||||||||
.jpg) Espécime tipo e único de E. schroederi | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Espécie-tipo | |||||||||||||
(Broili, 1938) | |||||||||||||
| Outras espécies | |||||||||||||
| |||||||||||||
| Sinónimos | |||||||||||||
E. schroederi
| |||||||||||||
Eichstaettisaurus (que significa "lagarto de Eichstätt") é um gênero de lagartos do Jurássico Superior e Cretáceo Inferior encontrado na Alemanha, Espanha e Itália. Com uma cabeça achatada, pés orientados para a frente e parcialmente simétricos, e garras altas, Eichstaettisaurus apresentava muitas adaptações a um estilo de vida escalador, semelhantes às das lagartixas.
A espécie-tipo, E. schroederi, está entre os membros mais antigos e completos dos Squamata, sendo conhecida por um espécime originário da formação Solnhofen do período Titoniano na Alemanha. Uma segunda espécie, E. gouldi, foi descrita a partir de outro esqueleto encontrado nas montanhas de Matese, na Itália. Apesar de muito semelhante a E. schroederi, viveu muito mais tarde, durante o estágio Albiano. Os fósseis de ambas as espécies apresentam preservação excepcional devido à deposição em ambientes marinhos de baixo oxigênio.
Inicialmente nomeada como uma espécie do gênero Ardeosaurus por Ferdinand Broili em 1938, E. schroederi teve uma história taxonômica complexa. Foi inicialmente reconhecida como parente dos lagartos noturnos Xantusiidae, até que pesquisas na década de 1960 a identificaram, junto com Ardeosaurus, como lagartixas. No entanto, análises filogenéticas realizadas na década de 1990 sugeriram que ela era mais próxima do ancestral de todos os esquamatas. Várias posições conflitantes foram encontradas até o surgimento de análises incorporando mais espécies e melhores dados, que resolveram E. schroederi como um parente próximo das lagartixas no grupo Gekkonomorpha. Contudo, a posição de Ardeosaurus em relação a E. schroederi permanece incerta: uma espécie, A. digitatellus, foi recuperada como parente próxima em algumas análises, mas a outra espécie, A. brevipes, foi considerada mais distantemente relacionada.
Descoberta e nomenclatura
E. schroederi

Em 1938, Ferdinand Broili descreveu um espécime de lagarto muito bem preservado, em posição dorsal, proveniente de depósitos rochosos do Jurássico no município de Wintershof, Eichstätt, Alemanha. O espécime foi adquirido pela Coleção Estatal de Paleontologia e Geologia Histórica da Bavária (BSPG), onde foi preparado por L. Haimerl. Consiste em um esqueleto quase completo, embora com o crânio esmagado, uma cauda ausente após a quinta vértebra (provavelmente resultado de amputação e substituição por cartilagem), e várias vértebras preservadas apenas como impressões (mas sua presença pôde ser verificada pelas costelas associadas). Uma contraplaca do espécime também existe.[1] Atualmente, os depósitos rochosos são conhecidos como pertencentes à Formação Altmühltal [en], e o espécime e sua contraplaca estão armazenados sob os números de acesso na coleção BSPG como 1937 I 1a e 1b.[2]
Embora não pudesse examiná-lo diretamente, Broili notou, a partir da descrição de Christian Hermann von Meyer do espécime tipo de Ardeosaurus brevipes, que ele apresentava semelhanças com este espécime, mas também diferenças importantes na forma do focinho e do osso frontal, bem como na contagem de vértebras. No entanto, hesitou em criar um novo gênero devido ao estado de preservação do crânio. Assim, atribuiu-o ao gênero Ardeosaurus com reservas, como uma nova espécie, A.? schröderi, nomeada em homenagem ao seu colega J. Schröder.[1] (O atual nome específico é schroederi: conforme o Artigo 32.5.2.1 do Código Internacional de Nomenclatura Zoológica, tremas em nomes alemães publicados antes de 1985 devem ser removidos e substituídos por um "e" inserido como a próxima letra.)[3]
Robert Hoffstetter reconheceu formalmente a distinção entre A.? schroederi e A. brevipes em 1953, e estabeleceu um novo nome genérico, Broilisaurus. No entanto, Oskar Kuhn já havia nomeado um anfíbio Stegocephalia com o mesmo nome em 1938; assim, em 1958, ele estabeleceu outro nome genérico para a espécie, Eichstättisaurus. (Novamente, devido ao ICZN, o nome genérico atual é Eichstaettisaurus.) Em 1963, Marguerite Cocude-Michel opinou que E. schroederi realmente representava a mesma espécie que Homoeosaurus digitatellus, que havia sido nomeada como uma espécie não-tipo de Homoeosaurus em 1914 por N.M. Grier. Assim, ela criou a nova combinação Eichstaettisaurus digitatellus.[4][5][6]
Ao contrário de Cocude-Michel, Hoffstetter identificou digitatellus como um membro de Ardeosaurus separado de Eichstaettisaurus em 1964. Mesmo ciente do artigo de Hoffstetter, em 1965 Cocude-Michel reiterou seu argumento sem tentar responder a Hoffstetter. Hoffstetter respondeu em 1966, chamando a sinonímia de Cocude-Michel de "insuportável". Ele observou que isso implicaria a revisão das características diagnósticas de Eichstaettisaurus a um ponto em que seria praticamente indistinguível de Ardeosaurus, o que contradiria sua separação em nível de família.[6] A literatura subsequente manteve E. schroederi e A. digitatellus como separados, embora tenham recebido pouca atenção até que Simões e colegas publicaram uma redescrição de ambos em 2017.[2]
E. gouldi

Por muitos anos, Eichstaettisaurus foi conhecido apenas pelo único espécime de E. schroederi até que restos do Cretáceo foram encontrados em outras partes da Europa. Em 2000, Susan Evans [en] e colegas descreveram um pequeno espécime de lagarto na localidade de Berriasiano–Valanginiano de Montsec, na Catalunha, Espanha. Apesar de sua preservação precária, o espécime era claramente distinto do lagarto mais comum na localidade, Meyasaurus [en]; Evans e colegas sugeriram que ele apresentava afinidades com Eichstaettisaurus.[7]
Em 2004, Evans e colegas relataram restos ainda mais jovens de Eichstaettisaurus, originários do Albiano da "Pietraroja Plattenkalk" na localidade de Pietraroja, localizada nas Montanhas Matese, no sul da Itália. Um espécime catalogado como MPN 19457 formou o espécime tipo de uma nova espécie, E. gouldi, que eles nomearam em homenagem ao falecido biólogo Stephen Jay Gould; é um esqueleto preservado em posição ventral, que inclui o crânio e partes da coluna vertebral e membros. Um segundo espécime consiste em um crânio desarticulado, preservado como parte do conteúdo intestinal de um lagarto Rhynchocephalia indeterminado catalogado como MPN A01/82. Ambos os espécimes estão armazenados no Museu de Paleontologia da Universidade de Nápoles (MPN).[4]
Descrição
Eichstaettisaurus era um animal relativamente pequeno. E. schroederi tinha um comprimento rostro-cloacal (medido da ponta do focinho até a abertura da cloaca) de 94 mm,[2] enquanto E. gouldi tinha um comprimento rostro-cloacal de 56 mm.[4]
Crânio
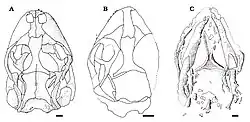
Eichstaettisaurus pode ser identificado por crânios achatados com focinhos curtos, rombos e arredondados. E. schroederi tinha narinas grandes. Os ossos portadores de dentes do focinho, a pré-maxila e a maxila, eram provavelmente conectados por uma faixa de tecido mole em E. schroederi. A pré-maxila tinha 6 ou 7 dentes em E. gouldi, mas o número em E. schroederi é desconhecido; a maxila tinha aproximadamente 22 dentes em E. schroederi, e pelo menos 30 em E. gouldi. Os dentes de ambas as espécies eram pequenos, estreitos, cônicos e com uma única cúspide; eles tinham uma implantação pleurodonte, ou seja, estavam fundidos ao lado interno do osso da mandíbula.
