Anoplotherium
Anoplotherium
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Eoceno Superior – Oligoceno inferior 37–33,4 Ma | |||||||||||||||
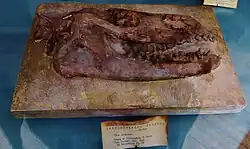
 Esqueleto incompleto de Anoplotherium commune da comuna de Pantin, Museu Nacional de História Natural, França | |||||||||||||||
| Classificação científica | |||||||||||||||
| |||||||||||||||
| Espécie-tipo | |||||||||||||||
| †Anoplotherium commune Cuvier, 1804 | |||||||||||||||
| Outras espécies | |||||||||||||||
| |||||||||||||||
| Sinónimos | |||||||||||||||
Sinonímia do gênero
Sinônimos de A. commune
Sinônimos de A. latipes
Sinônimos de A. laurillardi
| |||||||||||||||
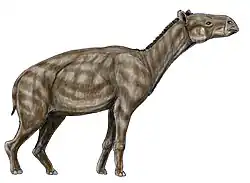
Anoplotherium é o gênero-tipo [en] da extinta família de artiodáctilos do Paleogeno, Anoplotheriidae, que era endêmica da Europa Ocidental. Viveu do Eoceno Superior ao começo do Oligoceno. Foi o quinto gênero de mamífero fóssil a ser descrito com autoridade taxonômica oficial, com uma história que remonta a 1804, quando seus fósseis de Montmartre, em Paris, França, foram descritos pela primeira vez pelo naturalista francês Georges Cuvier. Descobertas de esqueletos incompletos de Anoplotherium commune em 1807 levaram Cuvier a descrever minuciosamente características incomuns para as quais não existem análogos modernos. Suas reconstruções esqueléticas e musculares desenhadas de A. commune em 1812 foram um dos primeiros exemplos de reconstruções anatômicas baseadas em evidências fósseis. As contribuições de Cuvier para a paleontologia, baseadas em seus trabalhos sobre o gênero, foram revolucionárias para o campo, não apenas provando as ideias em desenvolvimento de extinção e sucessão ecológica, mas também abrindo caminho para subcampos como a paleoneurologia. Hoje, são conhecidas quatro espécies.
Anoplotherium estava entre os maiores artiodáctilos não whippomorfos do período Paleogeno, pesando em média de 115 kg a 271 kg e medindo pelo menos 2,5 m de comprimento de cabeça e corpo e 1,25 m de altura no ombro. Era um artiodáctilo evolutivamente avançado e incomum, ostentando pés de três dedos em certas espécies como Anoplotherium latipes, uma cauda longa e robusta, e um cérebro altamente desenvolvido com forte suporte tanto para o olfato quanto para a percepção sensorial. Sua constituição geral robusta pode tê-lo permitido ficar de pé bipedalmente para se alimentar de plantas em alturas maiores, atingindo aproximadamente 3 m de altura, competindo eficazmente com os poucos outros herbívoros de médio a grande porte com os quais conviveu. A extensão total de seu bipedalismo, no entanto, precisa ser confirmada por mais pesquisas. O maior, Anoplotherium commune de dois dedos, e o ligeiramente menor, Anoplotherium latipes de três dedos, podem ser dimorfos sexuais, sendo o primeiro fêmea e o último macho, mas essa ideia permanece especulativa. Seu parente mais próximo era Diplobune [en], que, de forma semelhante, é hipotetizado como tendo tido comportamentos especializados.
O artiodáctilo viveu na Europa Ocidental quando esta era um arquipélago isolado do resto da Eurásia, o que significa que viveu num ambiente com várias outras faunas que também evoluíram com altos níveis de endemismo. Suas origens exatas são desconhecidas, mas surgiu muito depois de uma mudança para condições mais secas, mas ainda subúmidas, que levaram a plantas abrasivas e à extinção de Lophiodontidae [en] de grande porte, alcançando gigantismo e estabelecendo-se como um herbívoro dominante em toda a região da Europa Ocidental, dada a sua abundante evidência fóssil.
