Xiphodon
Xiphodon
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Eoceno Médio – Oligoceno Inferior 40–33,4 Ma | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Espécie-tipo | |||||||||||||
| † Anoplotherium gracile (= †Xiphodon gracilis) Cuvier, 1822 | |||||||||||||
| Outras espécies | |||||||||||||
| Sinónimos | |||||||||||||
Sinônimos de X. gracilis
Espécies duvidosas
| |||||||||||||

Xiphodon é o gênero-tipo [en] da extinta família de artiodáctilos do Paleogeno, Xiphodontidae. Ele, como outros xifodontídeos, era endêmico da Europa Ocidental e viveu desde o Eoceno Médio até o início do Oligoceno. Fósseis de Montmartre, em Paris, França, que pertenciam a Xiphodon gracilis, foram descritos pela primeira vez pelo naturalista francês Georges Cuvier em 1804. Embora ele tenha atribuído a espécie a Anoplotherium, ele reconheceu que ela diferia de Anoplotherium commune pela sua dentição e ossos dos membros, movendo-a mais tarde para seu próprio subgênero em 1822. Xiphodon foi promovido ao status de gênero por outros naturalistas nas décadas seguintes. Hoje, é definido pela espécie-tipo Xiphodon gracilis e duas outras espécies, Xiphodon castrensis e Xiphodon intermedium.
Literalmente significando "dente de espada" em grego antigo, Xiphodon possuía uma dentição selenodonte especializada em forma de lâmina, com seus incisivos, caninos e pré-molares braquidontes (de coroa baixa) tendo bordas afiadas para cortar vegetação mais alta, como folhas e arbustos. Ele também retinha molares primitivos em comparação com seu parente Dichodon [en], indicando especializações dietéticas diferentes. Xiphodon também é o único xifodontídeo conhecido a partir de fósseis pós-cranianos. Sua morfologia craniana, combinada com membros delgados e alongados, sugere comportamentos semelhantes aos dos camelídeos do Paleogeno norte-americano, como Poebrotherium [en], incluindo adaptações para corrida. No entanto, a extensão total de seu comportamento e relações evolutivas permanece incerta, e suas semelhanças com os camelídeos são provavelmente um caso de evolução convergente.
Xiphodon viveu na Europa Ocidental quando esta era um arquipélago isolado do resto da Eurásia, o que significa que viveu em um ambiente com várias outras faunas endêmicas. O xifodonte fez sua primeira aparição no Eoceno Médio, pouco antes de uma mudança para condições mais secas, mas ainda subúmidas, que levaram a plantas cada vez mais abrasivas. As espécies de Xiphodon eram relativamente pequenas, com a segunda espécie a aparecer, Xiphodon intermedium, tendo um peso estimado de 4,6 kg. Xiphodon gracilis foi a última e maior espécie dentro do gênero, em uma tendência evolutiva de aumento de tamanho.
Ele e outros gêneros de xifodontes foram extintos no evento de extinção/mudança faunística da Grande Coupure, coincidindo com mudanças para maior glaciação e sazonalidade, além de dispersões de faunas imigrantes asiáticas para a Europa Ocidental. As causas de sua extinção são atribuídas a interações negativas com as faunas imigrantes (competição por recursos, predação), mudanças ambientais decorrentes das alterações climáticas, ou alguma combinação dos dois.
Taxonomia
História da pesquisa
Início da história


Em 1804, o naturalista francês Georges Cuvier estabeleceu várias espécies fósseis como pertencentes ao gênero Anoplotherium, além de Anoplotherium commune. Uma das espécies que ele nomeou foi Anoplotherium medium, que ele disse ter pés delgados, alongados e didáctilos (com dois dedos). Ele pensava que Anoplotherium tinha cascos didáctilos em vez de tridáctilos (com três dedos), o que o teria separado do outro "paquiderme" Palaeotherium [en]. Com base nos cascos e na dentição, ele concluiu que Anoplotherium era semelhante a ruminantes ou camelídeos.[1][2] Em 1807, Cuvier elaborou mais suas ideias sobre os ossos dos membros, sugerindo que se assemelhavam superficialmente aos das lhamas. Ele explicou que a terceira falange de Anoplotherium medium diferia das das lhamas por suas proporções ligeiramente maiores. Ele argumentou que, como sua terceira falange se assemelhava mais às dos ruminantes, estava mais proximamente relacionado a esse grupo de mamíferos do que Anoplotherium commune estava a eles. Cuvier também disse que outras morfologias pós-cranianas da cabeça do fêmur [en] e da tíbia se assemelhavam mais às dos ruminantes do que às dos camelos.[3] Ele atribuiu vértebras lombares danificadas a Anoplotherium medium em 1808.[4]

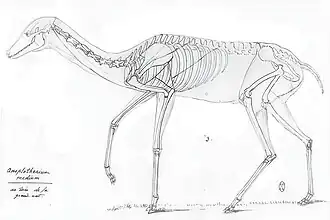
Cuvier publicou seus desenhos de reconstruções esqueléticas de duas espécies de Anoplotherium em 1812, com base em restos fósseis conhecidos, incluindo Anoplotherium medium. Ele observou que não tinha evidências de ossos do tronco ou da cauda de A. medium, mas que tinha fósseis de seu crânio, pescoço, tíbia e osso do tarso, somando-se à evidência do pé traseiro que ele descreveu anos antes. Ele afirmou que, em contraste com o mais robusto Anoplotherium commune, Anoplotherium medium era mais grácil na forma e, portanto, teria sido construído para a cursorialidade, semelhante a ungulados existentes, como gazelas ou corças. Ele hipotetizou, portanto, que, ao contrário de Anoplotherium commune, que ele pensava ter hábitos semiaquáticos, Anoplotherium medium não poderia ter vivido em pântanos ou lagoas. Em vez disso, teria pastado em ervas e arbustos em terras secas e teria comportamentos mais "tímidos", não muito diferentes dos ruminantes gráceis. Cuvier também propôs que provavelmente não tinha uma cauda longa como Anoplotherium commune e que tinha orelhas móveis como os cervos para ouvir o perigo com antecedência. Anoplotherium medium, segundo o naturalista, tinha pelo curto e provavelmente não ruminava.[5][6]
Em 1822, Cuvier estabeleceu o subgênero Xiphodon para o gênero Anoplotherium e mudou o nome da espécie Anoplotherium medium para Xiphodon gracile porque sentiu que era um nome de espécie mais adequado. Ele argumentou que a espécie tem uma cabeça com a forma e o tamanho aproximados da "corinne" (um termo arcaico para a gazela-dorcas), com focinhos afiados, e difere de Anoplotherium commune com base em molares longos e afiados. No entanto, ele também sugeriu que as duas espécies não diferem no nível de gênero.[7] Juntamente com outras espécies fósseis da bacia de Paris [en], foi retratado em desenhos de 1822 pelo paleontólogo francês Charles Léopold Laurillard, sob a direção de Cuvier, embora as restaurações não fossem tão detalhadas quanto as de Cuvier.[8] O nome do gênero Xiphodon significa "dente de espada" e é um composto das palavras gregas antigas ξίφος (xiphos, 'espada') e ὀδούς (odoús, 'dente').[9]
.jpg)
Em 1848, o naturalista francês Paul Gervais afirmou que Xiphodon era um gênero distinto de Anoplotherium. Ele transmitiu de forma semelhante que Xiphodon gracile era esbelto como os antílopes, mas ligeiramente menor que as gazelas-dorcas. Ele erigiu a segunda espécie, Xiphodon gelyense, da comuna francesa de Saint-Gély-du-Fesc.[10] Ele também reclassificou Hyopotamus (= Bothriodon [en]) crispus para Xiphodon.[11] A validade de Xiphodon como gênero também foi apoiada pelo naturalista britânico Richard Owen no mesmo ano, que também erigiu Dichodon [en].[12] Owen emendou as espécies Xiphodon gracile e Xiphodon gelyense para Xiphodon gracilis e Xiphodon Gelyensis, respectivamente, em 1857.[13]