Como Ardeosaurus, ambas as espécies não possuíam o osso lacrimal, geralmente parte da órbita ocular. Também na órbita ocular, os ossos pós-frontal e osso pós-orbital eram separados, ao contrário de Ardeosaurus, e a projeção posterior desses ossos era relativamente larga. Em E. schroederi, o contato entre esses ossos era mais anterior do que na maioria dos lagartos. Os ossos esquamosais eram delgados e projetavam-se para baixo para encontrar os ossos quadrados, como na maioria dos lagartos; ao contrário dos Iguania e Teiidae, no entanto, eles não possuíam projeções ascendentes. As lagartixas vivas não possuem os ossos pós-orbital e esquamosal, e também têm ossos jugais relativamente mais curtos do que Eichstaettisaurus. No palato de E. schroederi, os ossos ectopterigoides sobrepunham os ossos pterigoides em sua parte posterior.[2][4]
Ao contrário de A. brevipes, E. schroederi não possuía ornamentação óssea e osteodermos no teto craniano. Em ambas as espécies, os ossos frontais eram fundidos e mais estreitos entre as órbitas oculares do que na frente. As projeções descendentes nos frontais, conhecidas como processos subolfativos, eram bem desenvolvidas em ambas as espécies, mas se encontravam na linha média apenas em E. schroederi. A sutura entre os ossos frontal e parietal era convexa em E. schroederi e côncava em E. gouldi, ao contrário de Ardeosaurus, onde era reta. E. gouldi e a maioria dos outros lagartos têm ossos parietais fundidos, mas eles eram pareados e não fundidos em E. schroederi. Os parietais de ambas as espécies não possuíam projeções posteriores e fossas nucais. Em E. schroederi, as bordas externas dos parietais curvavam-se para dentro, e as projeções posteriores, conhecidas como processos supratemporais, eram curtas, amplamente separadas e apresentavam depressões. Também em E. schroederi, um par de cristas estava presente no osso supraoccipital da caixa craniana, provavelmente marcadas pelos canais semicirculares devido à reduzida ossificação do crânio.[2][4]
Vértebras e costelas
Comparado à maioria dos lagartos iguanianos, gekkotanos e scincomorphanos, que geralmente têm 24–26 vértebras à frente do sacro (ou quadril), E. schroederi tinha 31, incluindo 7 vértebras cervicais e aproximadamente 24 vértebras torácicas.[2] E. gouldi tinha 6–7 vértebras cervicais e um número desconhecido de vértebras dorsais.[4] Em E. gouldi, a segunda (áxis) e a terceira vértebras cervicais eram conectadas à seguinte por elementos conhecidos como intercentros.[4] Em E. schroederi, a maioria das vértebras cervicais não possuía costelas cervicais, exceto as duas últimas. As espinhas neurais projetadas para cima nas vértebras dorsais eram pequenas, e as últimas vértebras dorsais não possuíam costelas no estilo das vértebras lombares. Articulações "zigosfeno-zigantrum" estão presentes nas vértebras dorsais de ambas as espécies. As costelas eram de cabeça única e tinham superfícies de articulação circulares.[2] Em E. gouldi, os centros (corpos principais) das vértebras caudais eram largos, cilíndricos, amficélicos (côncavos em ambas as extremidades) e apresentavam processos transversais fortemente desenvolvidos.[4]
Membros e suas articulações
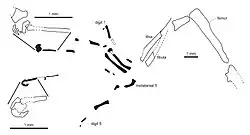
Eichstaettisaurus tinha um processo acrômio bem desenvolvido, ou seja, o ponto de fixação da clavícula, na escápula (omoplata). As clavículas de E. schroederi não se curvavam para a frente, ao contrário da maioria dos membros dos Autarchoglossa [en], e também não possuíam projeções posteriores. Como em Ardeosaurus, ambas as espécies de Eichstaettisaurus tinham membros anteriores relativamente robustos.[2][4] Em E. schroederi, o processo olecrano [en] da ulna era bem desenvolvido.[2] Em E. gouldi, os ossos carpais provavelmente não estavam completamente ossificados.[4] A mão de cinco dedos de E. schroederi apresentava 2, 3, 4, 5 e 3 falanges (ossos dos dedos) em cada dedo; as penúltimas falanges das pontas eram relativamente longas.[2] E. gouldi parece ter tido 4 falanges no quarto dedo, e suas garras eram curtas, profundas e afiadas, com tubérculos flexores bem desenvolvidos.[4]
Na pelve de E. schroederi, a expansão dos púbis no avental púbico fundido era relativamente estreita; os púbis também não possuíam as projeções anteriores conhecidas como tubérculo púbico. Os ísquios eram aumentados e entravam em contato extensivamente, e a expansão posterior da lâmina do ílio era longa. Ambas as espécies de Eichstaettisaurus tinham membros posteriores robustos, de comprimento semelhante aos anteriores. O fêmur tinha um eixo reto, ao contrário da maioria dos outros lagartos, onde é sigmoide. No pé, o primeiro osso metatarso era ligeiramente mais curto que o segundo ao quarto ossos metatarsais. Inusitadamente, o quinto metatarso era muito mais curto e em forma de gancho. Em E. schroederi, o pé era de cinco dedos, como as mãos, com 2, 3, 4, 5 e 4 falanges em cada dedo, e as penúltimas falanges e garras eram semelhantes. E. schroederi compartilhava duas características das garras do pé com a atual lagartixa-tokay: as garras tinham extremidades inferiores expandidas; e as articulações entre as falanges eram juntas côncavo-convexas simples, em vez de bicondilares (ou seja, baseadas em duas projeções arredondadas nas extremidades das falanges), ao contrário da maioria dos outros lagartos.[2][4]
Classificação

A história filogenética de Eichstaettisaurus foi tumultuada, assim como a de Ardeosaurus, consequência tanto de dados incompletos quanto de sua forma primitiva.[2][8] von Meyer não forneceu uma colocação para Ardeosaurus quando nomeou o gênero; Richard Lydekker e Karl Alfred von Zittel colocaram-no, junto com Homoeosaurus, nos Rhynchocephalia, Barão Franz Nopcsa colocou Ardeosaurus como parente próximo dos escincídeos, e Charles Camp [en] colocou-o próximo aos Gekkonidae.[9][10][11] Com base nas bordas bem desenvolvidas das órbitas oculares e fenestras supratemporais no crânio do espécime tipo de E. schroederi, Broili concordou com a conclusão de Nopcsa em 1938. Dentro dos Scincomorpha, ele observou que as vértebras amficélicas de Ardeosaurus (incluindo E. schroederi) se assemelhavam mais às dos Xantusiidae (lagartos noturnos), já que a maioria dos outros escincídeos tem vértebras procelosas (convexas na superfície posterior).[1]