Seu sucesso foi abruptamente interrompido pelo evento de extinção e renovação faunística da Grande Coupure no Oligoceno mais primitivo da Europa Ocidental, que foi causado por mudanças em direção a maior glaciação e sazonalidade. As florestas tropicais e subtropicais foram rapidamente substituídas por ambientes mais temperados, e a maioria das barreiras oceânicas que anteriormente separavam a Europa Ocidental da Eurásia oriental se fecharam, permitindo grandes dispersões faunísticas da Ásia. Embora as causas específicas sejam incertas, Anoplotherium provavelmente foi incapaz de se adaptar a essas grandes mudanças e sucumbiu à extinção.
Taxonomia
História da pesquisa
Identificações

Embora Georges Cuvier soubesse da existência de ossos fósseis nas pedreiras de gesso dos arredores de Paris (conhecidas como bacia de Paris [en]) já em 1800, foi apenas em 1804 que ele os descreveu. Após descrever Palaeotherium [en], ele escreveu sobre o próximo conjunto de fósseis que conseguiu discernir como sendo diferentes de Palaeotherium com base na forma da dentição, incluindo a aparente falta de caninos que deixava um grande espaço entre os incisivos e os pré-molares. Ele observou que a hemimandíbula (meia mandíbula) tinha três incisivos inferiores em vez de quatro incisivos ou nenhum, o que, segundo ele, caracterizava outros "paquidermes". Cuvier, baseando o nome em sua aparente falta de braços e caninos adequados para ataques ofensivos, criou o nome Anoplotherium.[1][2]
O nome do gênero Anoplotherium significa "besta desarmada" e é um composto das palavras gregas αν- (an, 'não'), ὅπλον (hóplon, 'armadura, grande escudo') e θήρ (thēr, 'besta, animal selvagem').[3]
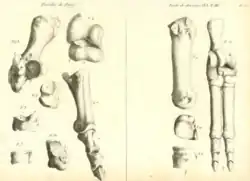
Cuvier nomeou três espécies de Anoplotherium no mesmo ano, a primeira das quais foi Anoplotherium commune, do "tamanho de uma ovelha", e as outras três eram "espécies menores" que ele nomeou Anoplotherium medium, Anoplotherium minus e Anoplotherium minimum. A etimologia do nome da espécie Anoplotherium commune refere-se a quão "comuns" eram os fósseis da espécie, enquanto as etimologias das outras duas espécies foram baseadas em seus tamanhos em comparação com A. commune.[a] Ele também atribuiu um casco fendido [en] (ou casco didáctilo) a A. commune, uma vez que o espécime parecia ser de grande porte. Ele pensou que Anoplotherium tinha cascos didáctilos em vez de cascos tridáctilos, o que o separaria de Palaeotherium. Com base nos cascos e na dentição, ele concluiu que Anoplotherium era semelhante a ruminantes ou camelídeos.[4][5] No entanto, em 1807, Cuvier descobriu que Anoplotherium commune tinha três dedos em seus membros posteriores, embora os terceiros dedos indicadores fossem de tamanhos menores em comparação com os outros dois.[6]
Esqueletos
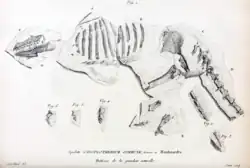
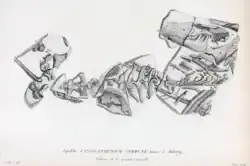
Em 1807, Cuvier escreveu sobre dois esqueletos incompletos que haviam sido recentemente descobertos, embora o primeiro estivesse parcialmente danificado porque não foi coletado com cuidado (o que ele expressou como tendo frustrado sua compreensão da anatomia esquelética de Anoplotherium inicialmente). O primeiro esqueleto, encontrado nas pedreiras de Montmartre, na comuna de Pantin, ajudou a confirmar as diagnósticos anteriores de Cuvier sobre Anoplotherium como corretas. O esqueleto embutido era do tamanho de um cavalo pequeno e ajudou a confirmar os grandes pés didáctilos e os 44 dentes totais que possuía (11 de cada lado da mandíbula). Também tinha 11 costelas completas e um fragmento de uma 12ª, correspondendo ao número de costelas dos camelídeos. O elemento mais surpreendente para Cuvier, no entanto, foi a enorme cauda com 22 vértebras no esqueleto, uma característica que ele disse que não teria conhecido anteriormente, pois não há análogos modernos da cauda alongada e espessa em nenhum grande mamífero quadrúpede.[7]