Xiphodon gracilis estava entre os táxons fósseis retratados no conjunto dos Dinossauros do Crystal Palace [en], no Parque Crystal Palace [en], no Reino Unido, aberto ao público desde 1854 e construído pelo escultor inglês Benjamin Waterhouse Hawkins. Benjamin aparentemente se recusou a reconhecer o nome do gênero ou não o conhecia, o que significa que as esculturas da espécie foram referidas como "A. gracile". As esculturas existentes de Anoplotherium commune foram historicamente confundidas com "A. gracile", resultado de ambas as espécies terem sido listadas nos primeiros guias do Crystal Palace. Uma ilustração da oficina de Hawkins revela que quatro esculturas representando "A. gracile" foram construídas por ele, três das quais desapareceram sem deixar vestígios.[14]
A quarta escultura foi confundida com um filhote de Megaloceros giganteus (alce-gigante) e foi associada às esculturas de Megaloceros por um período de tempo desconhecido. A única escultura sobrevivente mede 1,7 m de comprimento do focinho à cauda e tem uma aparência semelhante à de uma lhama, dado seu pescoço longo, cabeça pequena, olhos grandes, corpo robusto, nariz de camelo, lábios ramificados e um focinho estreito. A aparência geral da escultura corresponde à descrição anatômica da espécie feita por Cuvier, sendo a principal imprecisão a reconstrução de pequenos dedos adicionais, semelhantes aos de Anoplotherium commune. Seu design e a intenção de representá-lo como um rebanho foram provavelmente inspirados na aparência e nos comportamentos das lhamas sul-americanas. A ilustração da oficina de Hawkins implica que as esculturas de Xiphodon gracilis foram concebidas para representar um rebanho relaxado.[14]
Espécies e sinônimos adicionais
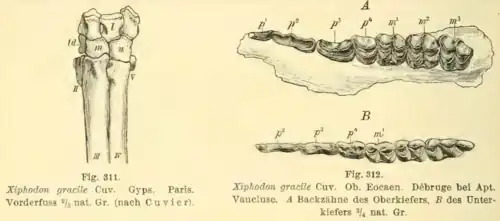
Em 1873, Vladimir Kovalevsky [en] rejeitou a reclassificação de Gervais de Hyopotamus crispus (= Elomeryx crispus) para Xiphodon.[15] Em 1876, o naturalista britânico William Henry Flower expressou incerteza sobre se Dichodon era suficientemente distinto de Xiphodon. Como não gostava do conceito de ter múltiplos gêneros proximamente relacionados, ele optou por colocar em Xiphodon a espécie recém-erigida Xiphodon platyceps.[16] No mesmo ano, Kovalevsky erigiu uma espécie menor recém-determinada que ele nomeou Xiphodon castrense em homenagem à comuna francesa de Castres. Ele também afirmou que seus pré-molares afiados justificavam a etimologia do gênero "dente de espada".[17] Gervais erigiu outra espécie que ele atribuiu provisoriamente a Xiphodon no mesmo ano, nomeando-a X? tragulinum.[18] Em 1884, o naturalista francês Henri Filhol [en] erigiu a espécie Xiphodon magnum com base em um fóssil de mandíbula inferior, argumentando que a espécie era maior que Xiphodon gracilis.[19]
O naturalista britânico Richard Lydekker revisou as espécies conhecidas de Dichodon e Xiphodon em 1885, confirmando que ambos são gêneros distintos. Ele também reafirmou a validade de Xiphodon gracilis e Xiphodon gelyensis, e então sinonimizou Xiphodontherium, erigido anteriormente por Filhol em 1877, com Xiphodon, reclassificando assim Xiphodontherium secundarius para Xiphodon. Ele também sugeriu que Xiphodon platyceps pode ser sinônimo de Dacrytherium [en] ovinum. Ele não fez referência a Xiphodon castrense em seu catálogo.[20] Em 1886, o paleontólogo alemão Max Schlosser [en] transferiu "X. gelyense" para o gênero mais novo Phaneromeryx.[21]
Em 1910, o paleontólogo suíço Hans Georg Stehlin [en] sinonimizou Xiphodontherium com Amphimeryx [en], tornando também Xiphodontherium primaevum e Xiphodontherium secundarium sinônimos de Amphimeryx murinus no processo. Ele afirmou que X. platyceps era muito provavelmente sinônimo de Dichodon cuspidatum, considerou X? tragulinum um nome duvidoso e expressou dúvida de que X. magnum, se válido, realmente pertencesse a Xiphodon. Ele também criou a espécie X. intermedium com base em medidas dentárias intermediárias entre o menor X. castrense e o maior X. gracile.[22]
Em 2000, Jerry J. Hooker e Marc Weidmann listaram X. castrensis como um nome emendado para X. castrense.[23] Segundo Jörg Erfurt e Grégoire Métais em 2007, X. castrensis e X. intermedium não possuem diagnósticos diferenciais definidos além dos tamanhos dentários.[24]
Classificação

Xiphodon é o gênero-tipo [en] da Xiphodontidae, uma família de artiodáctilos do Paleogeno endêmica da Europa Ocidental que viveu do Eoceno Médio ao Oligoceno Inferior (~44 Ma a 33 Ma). Como as outras famílias de artiodáctilos endêmicas contemporâneas da Europa Ocidental, as origens evolutivas da Xiphodontidae são pouco conhecidas. Embora se pensasse que Xiphodon tivesse aparecido já em MP10 das zonas do Paleogeno de mamíferos [en] com base em uma localidade, essa atribuição baseia-se em material fóssil muito pobre.[24] Em vez disso, geralmente se pensa que a Xiphodontidae apareceu pela primeira vez em MP14, tornando-os os primeiros representantes de artiodáctilos com dentição selenodonte a aparecer na massa de terra, juntamente com Amphimerycidae [en].[25] Mais especificamente, os primeiros representantes de xifodontes a aparecer foram os gêneros Dichodon e Haplomeryx [en].[26] Dichodon e Haplomeryx continuaram a persistir no Eoceno Superior, enquanto Xiphodon fez sua primeira aparição em MP16. Outro xifodonte, Paraxiphodon [en], é conhecido por ter ocorrido apenas em localidades de MP17a.[26] Os três primeiros gêneros viveram até o Oligoceno Inferior, onde foram registrados como extintos como resultado do evento de mudança faunística da Grande Coupure.[27]
As relações filogenéticas da Xiphodontidae, bem como da Anoplotheriidae, Mixtotheriidae [en] e Cainotheriidae, têm sido elusivas devido às morfologias selenodontes (ou com cristas em forma de crescente) dos molares, que eram convergentes com tilópodes ou ruminantes.[28] Alguns pesquisadores consideraram as famílias selenodontes Anoplotheriidae, Xiphodontidae e Cainotheriidae como estando dentro de Tylopoda, devido a características pós-cranianas que eram semelhantes aos tilópodes da América do Norte no Paleogeno.[29] Outros pesquisadores as vinculam como sendo mais proximamente relacionadas a ruminantes do que a tilópodes, com base na morfologia dentária. Diferentes análises filogenéticas produziram resultados diferentes para as famílias de artiodáctilos europeus do Eoceno com selenodontia "derivada" (ou com novas características evolutivas), tornando incerto se elas estavam mais próximas de Tylopoda ou Ruminantia.[30][31] Possivelmente, Xiphodontidae pode ter surgido de um grupo dicobunoide desconhecido, tornando assim sua semelhança com os tilópodes um caso de evolução convergente.[24]
Em um artigo publicado em 2019, Romain Weppe et al. conduziram uma análise filogenética sobre Cainotherioidea dentro de Artiodactyla, baseada em características mandibulares e dentárias, especificamente em termos de relações com artiodáctilos do Paleogeno. Os resultados recuperaram que a superfamília estava proximamente relacionada com Mixtotheriidae e Anoplotheriidae. Eles determinaram que Cainotheriidae, Robiacinidae, Anoplotheriidae e Mixtotheriidae formavam um clado que era o grupo irmão de Ruminantia, enquanto Tylopoda, juntamente com Amphimerycidae e Xiphodontidae, se separaram mais cedo na árvore.[31] A árvore filogenética publicada no artigo e em outro trabalho sobre os cainoterioides está delineada abaixo:[32]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Em 2020, Vincent Luccisano et al. criaram uma árvore filogenética dos artiodáctilos basais, a maioria endêmica da Europa Ocidental, do Paleogeno. Em um clado, os "bunoselenodontes endêmicos europeus" Mixtotheriidae, Anoplotheriidae, Xiphodontidae, Amphimerycidae, Cainotheriidae e Robiacinidae são agrupados com Ruminantia. A árvore filogenética produzida pelos autores é mostrada abaixo:[30]