Ao separar schroederi no gênero Broilisaurus, Hoffstetter manteve-o dentro dos Xantusiidae em 1953, mas notou que seu crânio curiosamente se assemelhava aos das lagartixas. Como xantusiídeos e lagartixas eram então colocados nos dois principais ramos reconhecidos de lagartos, Autarchoglossa e Ascalabota, Hoffstetter especulou que ele estava próximo ao estoque ancestral dos lagartos.[5] Ao renomeá-lo para Eichstaettisaurus, Kuhn estabeleceu a família Eichstaettisauridae, ainda nos Xantusiidae. Em 1961 e depois em 1963, Cocude-Michel concordou com Camp que Ardeosauridae (também extinto) e Eichstaettisauridae eram realmente lagartixas e não xantusiídeos, com base em grandes diferenças no esqueleto pós-craniano, e colocou ambos no clado Gekkonomorpha.[5] Hoffstetter aceitou essa identificação em 1966,[6] assim como Richard Estes em 1983.[2]
No entanto, análises filogenéticas conduzidas ao longo do final dos anos 1990 sugeriram que tanto Eichstaettisaurus quanto Ardeosaurus eram squamatas basais (divergindo cedo). Junto com Bavarisaurus, Scandensia e Hoyalacerta, os dois foram encontrados fora do grupo coroa de Squamata. Isso os tornaria squamatas-tronco, ou seja, não descendentes do ancestral comum de todos os squamatas vivos.[2][12][13] No entanto, em uma análise de 1999, pesquisadores reconheceram que a posição de Eichstaettisaurus era instável devido a dados ausentes (apenas 44% das características usadas em sua análise podiam ser codificadas para ele), e uma de suas árvores filogenéticas o colocou como o grupo irmão de Scleroglossa entre os squamatas coroa.[14] Em 2004 descobriram que E. gouldi era o grupo irmão de E. schroederi, e recuperaram uma posição semelhante para ambos.[4]
Análises filogenéticas subsequentes encontraram uma variedade de posições conflitantes para Eichstaettisaurus e Ardeosaurus. Em 2005, Evans e Wang conduziram duas análises, uma baseada na análise de Lee de 1998 e outra baseada na análise de Evans e Chure de 1998, mas adicionaram Dalinghosaurus [en], Dorsetisaurus e Parviraptor a ambas. Uma encontrou Eichstaettisaurus como o grupo irmão de um grupo contendo Xantusiidae, Gekkonidae, Pygopodidae e Amphisbaenia, com Ardeosaurus próximo a Lacertidae e Teiioidea; a outra uniu Eichstaettisaurus com Dalinghosaurus e os Xenosauridae nos Anguimorpha, com Ardeosaurus como o grupo irmão de Iguania.[15] Após Evans, Wang e Li adicionarem Yabeinosaurus [en] às mesmas análises, uma encontrou Eichstaettisaurus, Ardeosaurus, Yabeinosaurus e Parviraptor formando o grupo irmão de Scleroglossa, enquanto a outra recuperou Ardeosaurus como um squamata-tronco e Eichstaettisaurus, Hoyalacerta, Parviraptor e Scandensia formando um grupo entre Scleroglossa.[16]
Uma análise filogenética de 2006 por Conrad e Norell também encontrou Eichstaettisaurus próximo a Scleroglossa.[17] Em 2008, Conrad publicou uma grande análise filogenética de squamatas, incluindo 222 taxa e 363 características, que tentou resolver o problema de amostragem taxonômica incompleta (ou seja, um número insuficiente de táxons incluídos) causando instabilidade filogenética. Para incluir Eichstaettisaurus, ele combinou dados de E. schroederi e E. gouldi. Ele descobriu que tanto Eichstaettisaurus quanto Ardeosaurus, junto com Bavarisaurus, faziam parte do grupo-tronco de Scleroglossa. Junto com o grupo coroa de Scleroglossa, eles compunham o clado Scincogekkonomorpha.[18] Bolet e Evans conduziram duas análises baseadas na análise de Conrad, para as descrições de 2010 e 2012 de Pedrerasaurus e Jucaraseps, e recuperaram posições semelhantes para Eichstaettisaurus. No entanto, a primeira continha uma politomia, uma vez que a relação entre Iguania, Pedrerasaurus, um grupo contendo Ardeosaurus, Yabeinosaurus e Sakurasaurus, e todos os outros squamatas não pôde ser resolvida.[19][20]
Jacques Gauthier e colegas publicaram outra grande análise filogenética em 2012, contendo 192 táxons e 976 características. Como Conrad, eles também combinaram dados de E. schroederi e E. gouldi. Eles recuperaram uma posição mais derivada para Eichstaettisaurus como parte do grupo-tronco de Gekkota, junto com o espécime sem nome AMNH FR 21444; em particular, eles notaram que suas proporções de membros se assemelhavam fortemente às dos gekkos.[8] Após adicionar Ardeosaurus digitatellus e recodificar Eichstaettisaurus para incluir apenas dados de E. schroederi, Simões e colegas encontraram E. schroederi em uma posição semelhante, embora em uma politomia com A. digitatellus e AMNH FR 21444. Eles identificaram várias características de gekkos: os parietais pareados; o sulco meckeliano fechado na mandíbula inferior; os dentes maxilares pequenos e cônicos; o focinho curto e rombo e o crânio achatado; os ectopterigoides sobrepondo os pterigoides; os dois pterigoides e ossos palatinos sendo amplamente separados; a ampla sutura dividindo os pterigoides e palatinos uns dos outros; e as vértebras semelhantes a lombares.[2]
Em 2018, Tałanda adicionou A. brevipes à análise de Gauthier e colegas (mas não incluiu as revisões de Simões e colegas). Ele encontrou Eichstaettisaurus na mesma localização ao lado de Norellius [en], mas também recuperou A. brevipes como um scincomorfo intimamente relacionado aos escincídeos (em contraste com a posição de tronco-gekkotan de A. digitatellus). Apesar de suas diferentes posições filogenéticas, Tałanda observou que as duas espécies diferem apenas pela largura de seus parietais. Ele especulou que dados incompletos devido à má preservação de A. digitatellus poderiam ter influenciado esses resultados, e assim ele o reteve no gênero Ardeosaurus.[21] Outra análise conduzida por Simões, Tałanda e colegas em 2018, desta vez usando os dados revisados de Simões e colegas, encontrou uma posição semelhante para E. schroederi, mas uma mais basal para A. brevipes nos Autarchoglossa.[22] Em 2019, Alifanov considerou tanto Eichstaettisaurus quanto Ardeosaurus como scincomorfos intimamente relacionados a Carusia [en], mas não a Xenosauridae, mas não realizou uma análise filogenética.[23]