O segundo esqueleto incompleto veio de Antony, desta vez removido com mais cuidado sob a supervisão de especialistas do que o primeiro. Nele, ele pôde confirmar seis vértebras lombares e três vértebras sacrais, todas extremamente fortes e que provavelmente sustentavam a longa cauda. O mais notável para Cuvier foi a confirmação de que Anoplotherium tinha dois dedos grandes e um dedo pequeno em suas patas dianteiras, o que era incomum para mamíferos aparentados.[7]
Importância na história da paleontologia
Embora Palaeotherium e Anoplotherium não sejam bem reconhecidos em comparação com animais fósseis de outros períodos (isto é, dinossauros do Mesozoico e mamíferos do Neogeno-Quaternário), suas descobertas de fósseis em Montmartre e as descrições formais por Cuvier são reconhecidas como momentos críticos que pioneiraram a paleontologia para a era moderna. Ao contrário dos gêneros de fósseis do Pleistoceno nas Américas no início da história da paleontologia, como Megatherium e Mammut, Palaeotherium e Anoplotherium não foram encontrados em depósitos superficiais, mas embutidos em depósitos de rocha mais profundos e mais duros, datados do Eoceno. As pessoas em Paris já estavam familiarizadas há séculos com esqueletos de animais em sua área, alguns dos quais foram posteriormente guardados e formalmente descritos. No entanto, foi Cuvier quem formalmente erigiu dois gêneros de fósseis que vieram de depósitos mais antigos, e de sua terra natal no continente europeu, em vez das Américas, onde Megatherium e Mammut foram encontrados.[8] Os fósseis do Paleogeno não deixaram evidências de descendentes posteriores, extintos ou existentes, embora as semelhanças de Palaeotherium com as antas tenham dificultado a comprovação da teoria. Ele notou que abaixo do gesso havia sedimentos mais antigos de conchas marinhas e répteis, como o que Cuvier descreveu como um "crocodilo" gigante, que mais tarde seria conhecido como Mosasaurus. Cuvier soube então que o mundo de onde Anoplotherium e Palaeotherium vieram era um período de tempo diferente antes daquele do tempo precedente dos répteis marinhos e dos tempos seguintes de Megatherium e Mammut, provando assim o conceito de extinção natural.[9]
As descrições de Cuvier de um endocast (caixa craniana fossilizada) de um hemisfério cerebral pertencente a um crânio quebrado de Anoplotherium commune de Montmartre, a partir de 1804 até 1822, são reconhecidas como o primeiro verdadeiro exemplo de paleoneurologia, o estudo da evolução do cérebro. A primeira definição de um "endocasto" data de 1822, quando Cuvier descreveu um molde do cérebro de A. commune, notando que ele oferecia pistas sobre a verdadeira forma do cérebro do mamífero agora extinto (embora mais tarde se descobrisse que era uma porção do cérebro, e não a sua totalidade). Desde o primeiro estudo de endocasto, muitos outros estudos cerebrais foram conduzidos para outros mamíferos fósseis ao longo da segunda metade do século XIX em diante.[10][11][12] Uma descrição de Cuvier de 1822 de um fêmur fraturado e curado de A. commune é citada como um dos primeiros exemplos de paleopatologia, o estudo de doenças e lesões antigas em organismos pré-históricos.[13][14][15]
Primeiras representações
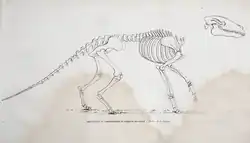
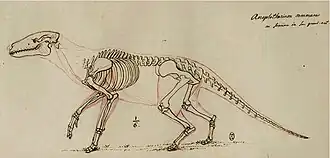
Em 1812, Cuvier publicou seu desenho de uma reconstrução esquelética de A. commune com base em restos fósseis conhecidos da espécie, incluindo os esqueletos incompletos mencionados anteriormente. Com base na constituição robusta da espécie de mamífero, ele hipotetizou que sua estrutura corporal era semelhante à das lontras, exceto pelas pernas, que era adaptada à vida semiaquática, nadando para consumir plantas aquáticas, sem orelhas longas, semelhante aos mamíferos semiaquáticos, e vivendo em ambientes pantanosos. Cuvier sugeriu que seu estilo de vida era, portanto, semelhante ao de mamíferos quadrúpedes semiaquáticos, como hipopótamos e roedores muroides. Ele pensava que, em comparação, outras espécies de Anoplotherium, como A. medium e A. minus, eram adaptadas a comportamentos terrestres e alimentação mista (pastoreio e ramoneio).[16][17] Hoje, a reconstrução da anatomia esquelética envelheceu bem, resistindo em grande parte ao teste do tempo desde 1812.[18] Anoplotherium e Palaeotherium também foram representados em desenhos de 1822 pelo paleontólogo francês Charles Léopold Laurillard, sob a direção de Cuvier, embora as restaurações não fossem tão detalhadas quanto as de Cuvier.[19]