| Artiodactyla |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Em 2022, Weppe criou uma análise filogenética em sua tese acadêmica sobre as linhagens de artiodáctilos do Paleogeno, focando mais especificamente nas famílias endêmicas europeias. Ele afirmou que sua filogenia foi a primeira formal a propor afinidades entre Xiphodontidae e Anoplotheriidae. Ele descobriu que Anoplotheriidae, Mixtotheriidae e Cainotherioidea formam um clado com base em características dentárias sinapomórficas (características que se acredita terem se originado de seu ancestral comum mais recente). O resultado, mencionou Weppe, corresponde a análises filogenéticas anteriores sobre Cainotherioidea com outros artiodáctilos endêmicos do Paleogeno europeu que apoiam as famílias como um clado. Como resultado, ele argumentou que a superfamília proposta Anoplotherioidea, composta pelas Anoplotheriidae e Xiphodontidae, como proposto por Alan W. Gentry e Hooker em 1988, é inválida devido à polifilia das linhagens na análise filogenética. No entanto, Xiphodontidae ainda foi encontrado como parte de um clado mais amplo com os outros três grupos. Dentro de Xiphodontidae, a árvore filogenética de Weppe classificou Haplomeryx como um táxon irmão do clado que consiste em Xiphodon com Dichodon.[28]
Descrição
Crânio

Xiphodon é diagnosticado por ter um crânio alongado que é convexo na área superior que leva à órbita. As próprias órbitas são bem abertas em suas áreas posteriores. O focinho é alongado e tem uma aparência arredondada. No gênero xifodonte também existem grandes partes timpânicas do osso temporal [en] e ossos perióticos visíveis. Os forames palatinos [en] maiores são extensos em tamanho, dos dentes I3 a P1.[33][24] A mandíbula parece ser baixa horizontalmente, dando um contorno retilíneo.[34]
A maxila constitui a maior parte das áreas laterais do crânio, enquanto a pré-maxila se estende até os processos alveolares. Os ossos nasais são estreitos e alongados, suas passagens mal se estendem sobre as aberturas das narinas externas e formam com elas uma estreita faixa óssea. Na visão posterior, o focinho parece ter um contorno em forma de U. O focinho de Xiphodon é semelhante ao de Dichodon, mas difere dele por seu alongamento mais aparência arredondada e as maxilas que constituem parte do focinho sendo menos extensas em altura. O focinho de Dichodon, em comparação, é mais curto e mais estreito.[35]
O palato duro da boca superior parece côncavo e tem uma sutura [en] pré-maxilar-maxilar visível que se estende da borda externa da mandíbula para trás. Ambos os tipos de forame palatino de Xiphodon têm proporções e posições semelhantes ao forame palatino de Dichodon, mas os de Xiphodon são maiores em comprimento e têm morfologias diferentes das de Dichodon.[35]
Além das grandes e ocas bulas timpânicas, o canal auditivo tem bordas elevadas e se abre em uma posição inclinada, ligeiramente à frente da sutura do osso occipital. O osso esquamosal forma um componente principal da abóbada craniana de Xiphodon. Uma crista acima da área externa do canal auditivo se estende até a borda convexa superior do arco zigomático. A morfologia do canal auditivo superior em Xiphodon é semelhante à do camelídeo do Paleogeno Poebrotherium [en]. A área posterior dos arcos zigomáticos é estreita e próxima à abóbada craniana. A fossa mandibular [en] parece plana e horizontal, com um pequeno processo pós-glenoide (ou projeção) em forma de colher.[35]
Anatomia do endocasto
Um endocasto parcial de Xiphodon gracilis do Museu Nacional de História Natural, na França, foi observado pela primeira vez por Colette Dechaseaux em 1963, que tinha um neocórtex visível. O sulco [en] suprassilviano (ou suprassílvio) tem uma posição alta dentro do neocórtex, mas pode ter tido uma posição ainda mais alta dentro do cérebro. O sulco lateral é longo e distinto, e um giro à sua frente parece ter sido elevado. O sulco entolateral não parece extenso em comprimento. O giro entre o sulco lateral e o sulco entolateral é estreito em comparação com o que fica entre o sulco lateral e o suprassílvio. Todos os três sulcos são distintamente profundos em elevação dentro do neocórtex, dando-lhe uma aparência de colina. O neocórtex tem uma aparência semelhante à dos tilópodes do Paleogeno, como Poebrotherium.[36]
Dechaseaux descobriu mais tarde um grande flóculo [en] esférico do cérebro do mesmo endocasto em 1967. O flóculo está separado do hemisfério cerebelar e ocupa espaço dentro da parte petrosa do osso temporal [en], dentro do osso periótico da orelha. Ele também apresenta uma aparência fechada em suas bordas externas.[35]
Dentição

Tanto Xiphodon quanto Dichodon exibem conjuntos completos de 3 incisivos, 1 canino, 4 pré-molares e 3 molares em cada metade das mandíbulas superior e inferior,[35][37] consistentes com a fórmula dentária primitiva de mamíferos placentários de para um total de 44 dentes.[38] Como membros de Xiphodontidae, eles compartilham tanto incisivos pequenos quanto a ausência de diastemas distintos.[34] Eles também são caracterizados por caninos indistintos em comparação com outros dentes e pré-molares alongados. Os xifodontídeos adicionalmente possuem dentes P4 molariformes, molares superiores com 4 a 5 cúspides em forma de crescente e molares inferiores selenodontes com 4 cristas, cúspides linguais comprimidas e cúspides labiais em forma de crescente.[24]

A dentição de Xiphodon é braquidonte.[35] Seus pré-molares são alongados e não especializados, enquanto seus molares superiores são de formato quadrangular, exibem ectolofos em forma de W e mostram aumentos de tamanho de M1 a M3. Eles exibem cinco cúspides, quatro das quais são em forma de crescente. As cúspides do paracônulo e do metacônulo se conectam às cúspides do parastilo e do metastilo, respectivamente. A cúspide do protocone é mais isolada de outras cúspides e tem uma crista preprotocrista curta.[24][33]
Os terceiros incisivos se assemelham a caninos, mas se projetam ligeiramente para a frente e são separados dos caninos por pequenos diastemas. Os dois primeiros outros incisivos não são conhecidos, mas, com base em seus alvéolos redondos, eles se projetariam ligeiramente para a frente, assim como os terceiros incisivos. O canino de Xiphodon é premolariforme, com sua afiação semelhante à dos pré-molares, mas difere deles pelo diâmetro mesiodistal menor e pela assimetria. Todos os três pré-molares dianteiros parecem comprimidos no lado labiolingual dos dentes, sendo o segundo pré-molar o mais alongado dos três. Eles parecem mais afiados quanto mais próximos estão do canino, com o primeiro pré-molar parecendo ser o mais afiado como resultado. As semelhanças dos terceiros incisivos, caninos e pré-molares de Xiphodon revelam que o artiodáctilo tinha uma dentição especializada em forma de lâmina.[35]