Abaixo, são mostradas árvores filogenéticas das duas principais análises filogenéticas incluindo Eichstaettisaurus: uma baseada no estudo de Conrad (Bolet & Evans, 2012, com relações dentro de Scleroglossa de Conrad, 2008),[18][20] e uma baseada na pesquisa de Gauthier e colegas (Simões e colegas, 2018).[22]
Topologia A: Bolet & Evans (2012) com Scleroglossa de Conrad (2008)[18][20]
| Squamata |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Topologia B: Simões et al. (2018)[22]
| Squamata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia

As lagartixas modernas são incomuns entre os lagartos, pois os dígitos de seus membros são relativamente simétricos em comprimento e dispostos em um arco amplo; em contraste, os dígitos de outros lagartos geralmente são quase paralelos entre si, especialmente nos pés. O padrão observado nas lagartixas facilita a aderência enquanto o corpo está em várias orientações, uma vez que espalha as setas [en] adesivas (cerdas) em suas almofadas digitais, permitindo que o primeiro e o último dígito se oponham um ao outro.[24][25] E. schroederi apresentava duas características que contribuíam para a simetria do pé: seu primeiro metatarso era longo em relação ao terceiro metatarso (com o terceiro metatarso sendo apenas 1,41 vezes mais longo que o primeiro, em oposição a 2 vezes em um lagarto típico), e seu quarto metatarso era relativamente curto. Em 2017, Simões e colegas observaram que E. schroederi tinha uma simetria do pé mais forte que Ardeosaurus digitatellus, e inferiram que os pés de ambos provavelmente eram mais direcionados para a frente do que em outros lagartos. Isso também foi corroborado pela ausência de articulações bicondilares nas falanges, que são uma adaptação para pés voltados para fora, resistindo ao deslocamento no plano horizontal enquanto permitem flexão no plano vertical.[2]
Os pesquisadores também identificaram várias outras características em E. schroederi, sugerindo que os estilos de vida escaladores (baseados em escalada) das lagartixas modernas surgiram antes do que se pensava anteriormente. Suas garras altas e falanges alongadas na penúltima posição dos dígitos são características fortemente correlacionadas com estilos de vida escalador em lagartos.[26][27][28][29][30] Em particular, as garras altas podem ter proporcionado braços de alavanca mais longos para os tendões flexores que as retraem, melhorando sua força de aderência.[31] Enquanto isso, seus membros relativamente curtos e corpo achatado podem ter melhorado o desempenho na escalada ao reduzir seu centro de gravidade, como foi sugerido para a lagartixa-tokay,[31] mas essa característica pode não estar correlacionada com estilos de vida escaladores.[32] Finalmente, seus membros de comprimentos semelhantes podem ter melhorado a aderência, como no lagarto Dalmatolacerta oxycephala [en]; a relevância dessa característica pode ser diminuída devido às almofadas adesivas das lagartixas, e à falta de correlação entre a proporção do comprimento dos membros e a capacidade de escalada em Lacertidae.[2][32]
Em 2004, Evans e colegas também discutiram o estilo de vida de E. gouldi à luz de suas proporções corporais. Eles apontaram um estudo anterior sobre como a variação nas proporções entre espécies de lagartos da neve (Carinascincus [en]) estava correlacionada com o uso do habitat: troncos curtos e membros posteriores longos estavam correlacionados com escalada em rochas (em espécies grandes) e escalada em árvores (em espécies pequenas), enquanto troncos longos e membros posteriores mais curtos estavam correlacionados com a vida no solo.[33] Evans e colegas descobriram que E. gouldi era mais próximo das espécies que vivem no solo, que têm velocidades de corrida lentas e são relativamente pobres em escalada, em suas proporções. No entanto, eles reconheceram que os corpos achatados e garras altas de E. gouldi eram adaptados para se agarrar a superfícies rugosas. Eles propuseram um estilo de vida híbrido para E. gouldi: um lagarto terrestre de movimento lento com alguma capacidade de escalada em rochas e de se esconder em fendas de predadores como rincocéfalos.[4]
Paleoecologia
E. schroederi
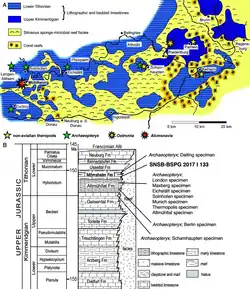
As unidades rochosas em Wintershof que produziram o único espécime conhecido de E. schroederi fazem parte dos calcários de Solnhofen (também conhecido como formação Altmühltal) do sul da Alemanha, que são bem conhecidos por seus fósseis excepcionalmente preservados.[2] Historicamente, a estratigrafia dos calcários de Solnhofen tem sido objeto de considerável confusão, com a variedade de ambientes e padrões de deposição contribuindo para a confusão.[34] Eles são agora entendidos como consistindo de várias formações geológicas no Grupo Weißjura: no sentido mais estrito, eles consistem na formação Altmühltal a noroeste de Ingolstadt (incluindo localidades ao redor de Eichstätt), e na formação Painten a nordeste de Ingolstadt. Dentro da Formação Altmühltal, a pedreira de Wintershof faz parte do Membro Eichstätt Superior, que, em termos de bioestratigrafia de amonites, está no horizonte Euvirgalithacoceras eigeltingense-β entre as subzonas de Lithacoceras riedense e Hybonoticeras hybonotum.[35][36] Os depósitos da formação Altmühltal, que foram datados do estágio inferior do Titoniano do período Jurássico, consistem de "plattenkalk" (mudas baseadas em calcário muito finamente granulado, como micrite [en]) formando camadas finas e uniformes com cerca de 1 cm de espessura que geralmente não apresentam evidências de bioturbação (perturbação por organismos vivos).[37][38][39]