A reconstrução de Anoplotherium como um nadador aquático foi apoiada por vários paleontólogos europeus do século XIX e persistiu por mais de um século[20][21] até 1938, quando M. Dor rejeitou a teoria do gênero como sendo adaptado à água, com base em diferenças anatômicas com lontras e hipopótamos que contradizem os comportamentos semiaquáticos e são mais consistentes com a vida terrestre.[22] Essa rejeição foi apoiada por Jerry J. Hooker em 2007 e por Svitozar Davydenko et al. em 2023, com base em características anatômicas, embora o primeiro tenha discordado das observações de Dor sobre a cauda. Hooker argumentou que, embora as vértebras caudais distais do anoplotério sejam menos proeminentes que as dos cangurus (Macropus), os padrões vertebrais de Anoplotherium são mais semelhantes a Macropus do que a ungulados como Bos ou Equus. Hoje, acredita-se que Anoplotherium seja um herbívoro terrestre com comportamentos especializados.[23][24]
A. commune foi notavelmente representado na atração dos Dinossauros do Crystal Palace [en], no Parque Crystal Palace [en], no Reino Unido, aberta ao público desde 1854 e construída pelo escultor inglês Benjamin Waterhouse Hawkins. Mais especificamente, foram feitas três estátuas de A. commune, duas das quais estão de pé e a terceira em posição de repouso. Essas estátuas assemelham-se a híbridos de cervos e grandes felinos e medem 3,6 m de comprimento. Sua inclusão no Parque Crystal Palace reflete a popularidade e o interesse público em Anoplotherium no século XIX, pois era um ícone da paleontologia, geologia e história natural, sendo regularmente incorporado em textos e salas de aula de paleontologia (sua popularidade diminuiu desde o século XX).[25][26]
As esculturas de A. commune foram, em geral, baseadas em Hawkins seguindo de perto a descrição de Cuvier do gênero com base nos restos conhecidos, incluindo as especulações musculares robustas não publicadas de Cuvier, que são consideradas precisas pelos padrões modernos. Hawkins também se desviou das descrições de Cuvier, no entanto, provavelmente baseando seus desenhos faciais e a presença imprecisa de membros tetradáctilos (quatro dedos em cada pé) em vez de membros didáctilos ou tridáctilos nos camelídeos existentes. Além desses erros, as estátuas têm sido em grande parte precisas em relação às representações modernas de Anoplotherium.[26]
Confusões com outros grupos de mamíferos
.jpg)
Durante grande parte do século XIX, os paleontólogos confundiram mamíferos de outras famílias com Anoplotherium, em grande parte devido ao fato de a paleontologia estar em seus estágios iniciais. Um dos exemplos mais antigos é de 1822, quando Cuvier erigiu os nomes A. gracile, A. murinum, A. obliquum, A. leporinum e A. secundaria, substituindo nomes de espécies anteriores dentro de Anoplotherium, exceto A. commune. Em A. gracile, ele notou diferenças nos molares que o levaram a erigir o subgênero Xiphodon. Para A. leporinum, A. murinum e A. obliquum, o subgênero Dichobune [en] foi criado por Cuvier com base em seu pequeno tamanho.[11] Em 1848, o paleontólogo francês Auguste Pomel promoveu os 2 subgêneros a nível de gênero e erigiu um gênero adicional, Amphimeryx [en], para A. murinus e A. obliquus. As taxonomias revisadas foram seguidas por paleontólogos subsequentes, como Paul Gervais. Portanto, as espécies não são mais classificadas como Anoplotherium, mas como gêneros distantes.[27][28][29]
Outros mamíferos inicialmente confundidos com o gênero Anoplotherium, mas eventualmente reclassificados no século XIX, representavam a família endêmica de artiodáctilos europeus Cainotheriidae (Cainotherium [en][30][31][29][32]), membros europeus e do subcontinente indiano da família de perissodáctilos Chalicotheriidae (Anisodon [en][33][34][35] e Nestoritherium [en][36][37][38]), e até mesmo membros endêmicos da América do Sul da ordem Litopterna (Scalabrinitherium [en] e Proterotherium [en][39]).