Jean Sudre, em 1978, argumentou que Xiphodon exibia a tendência evolutiva dos molares se tornarem mais quadrangulares e que suas formas selenodontes já estavam presentes na espécie mais basal, X. castrensis.[33]
Esqueleto pós-craniano
Xiphodon é o único membro de sua família para o qual se conhecem evidências pós-cranianas, representadas principalmente pelas pedreiras de gesso de Montmartre, no caso de Xiphodon gracilis, como descrito anteriormente por Cuvier.[24] As vértebras cervicais, representadas pelo áxis e duas vértebras seguintes, atingem quase 70% do comprimento total do crânio, indicando um pescoço longo.[35]
Os membros anteriores do xifodonte são finos e alongados.[24] O rádio e a ulna são mais alongados que o úmero. Os pés de Xiphodon têm dois dígitos proeminentes: o dígito III e o dígito IV. Os dígitos laterais II e V são muito reduzidos.[35] Como resultado, Xiphodon é um gênero didáctilo, ou de dois dedos. Seus metapodiais laterais são reduzidos, enquanto o osso cuboide e o osso navicular não são fundidos.[24][33] As pernas longas podem ter suportado um corpo suspenso. As características pós-cranianas de Xiphodon são consideradas semelhantes às dos camelídeos do Paleogeno, como Poebrotherium, embora não esteja claro se Xiphodon é mais primitivo ou mais derivado em relação aos tilópodes norte-americanos.[24] O astrágalo originalmente atribuído a Xiphodon é estreito e alongado, com seu sulco tibial parecendo estreito, mas profundo. A faceta calcaneana posterior, que ocupa uma porção significativa da face posterior do astrágalo, é larga em comparação com a de Dacrytherium. O calcâneo parece semelhante ao de Dacrytherium, mas difere por uma tuberosidade posterior mais alongada.[39] No entanto, os fósseis pós-cranianos foram posteriormente reatribuídos a Leptotheridium [en], enquanto o astrágalo originalmente atribuído a Dichodon foi reclassificado para Xiphodon.[23] O astrágalo reatribuído a Xiphodon foi descrito como sendo estreito e alongado e tendo um sulco tibial profundo.[39]
Tamanho
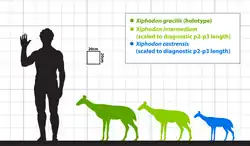
Xiphodontidae é caracterizado por suas espécies serem de tamanho muito pequeno a médio. Xifodontes com muitas espécies, como Dichodon e Haplomeryx, tendiam a exibir aumentos evolutivos de tamanho.[24] As espécies pertencentes a Xiphodon são diagnosticadas como artiodáctilos de tamanho médio. A espécie basal Xiphodon castrensis é a menor do gênero, seguida por Xiphodon intermedium com medidas dentárias ligeiramente maiores. A espécie mais tardia, Xiphodon gracilis, era a maior das três. Sudre apontou que as tendências de tamanho indicam aumentos evolutivos.[33]
A massa corporal estimada de Xiphodon intermedium foi calculada por Helder Gomes Rodrigues et al. em 2019 com base em um astrágalo da Universidade de Lyon que media 17 mm de comprimento e 8 mm de largura, resultando em 4,6 kg. A fórmula de massa corporal baseada em astrágalos foi estabelecida anteriormente por Jean-Noël Martinez e Sudre em 1995 para artiodáctilos do Paleogeno, embora Xiphodon não tenha sido incluído no estudo inicial.[40][41]
Paleobiologia

Xiphodontidae é um grupo de artiodáctilos selenodontes da Europa Ocidental, o que significa que a família era provavelmente adaptada a hábitos alimentares folívoros (comedores de folhas).[42] Este foi especialmente o caso de Xiphodon, que exibia uma dentição especializada para se alimentar de folhas, brotos de árvores e arbustos. Xiphodon reteve a característica primitiva de ter molares com cinco cúspides e mudou para uma dentição de corte, enquanto Dichodon tinha pré-molares progressivamente molarizados para a função de moer alimentos, o que significa que os dois gêneros tinham diferentes tipos de especializações ecológicas. Dechaseaux considerou que os dois gêneros de xifodontídeos podem ter sido mais derivados do que os tilópodes do Paleogeno da América do Norte.[35]
Os membros anteriores de Xiphodon parecem ser semelhantes aos dos camelídeos do Paleogeno, que tinham adaptações para a cursorialidade. Devido às semelhanças dentárias e pós-cranianas, Xiphodon poderia ter sido um equivalente ecológico europeu. No entanto, não se pode provar se Xiphodon tinha uma locomoção de passo como os camelídeos. Devido à falta de evidências pós-cranianas de outros xifodontes, não é possível provar que suas morfologias pós-cranianas sejam semelhantes às de Xiphodon.[24][35]
Paleoecologia
Eoceno Médio
Durante a maior parte do Eoceno, prevaleceu um clima de estufa com ambientes húmidos e tropicais, com precipitações consistentemente altas. As ordens de mamíferos modernos, incluindo Perissodactyla, Artiodactyla e Primatas (ou a subordem Euprimates), já haviam aparecido no Eoceno Inferior, diversificando-se rapidamente e desenvolvendo dentições especializadas para a folivoria. As formas onívoras, na sua maioria, ou mudaram para dietas folívoras ou extinguiram-se no Eoceno Médio (47-37 milhões de anos atrás), juntamente com os "condilartros" arcaicos. No Eoceno Superior (aprox. 37-33 Ma), a maioria das dentições de formas unguladas mudou de cúspides bunodontes (ou arredondadas) para cristas cortantes (ou seja, lofos) para dietas folívoras.[43][44]
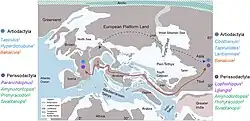
As conexões terrestres entre a Europa Ocidental e a América do Norte foram interrompidas por volta de 53 Ma. Desde o Eoceno Inferior até o evento de extinção Grande Coupure (56-33,9 Ma), a Eurásia Ocidental estava separada em três massas de terra: Europa Ocidental (um arquipélago), Balcanatolia (entre o Mar de Paratethys ao norte e o Oceano Neotethys ao sul) e Eurásia Oriental.[45] As faunas de mamíferos holárticos da Europa Ocidental estavam, portanto, em grande parte isoladas de outras massas de terra, incluindo a Gronelândia, África e Eurásia Oriental, permitindo o desenvolvimento de endemismo.[44] Portanto, os mamíferos europeus do Eoceno Superior (MP17-MP20 das zonas do Paleogeno de mamíferos) eram, na sua maioria, descendentes de grupos endêmicos do Eoceno Médio.[46]

Xiphodon fez sua primeira aparição conhecida em MP16 com base na localidade de Robiac, na França, como X. castrensis. A espécie está restrita a localidades de MP16.[47] Naquela época, teria coexistido com perissodáctilos (Palaeotheriidae, Lophiodontidae [en] e Hyrachyidae), artiodáctilos não endêmicos (Dichobunidae e Tapirulidae), artiodáctilos endêmicos europeus (Choeropotamidae [en] (possivelmente polifilético, no entanto), Cebochoeridae, Mixtotheriidae, Anoplotheriidae, Amphimerycidae e outros membros da Xiphodontidae) e primatas (Adapidae, Omomyidae).[42][25][48] Também coocorreu com metatérios (Herpetotheriidae [en]), roedores (Ischyromyidae, Theridomyoidea, Gliridae), eulipotiflanos, morcegos, apatotérios [en], carnivoraformes (Miacidae) e hienodontes [en] (Hyainailourinae [en], Proviverrinae [en]).[26]
Dentro de Robiac, fósseis de X. castrensis foram encontrados com os de outros mamíferos como os herpetoteriídeos Peratherium [en] e Amphiperatherium [en], o apatemídeo Heterohyus [en], o nictitério Saturninia, roedores (Glamys, Elfomys, Plesiarctomys, Remys), os omomiídeos Pseudoloris e Necrolemur [en], o adapídeo Adapis [en], os hienodontes Paroxyaena e Cynohyaenodon [en], os carnivoraformes Paramiacis [en] e Simamphicyon, paleotérios (Palaeotherium, Plagiolophus [en], Anchilophus, Leptolophus), o lofiodonte Lophiodon [en], o hiraquiídeo Chasmotherium, os ceboquerídeos Acotherulum e Cebochoerus [en], o queropotamídeo Choeropotamus [en], o tapirulídeo Tapirulus, os anoploteriídeos Dacrytherium e Catodontherium [en], o dicobunídeo Mouillacitherium, o robiacinídeo Robiacina, o anfimerídeo Pseudamphimeryx [en]e os outros xifodontes Dichodon e Haplomeryx.[26]