Durante o Titoniano, o plattenkalk da formação Altmühltal foi depositado em bacias oceânicas (chamadas "wannen") dentro de um mar raso e quente ao redor de um arquipélago.[39] Essas bacias, que podem ter sido lagunas, tinham uma paleolatitude de aproximadamente 34° N, e estavam localizadas na margem norte do oceano de Tétis. Fósseis de animais que viviam no fundo, como estrelas-do-mar e gastrópodes, são praticamente ausentes, o que sugere que as condições no fundo do mar eram inóspitas à vida; isso pode ter sido causado por vários fatores, incluindo hipersalinidade, depleção de oxigênio ou acúmulo de sulfeto de hidrogênio tóxico.[38][40] Essas condições foram responsáveis pela preservação excepcional de fósseis como o de E. schroederi,[41] que teriam sido predominantemente transportados da terra circundante durante chuvas fortes e tempestades.[42] Em terra, a ausência de samambaias e a predominância de coníferas adaptadas a ambientes secos sugerem que o clima era semiárido.[43]
.jpg)
Ambos A. brevipes e A. digitatellus foram descobertos em Wintershof com E. schroederi,[2] junto com Homoeosaurus maximiliani e o rincocéfalo Pleurosaurus goldfussi.[5] Também conhecidos de Wintershof estão o pterossauro Rhamphorhynchus muensteri,[44] o crocodiliforme Alligatorellus bavaricus,[45] e vários animais aquáticos: os peixes Anaethalion angustus,[46] Ascalabos voithii,[47] Aspidorhynchus acutirostris,[48] Belonostomus spyraenoides,[49] Caturus giganteus,[50] Gyrodus circularis,[51] Macrosemius rostratus, Palaeomacrosemius thiollieri,[52] Propterus elongatus,[53] e Zandtfuro tischlingeri;[54] o cação-anjo Pseudorhina alifera;[55] os coleoides semelhantes a lulas Acanthoteuthis problematica,[56] Belemnotheutis mayri,[57] e Plesioteuthis prisca;[58] o crinóide Saccocoma tenella, muito comum nos depósitos de Solnhofen;[59] o camarão Dusa reschi;[60] e o caranguejo-ferradura Mesolimulus walchi.[61] Pedreiras próximas produziram o espécime de Eichstätt do avialano dinossauro Archaeopteryx lithographica,[62] e os pterossauros Aerodactylus scolopaciceps, Germanodactylus cristatus e possivelmente Cycnorhamphus [en].[63]
E. gouldi

A localidade de Pietraroja, de onde são conhecidos os espécimes de E. gouldi, é formada por um afloramento de plattenkalk no topo de uma encosta rochosa (a "Civita di Pietraroia") localizada na margem leste das montanhas Matese. Essas montanhas começaram a se formar apenas no Mioceno, com a maior parte do levantamento tectônico que as trouxe à sua elevação atual ocorrendo entre o Plioceno e o Pleistoceno. Na localidade, há dois leitos distintos de plattenkalk; o leito inferior é mais grosseiro e praticamente desprovido de fósseis, enquanto o leito superior é mais argiloso e é a fonte da maioria dos fósseis. Os dois leitos são separados por calcários lagoonais. O plattenkalk é formado por camadas de packstone [en] e grainstone [en] com 2 - 25 cm de espessura, que se alternam com camadas de lamito, wackestone [en] e marga; o leito superior também é caracterizado pela presença de espículas de esponja, cherte, betume e coprólitos.[4][64]
Com base em foraminíferos, a localidade de Pietraroja foi datada da época Albiano do Cretáceo Inferior. Durante esse período, o plattenkalk de Pietraroja teria sido depositado em uma plataforma carbonatada de águas rasas próxima a uma pequena ilha. O ambiente de deposição foi originalmente pensado como uma lagoa,[65] mas foi reinterpretado como um canal subaquático com fluxo para o leste que foi gradualmente preenchido durante o Aptiano, com base nos padrões de disposição dos fósseis, correntes de água e transporte de sedimentos. A falta de bioturbação também aponta para um ambiente anóxico, possivelmente relacionado a um evento anóxico global. Alguns fósseis estão bem preservados, enquanto outros foram quase destruídos, sugerindo que eles se originaram de vários locais, com animais terrestres como E. gouldi sendo trazidos para o canal por fluxos de água doce.[4][64] O clima teria sido tropical a subtropical.[65]

Além de E. gouldi e do rincocéfalo MPN A01/82, outros lepidosauros de Pietraroia incluem os lagartos Chometokadmon fitzingeri e Costasaurus rusconi, e o rincocéfalo Derasmosaurus pietraroiae, bem como um terceiro espécime juvenil não nomeado.[4][66][67] O fóssil mais conhecido de Pietraroia é o do dinossauro compsognatídeo Scipionyx samniticus [en], que preserva tecidos moles, incluindo órgãos internos.[68] Outros répteis incluem dois espécimes de crocodiliformes pertencentes a Pietraroiasuchus ormezzanoi,[69] e o anfíbio albanerpetontídeo Celtedens megacephalus.[70]
Muitos peixes são conhecidos de Pietraroia, com o mais comum sendo um pycnodontídeo [en] anteriormente identificado como Coelodus costae mas agora conhecido como Gregoriopycnodus bassanii.[71] Outros incluem Anaethalion robustus, Belonostomus crassirostris, Caeus leopoldi, Cavinichthys pietrarojae, Chirocentrites coroninii, parentes de Diplomystus brevissimus e Elopopsis fenzii, Hemieloposis gibbus, Ionoscopus petrarojae, Italophiopsis derasmoi, uma espécie de Lepidotes, Notagogus pentlandi, Pleuropholis decastroi, Propterus scacchii e Sauropsidium laevissimum.[72][73][74][75][76] Peixes cartilaginosos incluíam o tubarão-anjo Phorcynis e a raia Rhinobatus obtusatus.[72] Invertebrados incluíam rudistas, bivalves, caracóis marinhos do gênero Nerinea, camarões e estrelas-do-mar.[64][65]
Referências
- ↑ a b c Broili, F. (1938). «Ein neuer fund von ?Ardeosaurus H. von Meyer» (PDF). Sitzungsberichte der Mathematisch-Naturwissenschaftlichen Abteilung der Bayerischen Akademie der Wissenschaften zu München: 97–114. Consultado em 14 de maio de 2020. Cópia arquivada (PDF) em 22 de fevereiro de 2022
- ↑ a b c d e f g h i j k l m n o p q r s Simões, T.R.; Caldwell, M.W.; Nydam, R.L.; Jiménez-Huidobro, P. (2017). «Osteology, phylogeny, and functional morphology of two Jurassic lizard species and the early evolution of scansoriality in geckoes». Zoological Journal of the Linnean Society. 180 (1): 216–241. doi:10.1111/zoj.12487
- ↑ Snitting, D.; Blom, H. (2009). «Correcting taxon names containing diacritics — examples from Paleozoic vertebrates». Journal of Vertebrate Paleontology. 29 (1): 269–270. Bibcode:2009JVPal..29..269S. doi:10.1080/02724634.2009.10010377
- ↑ a b c d e f g h i j k l m n o p q Evans, S.E.; Raia, P.; Barbera, C. (2004). «New lizards and rhynchocephalians from the Lower Cretaceous of southern Italy» (PDF). Acta Palaeontologica Polonica. 49 (3): 393–408. Consultado em 11 de dezembro de 2017. Cópia arquivada (PDF) em 9 de agosto de 2017