Revisões dentro dos Anoplotheriidae

Em 1851, Pomel observou que as espécies de Anoplotherium podiam ser determinadas como tendo cascos didáctilos (terceiro dedo reduzido) ou tridáctilos (terceiro dedo mais desenvolvido) e que as únicas espécies previamente erigidas que eram válidas eram A. commune e A. secundaria. Além disso, ele erigiu três novas espécies com base em restos adicionais: A. duvmoyi (baseado nas ilustrações de fósseis de A. commune de Cuvier), A. platypus, A. laurillardi (incisivos convexos na superfície anterior) e A. cuvieri. A. laurillardi deriva como nome de espécie de Charles Laurillard.[31]
O paleontólogo francês Paul Gervais, em 1852, nomeou o gênero Eurytherium com base na presença de cascos tridáctilos em vez de didáctilos, pois tornou a nova espécie E. latipis a espécie-tipo e Anoplotherium platypus um sinônimo da primeira.[20] Henri Filhol [en] seguiria Gervais ao erigir Eurytherium quercyi e E. minus com base nos tamanhos dentários e reclassificar Anoplotherium secundarium (ou A. secundaria) para Eurytherium.[40]
Em 1862, Ludwig Rütimeyer [en] erigiu o subgênero Diplobune [en] para o gênero Dichobune, com base no fato de que era uma transição evolutiva entre Anoplotherium secundarium e os dichobunídeos.[41] No entanto, foi promovido a um gênero distinto com uma espécie, Dichobune bavaricum, sendo colocada no gênero por Oscar Fraas [en] em 1870.[42]
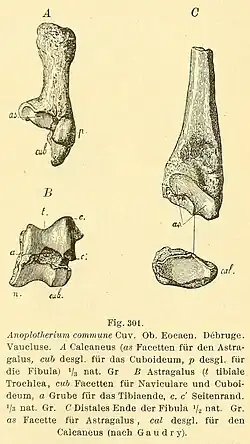
Em 1883, Max Schlosser [en] tornou Eurytherium um sinônimo de Anoplotherium porque argumentou que as anatomias dos membros e as dentições eram diferenças específicas em características, em vez de diferenças maiores que definiam um gênero inteiro. Schlosser destacou que todas as espécies de Anoplotherium de alguma forma tinham três dedos, apesar de A. commune ter os terceiros dedos menos desenvolvidos que A. latipes. Ele também reforçou a ideia de que "A. platypus" é um sinônimo de A. latipes. O nome A. latipes tem prioridade sobre A. platypus até os dias de hoje porque Pomel, em 1851, não listou nenhum espécime para a espécie, tornando-a efetivamente um nomen dubium. Ele também mencionou que o status de A. duvmoyi não era estável por se basear em ilustrações, o que considerou um "esforço sem esperança". Ele também apoiou Diplobune como um gênero válido, argumentando que Anoplotherium secundaria deveria ser renomeada para Diplobune secundaria com base na dentição e no tamanho menor. Schlosser também disse que Anoplotherium cuvieri era uma espécie inválida porque a diagnose baseada em ossos metatarsais isolados não era válida o suficiente.[21][29][23]
Richard Lydekker erigiu a espécie A. cayluxense em 1885 com base em seu tamanho menor e variações únicas nas cúspides dos molares. Ele também rebaixou o gênero Diplobune a sinônimo de Anoplotherium, o que significa que as espécies do primeiro foram adicionadas/readicionadas a Anoplotherium como A. secundarium, A. quercyi, A. modicum, A. bavaricum e A. minus (= A. minor, Filhol 1877).[29] A sinonímia de Diplobune com Anoplotherium não foi apoiada por Hans Georg Stehlin [en] em 1910, pois ele argumentou que o primeiro era genericamente distinto do último, apesar de suas relações próximas, restaurando assim as espécies anteriores para Diplobune (com exceção de D. modicum, que ele sinonimizou com D. bavarica) e adicionando "Anoplotherium secundarium" a Diplobune como D. secundaria. Ele também escreveu que Anoplotherium cayluxense era um sinônimo de Diplobune secundaria. Stehlin também atribuiu tentativamente "Anoplotherium" obliquum ao gênero Haplomeryx [en] como H? obliquum. Como resultado das revisões, as únicas espécies válidas de Anoplotherium eram A. commune, A. latipes e A. laurillardi.[43]