Por volta de MP16, ocorreu uma mudança faunística, marcando o desaparecimento dos lofiodontes e dos hiraquiídeos europeus, bem como a extinção de todos os crocodilomorfos europeus, exceto pelo aligatoroide Diplocynodon [en].[25][49][50][51] As causas da mudança faunística foram atribuídas a uma transição de ambientes húmidos e altamente tropicais para florestas mais secas e temperadas com áreas abertas e vegetação mais abrasiva. As faunas herbívoras sobreviventes adaptaram suas dentições e estratégias alimentares para se adequar à vegetação abrasiva e sazonal.[52][53] Os ambientes ainda eram subúmidos e cheios de florestas subtropicais perenes, no entanto. A Palaeotheriidae era o único grupo de perissodáctilos europeu remanescente, e os artiodáctilos frugívoros-folívoros ou puramente folívoros tornaram-se o grupo dominante na Europa Ocidental.[54][42]
Eoceno Superior

A próxima espécie de Xiphodon a aparecer no registro fóssil foi X. intermedium de MP17a, onde é exclusiva. Após uma breve lacuna no registro fóssil em MP17b, a última espécie a aparecer foi X. gracilis em MP18. O xifodonte coexistiu em grande parte com as mesmas famílias de artiodáctilos, bem como com os Palaeotheriidae, na Europa Ocidental,[26] embora Cainotheriidae e os anoploteriídeos derivados Anoplotherium e Diplobune tenham feito suas primeiras aparições no registro fóssil em MP18.[24][55] Além disso, vários grupos de mamíferos migrantes chegaram à Europa Ocidental por volta de MP17a-MP18, nomeadamente Anthracotheriidae, Hyaenodontinae [en] e Amphicyonidae.[26] Além de cobras, sapos e salamandrídeos, uma rica assembleia de lagartos também é conhecida na Europa Ocidental de MP16-MP20, representando Iguanidae, Lacertidae, Gekkonidae, Agamidae, Scincidae, Helodermatidae e Varanoidea [en], a maioria dos quais conseguiu prosperar nas temperaturas quentes da Europa Ocidental.[56]
X. gracilis está bem representado em localidades da França, Espanha e Reino Unido.[26][57][58] Ele tem o registro fóssil mais longo conhecido dentro de seu gênero, durando de MP18 a MP20. Com base na localidade francesa de Escamps, de MP19, coexistiu com os herpetoteriídeos Peratherium e Amphiperatherium, o pseudorincocionídeo Pseudorhyncocyon, os nictitérios Saturninia e Amphidozotherium, vários morcegos e roedores, o omomiídeo Microchoerus [en], o adapídeo Palaeolemur [en], o hienailurino Pterodon [en], o anficíonídeo Cynodictis [en], os paleotérios Palaeotherium e Plagiolophus, o dicobunídeo Dichobune, o queropotamídeo Choeropotamus, os anoploteriídeos Anoplotherium e Diplobune [en], o cainotério Oxacron, o anfimerídeo Amphimeryx e os outros xifodontes Dichodon e Haplomeryx.[26]
Extinção

O evento Grande Coupure, durante o final do Eoceno até o início do Oligoceno (MP20-MP21), é uma das maiores e mais abruptas mudanças faunísticas do Cenozoico da Europa Ocidental e coincide com eventos de forçamento climático de climas mais frios e sazonais.[59] O evento levou à extinção de 60% das linhagens de mamíferos da Europa Ocidental, que foram subsequentemente substituídas por imigrantes asiáticos.[58][60][61] A Grande Coupure é frequentemente datada diretamente no limite Eoceno-Oligoceno, a 33,9 Ma, embora alguns estimem que o evento tenha começado um pouco mais tarde, a 33,6-33,4 Ma.[62][63] O evento ocorreu durante ou após a transição Eoceno-Oligoceno, uma mudança abrupta de um mundo de estufa quente que caracterizou grande parte do Paleogeno para um mundo de casa fria/casa de gelo a partir do Oligoceno Inferior. A queda maciça de temperaturas resulta da primeira grande expansão das camadas de gelo da Antártida, que causou diminuições drásticas de pCO2 e uma queda estimada de ~70 m no nível do mar.[64]
Muitos paleontólogos concordam que a glaciação e as consequentes quedas no nível do mar permitiram um aumento das migrações entre a Balcanatolia e a Europa Ocidental. O estreito de Turgai [en], que antes separava grande parte da Europa da Ásia, é frequentemente proposto como a principal barreira marítima europeia antes da Grande Coupure, mas alguns pesquisadores contestaram recentemente essa percepção, argumentando que ele já havia recuado completamente há 37 Ma, muito antes da transição Eoceno-Oligoceno. Em 2022, Alexis Licht et al. sugeriram que a Grande Coupure poderia possivelmente ter sido síncrona com a glaciação Oi-1 (33,5 Ma), que registra um declínio no CO2 atmosférico, impulsionando a glaciação antártica que já havia começado na transição Eoceno-Oligoceno.[45][65]
O evento Grande Coupure também marcou uma grande mudança faunística, com as chegadas de antracotérios posteriores, entelodontes, ruminantes (Gelocidae, Lophiomerycidae), rinocerotoides (Rhinocerotidae, Amynodontidae, Eggysodontidae [en]), carnívoros (Amphicyonidae posteriores, Amphicynodontidae, Nimravidae e Ursidae), roedores da Eurásia Oriental (Eomyidae [en], Cricetidae e Castoridae) e eulipotiflanos (Erinaceidae).[66][67][58][68]
Todos os três representantes, Xiphodon, Dichodon e Haplomeryx, são registrados pela última vez em localidades de MP20. O desaparecimento dos três gêneros significou a extinção completa da Xiphodontidae. Muitos outros gêneros de artiodáctilos da Europa Ocidental também desapareceram como resultado do evento de extinção da Grande Coupure.[27][58][24] As extinções de Xiphodon e de muitos outros mamíferos foram atribuídas a interações negativas com faunas imigrantes (competição, predação), mudanças ambientais decorrentes do resfriamento climático, ou alguma combinação dos dois.[62][27]
Referências
- ↑ Cuvier, Georges (1804). «Suite des Recherches: Suite de recherches sur les os fossiles de la pierre à plâtre des environs de Paris. Troisième mémoire. Restitution des pieds. Première section. Restitution des différens pieds de derrière.». Annales du Muséum National d'Histoire Naturelle, Paris (em francês). 3: 442–472. Consultado em 6 de março de 2024. Cópia arquivada em 27 de julho de 2023
- ↑ Cuvier, Georges (1805). «Troisième mémoire. Deuxième section. Restitution des différens pieds de devant.». Annales du Muséum National d'Histoire Naturelle, Paris (em francês). 6: 253–283. Consultado em 6 de março de 2024. Cópia arquivada em 10 de novembro de 2012
- ↑ Cuvier, Georges (1807). «Suite des recherches sur les os fossiles des environs de Paris. Troisième mémoire, troisième section, les phalanges. Quatrième mémoire sur les os des extrémités, première section, les os longs des extrémités postérieures.». Annales du Muséum d'Histoire Naturelle. 9: 10–44. Consultado em 6 de março de 2024. Cópia arquivada em 2 de setembro de 2023
- ↑ Cuvier, Georges (1808). «Cinquième mémoire. Troisième section. Vertèbres et côtes isolées». Annales du Muséum d'Histoire Naturelle. 12: 271–284
- ↑ Cuvier, Geoges (1812). «Résumé général et rétablissement des Squelettes des diverses espèces». Recherches sur les ossemens fossiles de quadrupèdes: où l'on rétablit les caractères de plusieurs espèces d'animaux que les révolutions du globe paroissent avoir détruites (em francês). 3. [S.l.]: Chez Deterville. Consultado em 6 de março de 2024. Cópia arquivada em 31 de julho de 2023