- ↑ a b c d Cocude−Michel, M. (1963). «Les rhynchocéphales et les sauriens des calcaires lithographiques (Jurassique supérieur) d'Europe occidentale». Nouvelles Archives de la Muséum d'Histoire Naturelle de Lyon. 7: 143–185
- ↑ a b c Hoffstetter, R. (1966). «A propos des genres Ardeosaurus et Eichstaettisaurus (Reptilia, Sauria, Gekkonoidea) du Jurassique Supèrieur de Franconie». Bulletin de la Société Géologique de France. 8 (4): 592–595. doi:10.2113/gssgfbull.S7-VIII.4.592. Consultado em 14 de maio de 2020. Cópia arquivada em 7 de junho de 2020
- ↑ Evans, S.E.; Ruiz, A.L.; Rey, J. (2000). «A lizard from the Early Cretaceous (Berriasian-Valanginian) of Montsec, Catalonia, Spain». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 215: 1–15. doi:10.1127/njgpa/215/2000/1
- ↑ a b Gauthier, J.A.; Kearney, M.; Maisano, J.A.; Rieppel, O.; Behlke, A.D.B. (2012). «Assembling the Squamate Tree of Life: Perspectives from the Phenotype and the Fossil Record». Bulletin of the Peabody Museum of Natural History. 53 (1): 3–308. doi:10.3374/014.053.0101
- ↑ Lydekker, R. (1888). «Order Rhynchocephalia». Catalogue of the Fossil Reptilia and Amphibia in the British Museum (Natural History). 1. London: British Museum of Natural History. pp. 290–301. doi:10.5962/bhl.title.61848. Consultado em 15 de maio de 2020. Cópia arquivada em 6 de junho de 2020
- ↑ Nopcsa, F. (1908). «Zur Kenntnis der fossilen Eidechsen» [To the knowledge of fossil lizards]. Beiträge zur Paläontologie und Geologie Österreich-Ungarns und des Orients. 21: 33–62
- ↑ Camp, C. (1923). «Classification of the lizards». Bulletin of the American Museum of Natural History. 48: 289–481. hdl:2246/898
- ↑ Evans, S.; Barbadillo, L.J. (1998). «An unusual lizard (Reptilia: Squamata) from the Early Cretaceous of Las Hoyas, Spain». Zoological Journal of the Linnean Society. 124 (3): 235–265. doi:10.1006/zjls.1997.0139

- ↑ Evans, S.E.; Chure, D.C. (1998). «Paramacellodid lizard skulls from the Jurassic Morrison Formation at Dinosaur National Monument, Utah». Journal of Vertebrate Paleontology. 18 (1): 99–114. Bibcode:1998JVPal..18...99E. doi:10.1080/02724634.1998.10011037
- ↑ Evans, S.E.; Barbadillo, L.J. (1999). «An short-limbed lizard from the Lower Cretaceous of Spain». Special Papers in Palaeontology. 60: 73–85
- ↑ Evans, S.E.; Wang, Y. (2005). «The Early Cretaceous lizard Dalinghosaurus from China» (PDF). Acta Palaeontologica Polonica. 50: 725–742
- ↑ Evans, S.E.; Wang, Y.; Li, C. (2005). «The early Cretaceous lizard genus Yabeinosaurus from China: Resolving an enigma». Journal of Systematic Palaeontology. 3 (4): 319–335. Bibcode:2005JSPal...3..319E. doi:10.1017/s1477201905001641
- ↑ Conrad, J.L.; Norell, M. (2007). «A complete Late Cretaceous iguanian (Squamata: Reptilia) from the Gobi and identification of a new iguanian clade». American Museum Novitates (3584): 1–47. doi:10.1206/0003-0082(2007)3584[1:ACLCIS]2.0.CO;2. hdl:2246/5877. Consultado em 14 de junho de 2021. Cópia arquivada em 15 de novembro de 2022
- ↑ a b c Conrad, J.L. (2008). «Phylogeny and systematics of Squamata (Reptilia) based on morphology». Bulletin of the American Museum of Natural History. 310: 1–182. doi:10.1206/310.1. hdl:2246/5915
- ↑ Bolet, A.; Evans, S.E. (2010). «A new lizard from the Early Cretaceous of Catalonia (Spain), and the Mesozoic lizards of the Iberian Peninsula». Cretaceous Research. 31 (4): 447–457. Bibcode:2010CrRes..31..447B. doi:10.1016/j.cretres.2010.06.002
- ↑ a b c Bolet, A.; Evans, S.E. (2012). «A tiny lizard (Lepidosauria, Squamata) from the Lower Cretaceous of Spain». Palaeontology. 55 (3): 491–500. Bibcode:2012Palgy..55..491B. doi:10.1111/j.1475-4983.2012.01145.x
- ↑ Tałanda, M. (2018). «An exceptionally preserved Jurassic skink suggests lizard diversification preceded fragmentation of Pangaea». Palaeontology. 61 (5): 659–677. Bibcode:2018Palgy..61..659T. doi:10.1111/pala.12358
- ↑ a b c Simões, T.R.; Caldwell, M.W.; Tałanda, M.; Bernardi, M.; Palci, A.; Vernygora, O.; Bernardini, F.; Mancini, L.; Nydam, R.L. (2018). «The origin of squamates revealed by a Middle Triassic lizard from the Italian Alps». Nature. 557 (7707): 706–709. Bibcode:2018Natur.557..706S. PMID 29849156. doi:10.1038/s41586-018-0093-3
- ↑ Alifanov, V.R. (2019). «Lizards of the Families Eoxantidae, Ardeosauridae, Globauridae, and Paramacellodidae (Scincomorpha) from the Aptian–Albian of Mongolia». Paleontological Journal. 53 (1): 74–88. Bibcode:2019PalJ...53...74A. doi:10.1134/s0031030119010039
- ↑ Russell, A.P. (1986). «The morphological basis of weight-bearing in the scansors of the tokay gecko (Reptilia: Sauria).». Canadian Journal of Zoology. 64 (4): 948–955. Bibcode:1986CaJZ...64..948R. doi:10.1139/z86-144
- ↑ Russell, A.P.; Bauer, A.M.; Laroiya, R. (1997). «Morphological correlates of the secondarily symmetrical pes of gekkotan lizards». Journal of Zoology. 241 (4): 767–790. doi:10.1111/j.1469-7998.1997.tb05747.x
- ↑ Zani, P.A. (2000). «The comparative evolution of lizard claw and toe morphology and clinging performance.». Journal of Evolutionary Biology. 13 (2): 316–325. doi:10.1046/j.1420-9101.2000.00166.x
- ↑ Tulli, M.J.; Cruz, F.B.; Herrel, A.; Vanhooydonck, B.; Abdala, V. (2009). «The interplay between claw morphology and microhabitat use in neotropical iguanian lizards». Zoology. 112 (5): 379–392. Bibcode:2009Zool..112..379T. PMID 19632100. doi:10.1016/j.zool.2009.02.001. hdl:11336/78058
- ↑ Crandell, K.E.; Herrel, A.; Sasa, M.; Losos, J.B.; Autumn, K. (2014). «Stick or grip? Co-evolution of adhesive toepads and claws in Anolis lizards». Zoology. 117 (6): 363–369. Bibcode:2014Zool..117..363C. PMID 25069967. doi:10.1016/j.zool.2014.05.001
- ↑ Kavanagh, K.D.; Shoval, O.; Winslow, B.B.; Alon, U.; Leary, B.P.; Kan, A.; Tabin, C.J. (2013). «Developmental bias in the evolution of phalanges». Proceedings of the National Academy of Sciences. 110 (45): 18190–18195. Bibcode:2013PNAS..11018190K. PMC 3831474. PMID 24151335. doi:10.1073/pnas.1315213110
- ↑ Rothier, P.S.; Brandt, R.; Kohlsdorf, T. (2017). «Ecological associations of autopodial osteology in Neotropical geckos». Journal of Morphology. 278 (3): 290–299. PMID 28112828. doi:10.1002/jmor.20635