Em 1922, Wilhelm Otto Dietrich [en] erigiu a quarta espécie, A. pompeckji, da localidade de Mähringen, na Alemanha, nomeada em homenagem ao paleontólogo alemão Josef Felix Pompeckj. A espécie foi descrita como uma espécie tridáctila de tamanho médio, com membros dianteiros de 4 dedos e membros traseiros de 3 dedos, com proporções de ossos da mão mais delgadas e um astrágalo menor.[44] A. pompeckji é a espécie menos caracterizada e tem uma dentição semelhante à de A. laurillardi, tornando seu status menos certo em comparação com as outras três espécies.[23][45]
Em 1964, o paleontólogo Louis de Bonis revisou brevemente os sinônimos taxonômicos de Anoplotherium, considerando que A. duvernoyi se baseava em um indivíduo jovem com características de incisivos que Pomel não especificou e que A. cuvieri não difere nas dimensões dos metacarpos de A. laurillardi. Ele seguiu Stehlin ao reconhecer as três espécies principais de Anoplotherium, embora não tenha mencionado A. pompeckji em sua revisão.[46]
Classificação

Anoplotherium é o gênero-tipo [en] de Anoplotheriidae, uma família de artiodáctilos do Paleogeno endêmica da Europa Ocidental que viveu do Eoceno Médio ao Oligoceno Inferior (~44 a 30 Ma, com um possível registro mais antigo em ~48 Ma). As origens evolutivas exatas e as dispersões dos anoploteriídeos são incertas, mas eles residiram exclusivamente no continente quando este era um arquipélago isolado por barreiras marítimas de outras regiões, como a Balkanatolia e o resto da Eurásia oriental. As relações de Anoplotheriidae com outros membros de Artiodactyla não estão bem resolvidas, com alguns determinando que seja um tilópode (que inclui camelídeos e mericoidodontes do Paleogeno) ou um parente próximo da infraordem, e outros acreditando que possa ter sido mais próximo de Ruminantia (que inclui tragulídeos e outros parentes próximos do Paleogeno).[47][45]
Anoplotheriidae consistem em duas subfamílias, Dacrytheriinae e Anoplotheriinae, sendo esta última a subfamília mais jovem à qual Anoplotherium pertence. Dacrytheriinae é a subfamília mais antiga das duas, que apareceu pela primeira vez no Eoceno Médio (desde as Zonas de Mamíferos do Paleogeno [en], unidade MP13, possivelmente até MP10), embora alguns autores os considerem uma família separada, como Dacrytheriidae.[48][49] Os anoploteriíneos fizeram suas primeiras aparições no Eoceno Superior (MP15-MP16), ou ~41-40 Ma, na Europa Ocidental com Duerotherium [en] e Robiatherium [en]. No entanto, por MP17a-MP17b, há uma lacuna notável no registro fóssil dos anoploteriíneos em geral, pois os dois gêneros anteriores parecem ter feito suas últimas aparições no nível MP anterior, MP16.[50]
Por MP18, Anoplotherium e Diplobune fizeram suas primeiras aparições na Europa Ocidental, mas suas origens exatas são desconhecidas. Os dois gêneros eram amplamente distribuídos por toda a Europa Ocidental, com base em abundantes evidências fósseis que abrangem Portugal, Espanha, Reino Unido, França, Alemanha e Suíça durante grande parte da Europa pré-Grande Coupure (antes de MP21), o que significa que eram elementos típicos do Eoceno Superior até o Oligoceno Inferior.[51][50][45] Os anoploteriíneos mais antigos são considerados espécies menores, enquanto os mais recentes eram maiores. Anoplotherium e Diplobune são considerados os anoploteriídeos mais derivados (ou evolutivamente recentes) com base na morfologia dentária e alcançaram gigantismo entre os artiodáctilos não-whippomorfos, tornando-os alguns dos maiores artiodáctilos não-whippomorfos do Paleogeno, bem como alguns dos maiores mamíferos a vagar pela Europa Ocidental na época (todas as espécies de Anoplotherium eram grandes a muito grandes, enquanto nem todas as espécies de Diplobune eram grandes).[45][52][53][12]