- ↑ Rudwick, Martin J. S. (1997). «Chapter 6: The Animals from the Gypsum Beds around Paris». Georges Cuvier, Fossil Bones, and Geological Catastrophes: New Translations and Interpretations of the Primary Texts. [S.l.]: University of Chicago Press
- ↑ Cuvier, Georges (1822). Recherches sur les ossemens fossiles, où l'on rétablit les caractères de plusieurs animaux dont les révolutions du globe ont détruit les espèces. 3. [S.l.]: G. Dufour and E. d'Ocagne. pp. 69–70. Consultado em 6 de março de 2024. Cópia arquivada em 19 de agosto de 2023
- ↑ Rudwick, Martin J.S. (1992). «Chapter 2: Keyholes into the Past». Scenes from Deep Time: Early Pictorial Representations of the Prehistoric World. [S.l.: s.n.] pp. 27–58
- ↑ Roberts, George (1839). An etymological and explanatory dictionary of the terms and language of geology (em inglês). London: Longman, Orme, Brown, Green, & Longmans. p. 183. Consultado em 29 de dezembro de 2021
- ↑ Gervais, Paul (1848–1852). Zoologie et paléontologie françaises (animaux vertébrés): ou nouvelles recherches sur les animaux vivants et fossiles de la France. 1. [S.l.]: Arthus Bertrand. pp. 90–91
- ↑ Gervais, Paul (1848–1852). «Note sur les genres Anthracotherium, Hyopotamus et Bothriodon». Zoologie et paléontologie françaises (animaux vertébrés): ou nouvelles recherches sur les animaux vivants et fossiles de la France. 2. [S.l.]: Arthus Bertrand. p. 2
- ↑ Owen, Richard (1848). «Description of Teeth and portions of Jaws of two extinct Anthracotherioid Quadrupeds (Hyopotamus vectianus and Hyop. bovinus) discovered by the Marchioness of Hastings in the Eocene Deposits on the N.W. coast of the Isle or Wight: with an attempt to develop Cuvier's idea of the Classification of Pachyderms by the Number of their Toes». The Quarterly Journal of the Geological Society of London. 4 (1–2): 103–141. Bibcode:1848QJGS....4..103O. doi:10.1144/GSL.JGS.1848.004.01-02.21
- ↑ Owen, Richard (1857). «Description of the Lower Jaw and Teeth of an anoplotherioid quadruped (Dichobune ovina, Ow.) of the size of the Xiphodon gracilis, Cuv., from the Upper Eocene Mart, Isle of Wight». The Quarterly Journal of the Geological Society of London. 13 (1–2): 254–260. Bibcode:1857QJGS...13..254O. doi:10.1144/GSL.JGS.1857.013.01-02.38
- ↑ a b Witton, Mark P.; Michel, Ellinor (2022). «Chapter 4: The sculptures: mammals». The Art and Science of the Crystal Palace Dinosaurs. [S.l.]: The Crowood Press. pp. 68–91
- ↑ Kovalevsky, Vladimir (1873). «On the Osteology of the Hyopotamidae». Philosophical Transactions of the Royal Society of London. 163: 19–94. Bibcode:1873RSPT..163...19K
- ↑ Flower, William Henry (1876). «Description of the Skull of a Species of Xiphodon, Cuvier». Proceedings of the Zoological Society of London. 44: 3–7. doi:10.1111/j.1096-3642.1876.tb02534.x
- ↑ Kovalevsky, Vladimir (1876). «Monographie der gattung Anthracotherium Cuv. und Versuch ein natürlich Classification der fossilen Hufthiere». Palaeontographica. 22: 241–247
- ↑ Gervais, Paul (1876). Zoologie et paléontologie générales. Nouvelles recherches sur les animaux vertébrés dont on trouve les ossements enfouis dans le sol et sur leur comparaison avec les espèces actuellement existantes. [S.l.]: A. Bertrand. p. 44
- ↑ Filhol, Henri (1884). «Pachydermes». Descriptions de Quelques Mammifères Fossiles des Phosphorites du Quercy. [S.l.]: Vialelle Printing Company and Co. p. 40
- ↑ Lydekker, Richard (1885). Catalogue of the fossil Mammalia in the British museum, (Natural History): Part II. Containing the Order Ungulata, Suborder Artiodactyla. [S.l.]: Order of the Trustees, London
- ↑ Schlosser, Max (1886). «Beiträge zur Kenntnis der Stammesgeschichte der Hufthiere und Versucheiner Systematik der Paar- und Unpaarhufer». Morphologisches Jahrbuch. 12: 95
- ↑ Stehlin, Hans Georg (1910). «Die Säugertiere des schweizerischen Eocaens. Sechster Teil: Catodontherium – Dacrytherium – Leptotherium – Anoplotherium – Diplobune – Xiphodon – Pseudamphimeryx – Amphimeryx – Dichodon – Haplomeryx – Tapirulus – Gelocus. Nachträge, Artiodactyla incertae sedis, Schlussbetrachtungen über die Artiodactylen, Nachträge zu den Perissodactylen». Abhandlungen der Schweizerischen Paläontologischen Gesellschaft. 36
- ↑ a b Hooker, Jerry J.; Weidmann, Marc (2000). Eocene Mammal Faunas of Mormont, Switzerland: Systematic Revision and Resolution of Dating Problems. 120. [S.l.]: Kommission der Schweizerischen Paläontologischen Abhandlungen. p. 95
- ↑ a b c d e f g h i j k l m n Erfurt, Jörg; Métais, Grégoire (2007). «Endemic European Paleogene Artiodactyls». In: Prothero, Donald R.; Foss, Scott E. The Evolution of Artiodactyls. [S.l.]: Johns Hopkins University Press. pp. 59–84
- ↑ a b c Franzen, Jens Lorenz (2003). «Mammalian faunal turnover in the Eocene of central Europe». Geological Society of America Special Papers. 369: 455–461. ISBN 9780813723693. doi:10.1130/0-8137-2369-8.455
- ↑ a b c d e f g h Aguilar, Jean-Pierre; Legendre, Serge; Michaux, Jacques (1997). «Synthèses et tableaux de corrélations». Actes du Congrès Bio-chroM'97. Mémoires et Travaux de l'EPHE Institut de Montpellier 21 (em francês). [S.l.]: École Pratique des Hautes Études-Sciences de la Vie et de la Terre, Montpellier. pp. 769–850
- ↑ a b c Weppe, Romain; Condamine, Fabien L.; Guinot, Guillaume; Maugoust, Jacob; Orliac, Maëva J. (2023). «Drivers of the artiodactyl turnover in insular western Europe at the Eocene–Oligocene Transition». Proceedings of the National Academy of Sciences. 120 (52): e2309945120. Bibcode:2023PNAS..12009945W. PMC 10756263
 . PMID 38109543. doi:10.1073/pnas.2309945120
. PMID 38109543. doi:10.1073/pnas.2309945120
- ↑ a b Weppe, Romain (2022). Déclin des artiodactyles endémiques européens, autopsie d'une extinction (Tese) (em francês). University of Montpellier. Consultado em 6 de março de 2024. Cópia arquivada em 11 de agosto de 2023
- ↑ Hooker, Jerry J. (2007). «Bipedal browsing adaptations of the unusual Late Eocene–earliest Oligocene tylopod Anoplotherium (Artiodactyla, Mammalia)». Zoological Journal of the Linnean Society. 151 (3): 609–659. doi:10.1111/j.1096-3642.2007.00352.x
- ↑ a b Luccisano, Vincent; Sudre, Jean; Lihoreau, Fabrice (2020). «Revision of the Eocene artiodactyls (Mammalia, Placentalia) from Aumelas and Saint-Martin-de-Londres (Montpellier limestones, Hérault, France) questions the early European artiodactyl radiation». Journal of Systematic Palaeontology. 18 (19): 1631–1656. Bibcode:2020JSPal..18.1631L. doi:10.1080/14772019.2020.1799253
- ↑ a b Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Escarguel, Gilles; Pélissié, Thierry; Antoine, Pierre-Olivier; Orliac, Maëva Judith (2020). «Cainotheriidae (Mammalia, Artiodactyla) from Dams (Quercy, SW France): phylogenetic relationships and evolution around the Eocene–Oligocene transition (MP19–MP21)» (PDF). Journal of Systematic Palaeontology. 18 (7): 541–572. Bibcode:2020JSPal..18..541W. doi:10.1080/14772019.2019.1645754. Consultado em 6 de março de 2024. Cópia arquivada (PDF) em 7 de março de 2022
- ↑ Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Pélissié, Thierry; Orliac, Maëva Judith (2020). "A new Cainotherioidea (Mammalia, Artiodactyla) from Palembert (Quercy, SW France): Phylogenetic relationships and evolutionary history of the dental pattern of Cainotheriidae". Palaeontologia Electronica (23(3):a54). doi:10.26879/1081. S2CID 229490410.