- ↑ a b Russell, A.P. (1975). «A contribution to the functional analysis of the foot of the Tokay, Gekko gecko (Reptilia: Gekkonidae)». Journal of Zoology. 176 (4): 437–476. doi:10.1111/j.1469-7998.1975.tb03215.x
- ↑ a b Vanhooydonck, B.; Van Damme, R. (1999). «Evolutionary relationships between body shape and habitat use in lacertid lizards» (PDF). Evolutionary Ecology Research. 1: 785–805. Consultado em 15 de maio de 2020. Cópia arquivada (PDF) em 18 de junho de 2022
- ↑ Melville, J.; Swain, R. (2000). «Evolutionary relationships between morphology, performance and habitat openness in the lizard genus Niveoscincus (Scincidae: Lygosominae)». Biological Journal of the Linnean Society. 70 (4): 667–683. doi:10.1111/j.1095-8312.2000.tb00222.x
- ↑ Schweigert, G. (2007). «Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany – first results and open questions». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 245 (1): 117–125. doi:10.1127/0077-7749/2007/0245-0117
- ↑ Niebuhr, B.; Pürner, T. (2014). «Plattenkalk und Frankendolomit – Lithostratigraphie der Weißjura-Gruppe der Frankenalb (außeralpiner Oberjura, Bayern)». Schriftenreihe der Deutschen Gesellschaft für Geowissenschaften. 83: 5–72. doi:10.1127/sdgg/83/2014/5
- ↑ Rauhut, O.W.M.; Tischlinger, H.; Foth, C. (2019). «A non-archaeopterygid avialan theropod from the Late Jurassic of southern Germany». eLife. 8: e43789. PMC 6516837. PMID 31084702. doi:10.7554/eLife.43789
- ↑ Mönnig, E.; Franz, M.; Schweigert, G. (2018). «Der Jura in der Stratigraphischen Tabelle von Deutschland (STD 2016)» [The Stratigraphic Chart of Germany (STD 2016): Jurassic]. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften. 169 (2): 225–246. doi:10.1127/zdgg/2018/0148
- ↑ a b Munnecke, A.; Westphal, H.; Kölbl-Ebert, M. (2008). «Diagenesis of plattenkalk: examples from the Solnhofen area (Upper Jurassic, southern Germany)». Sedimentology. 55 (6): 1931–1946. Bibcode:2008Sedim..55.1931M. doi:10.1111/j.1365-3091.2008.00975.x
- ↑ a b Wilkin, J. (2020). «The south German Plattenkalks». Geology Today. 36 (1): 27–32. Bibcode:2020GeolT..36...27W. doi:10.1111/gto.12288
- ↑ Viohl, G. (1994). «Fish taphonomy of the Solnhofen plattenkalk — an approach to the reconstruction of the palaeoenvironment». Geobios. 27: 81–90. Bibcode:1994Geobi..27...81V. doi:10.1016/s0016-6995(94)80023-5
- ↑ Seilacher, A.; Reif, W.E.; Westphal, F. (1985). «Sedimentological, ecological and temporal patterns of fossil Lagerstätten». Philosophical Transactions of the Royal Society of London B: Biological Sciences. 311 (1148): 5–24. Bibcode:1985RSPTB.311....5S. doi:10.1098/rstb.1985.0134. Consultado em 22 de janeiro de 2025. Cópia arquivada em 14 de abril de 2024
- ↑ Beardmore, S.R.; Lawlor, E.; Hone, D.W.E. (2017). «Using taphonomy to infer differences in soft tissues between taxa: an example using basal and derived forms of Solnhofen pterosaurs». The Science of Nature. 104 (7–8). 65 páginas. Bibcode:2017SciNa.104...65B. PMID 28721556. doi:10.1007/s00114-017-1486-0
- ↑ Sues, H.-D. (2009). «Archaeopteryx: Der Urvogel von Solnhofen, by Peter Wellnhofer». Journal of Vertebrate Paleontology. 29 (1): 310–311. Bibcode:2009JVPal..29..310S. doi:10.1080/02724634.2009.10010387
- ↑ Bennett, S.C. (1995). «A statistical study of Rhamphorhynchus from the Solnhofen Limestone of Germany: year-classes of a single large species». Journal of Paleontology. 69 (3): 569–580. Bibcode:1995JPal...69..569B. doi:10.1017/S0022336000034946
- ↑ Tennant, J.P.; Mannion, P.D.; Upchurch, P. (2016). «Evolutionary relationships and systematics of Atoposauridae (Crocodylomorpha: Neosuchia): implications for the rise of Eusuchia». Zoological Journal of the Linnean Society. 177 (4): 854–936. doi:10.1111/zoj.12400. hdl:10044/1/29026
- ↑ Ebert, M. (2012). «Crustaceans as prey in fishes of the Solnhofen Archipelago». Archaeopteryx. 30: 1–4
- ↑ Arratia, G. (2016). «New remarkable Late Jurassic teleosts from southern Germany: Ascalaboidae n. fam., its content, morphology, and phylogenetic relationships». Fossil Record. 19 (1): 31–59. Bibcode:2016FossR..19...31A. doi:10.5194/fr-19-31-2016
- ↑ Ebert, M.; Kölbl-Ebert, M. (2010). «Morphology of the leading edges of the caudal and other unpaired fins in the Late Jurassic Aspidorhynchidae and some other actinopterygian fish in light of their function». Archaeopteryx. 28: 1–22
- ↑ Ebert, M. (2014). «The genus Belonostomus Agassiz, 1834 (Neopterygii, Aspidorhynchiformes) in the late Jurassic of the Solnhofen Archipelago, with a focus on Belonostomus kochii Münster, 1836 from Ettling (Germany)». Archaeopteryx. 32: 15–43
- ↑ Lambers, P.H. (1994). «The halecomorph fishes Caturus and Amblysemius in the lithographic limestone of Solnhofen (Tithonian), Bavaria». Geobios. 27: 91–99. Bibcode:1994Geobi..27...91L. doi:10.1016/S0016-6995(94)80024-3. hdl:1874/420973
- ↑ Kriwet, J.; Schmitz, L. (2005). «New insight into the distribution and palaeobiology of the pycnodont fish Gyrodus». Acta Palaeontologica Polonica. 50 (1). Consultado em 17 de maio de 2020. Cópia arquivada em 29 de setembro de 2021
- ↑ Ebert, M.; Lane, J.A.; Kölbl-Ebert, M. (2016). «Palaeomacrosemius thiollieri, gen. et sp. nov., a new Macrosemiidae (Neopterygii) from the Upper Jurassic of the Solnhofen Archipelago (Germany) and Cerin (France), with a revision of the genus Macrosemius». Journal of Vertebrate Paleontology. 36 (5): e1196081. Bibcode:2016JVPal..36E6081E. doi:10.1080/02724634.2016.1196081
- ↑ Ebert, M. (2012). «Histionotus (Actinopterygii, Macrosemiidae) – Eine Gattung mit vielen Fragezeichen» [Histionotus (Actinopterygii, Macrosemiidae) — a genus with many question marks]. Archaeopteryx. 30: 5–15
- ↑ Ebert, M. (2019). «Zandtfuro and Schernfeldfuro, New Genera of Halecomorphi (Actinopterygii) from the Upper Jurassic Solnhofen Archipelago». Journal of Vertebrate Paleontology. 39 (2): e1592759. Bibcode:2019JVPal..39E2759E. doi:10.1080/02724634.2019.1592759
- ↑ Thies, D.; Leidner, A. (2011). «Sharks and guitarfishes (Elasmobranchii) from the Late Jurassic of Europe» (PDF). Palaeodiversity. 4: 63–184
- ↑ Fuchs, D.; Heyng, A.M.; Keupp, H. (2013). «Acanthoteuthis problematica Naef, 1922, an almost forgotten taxon and its role in the interpretation of cephalopod arm armatures». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 269 (3): 241–250. doi:10.1127/0077-7749/2013/0347
- ↑ Engeser, T.; Reitner, J. (1992). «Ein neues Exemplar von Belemnoteuthis mayri Engeser & Reitner, 1981 (Coleoidea, Cephalopoda) aus dem Solnhofener Plattenkalk (Untertithonium) von Wintershof, Bayern» [A new example of Belemnoteuthis mayri Engeser & Reitner, 1981 (Coleoidea, Cephalopoda) from the Solnhofen limestone (Lower Tithonian) from Wintershof, Bavaria] (PDF). Archaeopteryx. 10: 13–17
- ↑ Klug, C.; Fuchs, D.; Schweigert, G.; Röper, M.; Tischlinger, H. (2015). «New anatomical information on arms and fins from exceptionally preserved Plesioteuthis (Coleoidea) from the Late Jurassic of Germany» (PDF). Swiss Journal of Palaeontology. 134 (2): 245–255. Bibcode:2015SwJP..134..245K. doi:10.1007/s13358-015-0093-y
- ↑ Hess, H.; Etter, W. (2011). «Life and death of Saccocoma tenella (Goldfuss)». Swiss Journal of Geosciences. 104 (1): 99–106. Bibcode:2011SwJG..104...99H. doi:10.1007/s00015-011-0059-z
- ↑ Winkler, N. (2017). «Two new penaeid shrimps (Crustacea: Decapoda: Dendrobranchiata) from the Solnhofen lithographic limestones (Upper Jurassic, southern Germany)». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 283 (1): 9–24. doi:10.1127/njgpa/2017/0623
- ↑ Lomax, D.R.; Racay, C.A. (2012). «A Long Mortichnial Trackway of Mesolimulus walchi from the Upper Jurassic Solnhofen Lithographic Limestone near Wintershof, Germany». Ichnos. 19 (3): 175–183. Bibcode:2012Ichno..19..175L. doi:10.1080/10420940.2012.702704. Consultado em 14 de junho de 2021. Cópia arquivada em 29 de agosto de 2024
- ↑ Göhlich, U.B. (2017). «Catalogue of the fossil bird holdings of the Bavarian State Collection of Palaeontology and Geology in Munich» (PDF). Zitteliana. 89: 331–349. Consultado em 17 de maio de 2020. Cópia arquivada (PDF) em 18 de fevereiro de 2020
- ↑ Vidovic, S.U.; Martill, D.M. (2017). «The taxonomy and phylogeny of Diopecephalus kochi (Wagner, 1837) and "Germanodactylus rhamphastinus" (Wagner, 1851)» (PDF). Geological Society, London, Special Publications. 455 (1): 125–147. doi:10.1144/sp455.12. Consultado em 6 de junho de 2020. Cópia arquivada (PDF) em 27 de abril de 2019
- ↑ a b c Carannante, G.; Signore, M.; Vigorito, M. (2006). «Vertebrate-rich Plattenkalk of Pietraroia (Lower Cretaceous, Southern Apennines, Italy): a new model». Facies. 52 (4): 555–577. Bibcode:2006Faci...52..555C. doi:10.1007/s10347-006-0075-z
- ↑ a b c Bravi, S.; Garassino, A (1998). «New biostratographic and palaeoecologic observations on the "Plattenkalk" of the lower Cretaceous (Albian) of Pietraroia (Benevento, S−Italy) and its decapod crustaceans assemblage». Atti Societá Italiana Scienze Naturali, Milano. 138: 119–171
- ↑ Evans, S.E.; Raia, P.; Barbera, C. (2006). «The Lower Cretaceous lizard genus Chometokadmon from Italy» (PDF). Cretaceous Research. 27 (5): 673–683. Bibcode:2006CrRes..27..673E. doi:10.1016/j.cretres.2006.03.004. Consultado em 22 de janeiro de 2025. Cópia arquivada (PDF) em 10 de julho de 2024
- ↑ Cau, A.; Baiano, M.A.; Raia, P. (2014). «A new sphenodontian (Reptilia, Lepidosauria) from the Lower Cretaceous of Southern Italy and the phylogenetic affinities of the Pietraroia Plattenkalk rhynchocephalians». Cretaceous Research. 49: 172–180. Bibcode:2014CrRes..49..172C. doi:10.1016/j.cretres.2014.02.001
- ↑ Dal Sasso, C.; Signore, M. (1998). «Exceptional soft-tissue preservation in a theropod dinosaur from Italy» (PDF). Nature. 392 (6674): 383–387. Bibcode:1998Natur.392..383D. doi:10.1038/32884. Consultado em 22 de janeiro de 2025. Cópia arquivada (PDF) em 20 de setembro de 2016
- ↑ Buscalioni, A.D.; Piras, P.; Vullo, R.; Signore, M.; Barbera, C. (2011). «Early eusuchia crocodylomorpha from the vertebrate-rich Plattenkalk of Pietraroia (Lower Albian, southern Apennines, Italy)». Zoological Journal of the Linnean Society. 163 (S1): S199–S227. doi:10.1111/j.1096-3642.2011.00718.x
- ↑ McGowan, G.; Evans, S.E. (1995). «Albanerpetontid amphibians from the Cretaceous of Spain». Nature. 373 (6510): 143–145. Bibcode:1995Natur.373..143M. doi:10.1038/373143a0
- ↑ Taverne, L.; Capasso, L.; Del Re, M. (2020). «Osteology and phylogenetic relationships of Gregoriopycnodus bassanii gen. nov., a pycnodont fish (Pycnodontidae) from the marine Albian (Lower Cretaceous) of Pietraroja (southern Italy)» (PDF). Geo Eco Trop. 44 (1): 161–174. Consultado em 17 de maio de 2020. Cópia arquivada (PDF) em 23 de fevereiro de 2020
- ↑ a b Dalla Vecchia, F.; Barbera, C.; Bizzarini, F.; Bravi, S.; Delfino, M.; Giusberti, L.; Guidotti, G.; Mietto, P.; Palazzoni, C.; Roghi, G.; Signore, M.; Simone, O. (2005). «Il Cretaceo Marino» [The Marine Cretaceous]. In: Bonfiglio, L. Paleontologia dei Vertebrati in Italia: Evoluzione Biologica, Significato Ambientale e Paleogeografico [Vertebrate Paleontology in Italy: Biological Evolution, Environmental and Paleogeographic Significance]. Col: Sezione Scienze della Terra. 6. [S.l.]: Memorie del Museo Civico di Storia Naturale di Verona. pp. 101–112
- ↑ Taverne, L.; Capasso, L. (2019). «Osteology of Cavinichthys pachylepis gen. and sp. nov. (Teleostei, Crossognathiformes, Pachyrhizodontidae) from Pietraroja (Lower Cretaceous, Campania, south Italy)». Thalassia Salentina. 41: 53–70. doi:10.1285/i15910725v41p53
- ↑ Taverne, L.; Capasso, L. (2017). «Osteology and relationships of Caeus ("Chanos") leopoldi (Teleostei, Gonorynchiformes, Chanidae) from the marine Albian (Early Cretaceous) of Pietraroja (Campania, southern Italy)». Bollettino dem Museo Civico di Storia Naturale di Verona, Geologia Paleontologia Preistoria. 41: 3–20
- ↑ Taverne, L.P.; Capasso, L. (2016). «Revision of Ionoscopus petrarojae (Ionoscopiformes, Osteichthyes) from the Albian of Pietraroja (Campania, southern Italy)». Thalassia Salentina. 38: 65–80. doi:10.1285/i15910725v38p65
- ↑ Taverne, L.; Capasso, L. (2017). «Italophiopsis derasmoi gen. and sp. nov. (Ionoscopiformes, Italophiopsidae fam. nov.) from the Cretaceous of Pietraroja (Italy)». Thalassia Salentina. 39: 9–24. doi:10.1285/i15910725v39p9