- ↑ a b c d e Sudre, Jean (1978). Les Artiodactyles de l'Eocéne moyen et supérieur d'Europe occidentale. [S.l.]: University of Montpellier
- ↑ a b Viret, Jean (1961). Traitè de Palèontologie. [S.l.]: Masson. pp. 887–1104
- ↑ a b c d e f g h i j k Dechaseaux, Colette (1967). «Artiodactyles des Phosphorites du Quercy: Étude sur le genre Xiphodon». Annales de Paléontologie. Vertébrés. 53: 27–47
- ↑ Dechaseaux, Colette (1963). «Une forme européenne du groupe des chameaux (Tylopodes): le genre Xiphodon». Comptes Rendus de l'Académie des Sciences. 256: 5607–5609
- ↑ Dechaseaux, Colette (1965). «Artiodactyles des phosphorites du Quercy. I. Étude sur le genre Dichodon». Annales de Paléontologie. Vertébrés. 51: 191–208
- ↑ Lihoreau, Fabrice; Boisserie, Jean-Renaud; Viriot, Laurent; Brunet, Michel (2006). «Anthracothere dental anatomy reveals a late Miocene Chado-Libyan bioprovince». Proceedings of the National Academy of Sciences. 103 (23): 8763–8767. Bibcode:2006PNAS..103.8763L. PMC 1482652. PMID 16723392. doi:10.1073/pnas.0603126103
- ↑ a b Depéret, Charles (1917). Monographie de la faune de mammifères fossiles du Ludien inférieur d'Euzet-les-Bains (Gard). [S.l.]: Lyon A. Rey
- ↑ Rodrigues, Helder Gomes; Lihoreau, Fabrice; Orliac, Maëva; Thewissen, J. G. M.; Boisserie, Jean-Renaud (2019). «Unexpected evolutionary patterns of dental ontogenetic traits in cetartiodactyl mammals». Proceedings of the Royal Society B. 286 (1896). PMC 6408598. PMID 30963938. doi:10.1098/rspb.2018.2417
- ↑ Sudre, Jean; Martinez, Jean-Noël (1995). «The astragalus of Paleogene artiodactyls: comparative morphology, variability and prediction of body mass». Lethaia. 28 (3): 197–209. Bibcode:1995Letha..28..197M. doi:10.1111/j.1502-3931.1995.tb01423.x
- ↑ a b c Blondel, Cécile (2001). «The Eocene-Oligocene ungulates from Western Europe and their environment» (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 168 (1–2): 125–139. Bibcode:2001PPP...168..125B. doi:10.1016/S0031-0182(00)00252-2
- ↑ Eronen, Jussi T.; Janis, Christine M.; Chamberlain, Charles Page; Mulch, Andreas (2015). «Mountain uplift explains differences in Palaeogene patterns of mammalian evolution and extinction between North America and Europe». Proceedings of the Royal Society B: Biological Sciences. 282 (1809): 20150136. PMC 4590438. PMID 26041349. doi:10.1098/rspb.2015.0136
- ↑ a b Maitre, Elodie (2014). «Western European middle Eocene to early Oligocene Chiroptera: systematics, phylogeny and palaeoecology based on new material from the Quercy (France)». Swiss Journal of Palaeontology. 133 (2): 141–242. Bibcode:2014SwJP..133..141M. doi:10.1007/s13358-014-0069-3
- ↑ a b Licht, Alexis; Métais, Grégoire; Coster, Pauline; İbilioğlu, Deniz; Ocakoğlu, Faruk; Westerweel, Jan; Mueller, Megan; Campbell, Clay; Mattingly, Spencer; Wood, Melissa C.; Beard, K. Christopher (2022). «Balkanatolia: The insular mammalian biogeographic province that partly paved the way to the Grande Coupure». Earth-Science Reviews. 226: 103929. Bibcode:2022ESRv..22603929L. doi:10.1016/j.earscirev.2022.103929
- ↑ Badiola, Ainara; Perales-Gogenola, Leire; Astibia, Humberto; Suberbiola, Xabier Pereda (2022). «A synthesis of Eocene equoids (Perissodactyla, Mammalia) from the Iberian Peninsula: new signs of endemism». Historical Biology. 34 (8): 1623–1631. Bibcode:2022HBio...34.1623B. doi:10.1080/08912963.2022.2060098
- ↑ Schmidt-Kittler, Norbert; Godinot, Marc; Franzen, Jens L.; Hooker, Jeremy J. (1987). «European reference levels and correlation tables». Münchner geowissenschaftliche Abhandlungen A10. [S.l.]: Pfeil Verlag, München. pp. 13–31
- ↑ Bai, Bin; Wang, Yuan-Qing; Theodor, Jessica M.; Meng, Jin (2023). «Small artiodactyls with tapir-like teeth from the middle Eocene of the Erlian Basin, Inner Mongolia, China». Frontiers in Earth Science. 11: 1–20. Bibcode:2023FrEaS..1117911B. doi:10.3389/feart.2023.1117911
- ↑ Martin, Jeremy E.; Pochat-Cottilloux, Yohan; Laurent, Yves; Perrier, Vincent; Robert, Emmanuel; Antoine, Pierre-Olivier (2022). «Anatomy and phylogeny of an exceptionally large sebecid (Crocodylomorpha) from the middle Eocene of southern France». Journal of Vertebrate Paleontology. 42 (4). Bibcode:2022JVPal..42E3828M. doi:10.1080/02724634.2023.2193828
- ↑ Martin, Jeremy E. (2015). «A sebecosuchian in a middle Eocene karst with comments on the dorsal shield in Crocodylomorpha». Acta Palaeontologica Polonica. 60 (3): 673–680. doi:10.4202/app.00072.2014
- ↑ Antunes, Miguel Telles (2003). «Lower Paleogene Crocodilians from Silveirinha, Portugal». Palaeovertebrata. 32: 1–26
- ↑ Robinet, Céline; Remy, Jean Albert; Laurent, Yves; Danilo, Laure; Lihoreau, Fabrice (2015). «A new genus of Lophiodontidae (Perissodactyla, Mammalia) from the early Eocene of La Borie (Southern France) and the origin of the genus Lophiodon Cuvier, 1822». Geobios. 48 (1): 25–38. Bibcode:2015Geobi..48...25R. doi:10.1016/j.geobios.2014.11.003
- ↑ Perales-Gogenola, Leire; Badiola, Ainara; Gómez-Olivencia, Asier; Pereda-Suberbiola, Xabier (2022). «A remarkable new paleotheriid (Mammalia) in the endemic Iberian Eocene perissodactyl fauna». Journal of Vertebrate Paleontology. 42 (4). Bibcode:2022JVPal..42E9447P. doi:10.1080/02724634.2023.2189447
- ↑ Solé, Floréal; Fischer, Valentin; Le Verger, Kévin; Mennecart, Bastien; Speijer, Robert P.; Peigné, Stéphane; Smith, Thierry (2022). «Evolution of European carnivorous mammal assemblages through the Paleogene». Biological Journal of the Linnean Society. 135 (4): 734–753. doi:10.1093/biolinnean/blac002
- ↑ Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Escarguel, Gilles; Pelissie, Thierry; Antoine, Pierre-Olivier; Orliac, Maeva J. (2020). «Cainotheriidae (Mammalia, Artiodactyla) from Dams (Quercy, SW France): phylogenetic relationships and evolution around the Eocene–Oligocene transition (MP19–MP21)» (PDF). Journal of Systematic Palaeontology. 18 (7): 541–572. Bibcode:2020JSPal..18..541W. doi:10.1080/14772019.2019.1645754
- ↑ Rage, Jean-Claude (2012). «Amphibians and squamates in the Eocene of Europe: what do they tell us?». Palaeobiodiversity and Palaeoenvironments. 92 (4): 445–457. Bibcode:2012PdPe...92..445R. doi:10.1007/s12549-012-0087-3
- ↑ Minwer-Barakat, Raef; Badiola, Ainara; Marigó, Judit; Moyà-Solà, Salvador (2013). «First record of the genus Microchoerus (Omomyidae, Primates) in the western Iberian Peninsula and its palaeobiogeographic implications». Journal of Human Evolution. 65 (3): 313–321. Bibcode:2013JHumE..65..313M. PMID 23916791. doi:10.1016/j.jhevol.2013.07.002
- ↑ a b c d Hooker, Jerry J.; Collinson, Margaret E.; Sille, Nicholas P. (2004). «Eocene–Oligocene mammalian faunal turnover in the Hampshire Basin, UK: calibration to the global time scale and the major cooling event» (PDF). Journal of the Geological Society. 161 (2): 161–172. Bibcode:2004JGSoc.161..161H. doi:10.1144/0016-764903-091. Consultado em 6 de março de 2024. Cópia arquivada (PDF) em 8 de agosto de 2023
- ↑ Sun, Jimin; Ni, Xijun; Bi, Shundong; Wu, Wenyu; Ye, Jie; Meng, Jin; Windley, Brian F. (2014). «Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene Boundary in Asia». Scientific Reports. 4 (7463): 7463. Bibcode:2014NatSR...4.7463S. PMC 4264005. PMID 25501388. doi:10.1038/srep07463
- ↑ Legendre, Serge; Mourer-Chauviré, Cécile; Hugueney, Marguerite; Maitre, Elodie; Sigé, Bernard; Escarguel, Gilles (2006). «Dynamique de la diversité des mammifères et des oiseaux paléogènes du Massif Central (Quercy et Limagnes, France)». STRATA. 1 (em francês). 13: 275–282
- ↑ Escarguel, Gilles; Legendre, Serge; Sigé, Bernard (2008). «Unearthing deep-time biodiversity changes: The Palaeogene mammalian metacommunity of the Quercy and Limagne area (Massif Central, France)». Comptes Rendus Geoscience. 340 (9–10): 602–614. Bibcode:2008CRGeo.340..602E. doi:10.1016/j.crte.2007.11.005. Consultado em 6 de março de 2024. Cópia arquivada em 13 de outubro de 2023
- ↑ a b Costa, Elisenda; Garcés, Miguel; Sáez, Alberto; Cabrera, Lluís; López-Blanco, Miguel (2011). «The age of the "Grande Coupure" mammal turnover: New constraints from the Eocene–Oligocene record of the Eastern Ebro Basin (NE Spain)». Palaeogeography, Palaeoclimatology, Palaeoecology. 301 (1–4): 97–107. Bibcode:2011PPP...301...97C. doi:10.1016/j.palaeo.2011.01.005. hdl:2445/34510
- ↑ Hutchinson, David K.; Coxall, Helen K.; Lunt, Daniel J.; Steinthorsdottir, Margret; De Boer, Agatha M.; Baatsen, Michiel L.J.; Von der Heydt, Anna S.; Huber, Matthew; Kennedy-Asser, Alan T.; Kunzmann, Lutz; Ladant, Jean-Baptiste; Lear, Caroline; Moraweck, Karolin; Pearson, Paul; Piga, Emanuela; Pound, Matthew J.; Salzmann, Ulrich; Scher, Howie D.; Sijp, Willem P.; Śliwińska, Kasia K; Wilson, Paul A.; Zhang, Zhongshi (2021). «The Eocene-Oligocene transition: A review of marine and terrestrial proxy data, models and model-data comparisons». Climate of the Past. 17 (1): 269–315. Bibcode:2021CliPa..17..269H. doi:10.5194/cp-17-269-2021. hdl:11250/3135351
- ↑ Toumoulin, Agathe; Tardif, Delphine; Donnadieu, Yannick; Licht, Alexis; Ladant, Jean-Baptiste; Kunzmann, Lutz; Dupont-Nivet, Guillaume (2022). «Evolution of continental temperature seasonality from the Eocene greenhouse to the Oligocene icehouse –a model–data comparison». Climate of the Past. 18 (2): 341–362. Bibcode:2022CliPa..18..341T. doi:10.5194/cp-18-341-2022
- ↑ Boulila, Slah; Dupont-Nivet, Guillaume; Galbrun, Bruno; Bauer, Hugues; Châteauneuf, Jean-Jacques (2021). «Age and driving mechanisms of the Eocene–Oligocene transition from astronomical tuning of a lacustrine record (Rennes Basin, France)». Climate of the Past. 17 (6): 2343–2360. Bibcode:2021CliPa..17.2343B. doi:10.5194/cp-17-2343-2021
- ↑ Rivals, Florent; Belyaev, Ruslan I.; Basova, Vera B.; Prilepskaya, Natalya E. (2023). «Hogs, hippos or bears? Paleodiet of European Oligocene anthracotheres and entelodonts». Palaeogeography, Palaeoclimatology, Palaeoecology. 611: 111363. Bibcode:2023PPP...61111363R. doi:10.1016/j.palaeo.2022.111363
- ↑ Becker, Damien (2009). «Earliest record of rhinocerotoids (Mammalia: Perissodactyla) from Switzerland: systematics and biostratigraphy». Swiss Journal of Geosciences. 102 (3): 489–504. Bibcode:2009SwJG..102..489B. doi:10.1007/s00015-009-1330-4
- ↑ Solé, Floréal; Fischer, Fischer; Denayer, Julien; Speijer, Robert P.; Fournier, Morgane; Le Verger, Kévin; Ladevèze, Sandrine; Folie, Annelise; Smith, Thierry (2020). «The upper Eocene-Oligocene carnivorous mammals from the Quercy Phosphorites (France) housed in Belgian collections». Geologica Belgica. 24 (1–2): 1–16. doi:10.20341/gb.2020.006