A realização de estudos focados nas relações filogenéticas dentro de Anoplotheriidae tem se mostrado difícil devido à escassez geral de espécimes fósseis da maioria dos gêneros.[50] As relações filogenéticas de Anoplotheriidae, bem como de Xiphodontidae, Mixtotheriidae [en] e Cainotheriidae, também têm sido elusivas devido às morfologias selenodontes dos molares, que eram convergentes com os tilópodes ou ruminantes.[53] Alguns pesquisadores consideraram as famílias selenodontes Anoplotheriidae, Xiphodontidae e Cainotheriidae como estando dentro de Tylopoda devido a características pós-cranianas que eram semelhantes aos tilópodes da América do Norte no Paleogeno.[23] Outros pesquisadores os relacionam mais proximamente aos ruminantes do que aos tilópodes, com base na morfologia dentária. Diferentes análises filogenéticas produziram resultados diferentes para as famílias de artiodáctilos europeus do Eoceno "derivados" selenodontes, tornando incerto se eles eram mais próximos de Tylopoda ou de Ruminantia.[54][55]
Num artigo publicado em 2019, Romain Weppe et al. conduziram uma análise filogenética sobre Cainotherioidea dentro de Artiodactyla, baseada em características mandibulares e dentárias, especificamente em termos de relações com artiodáctilos do Paleogeno. Os resultados indicaram que a superfamília estava proximamente relacionada com Mixtotheriidae e Anoplotheriidae. Eles determinaram que Cainotheriidae, Robiacinidae, Anoplotheriidae e Mixtotheriidae formavam um clado que era o grupo irmão de Ruminantia, enquanto Tylopoda, juntamente com Amphimerycidae [en] e Xiphodontidae, se separaram mais cedo na árvore.[55] A árvore filogenética usada para o jornal e outro trabalho publicado sobre os cainoterioídeos é delineada abaixo:[56]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Em 2020, Vincent Luccisano et al. criaram uma árvore filogenética dos artiodáctilos basais, a maioria endêmica da Europa Ocidental, do Paleogeno. Em um clado, os "endêmicos europeus bunoselenodontes" Mixtotheriidae, Anoplotheriidae, Xiphodontidae, Amphimerycidae, Cainotheriidae e Robiacinidae são agrupados com Ruminantia. A árvore filogenética produzida pelos autores é mostrada abaixo:[54]
| Artiodactyla |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Em 2022, Weppe criou uma análise filogenética em sua tese acadêmica sobre as linhagens de artiodáctilos do Paleogeno, focando mais especificamente nas famílias endêmicas europeias. A árvore filogenética, segundo Weppe, é a primeira a conduzir afinidades filogenéticas de todos os gêneros de anoploteriídeos, embora nem todas as espécies individuais tenham sido incluídas. Ele descobriu que Anoplotheriidae, Mixtotheriidae e Cainotherioidea formam um clado com base em características dentárias sinapomórficas (características que se acredita terem se originado de seu ancestral comum mais recente). O resultado, mencionou Weppe, corresponde a análises filogenéticas anteriores sobre os Cainotherioidea com outros artiodáctilos endêmicos do Paleogeno europeu que apoiam as famílias como um clado. Como resultado, ele argumentou que a superfamília proposta Anoplotherioidea, composta por Anoplotheriidae e Xiphodontidae, como proposto por Alan W. Gentry e Hooker em 1988, é inválida devido à polifilia das linhagens na análise filogenética. No entanto, Xiphodontidae ainda foram encontrados como parte de um clado mais amplo com os outros três grupos. Anoplotherium e Diplobune compõem um clado de Anoplotheriidae por causa de suas características dentárias derivadas, apoiadas por serem os anoploteriídeos que apareceram mais tardiamente.[53][57]
Descrição
Crânio

Os Anoplotheriidae são caracterizados em parte por crânios de proporções baixas com focinhos alongados (o focinho se alinha com o topo do crânio no caso de Anoplotherium), e uma órbita craniana bem aberta.[58][48][45] Anoplotherium não possui processos ósseos e fossas lacrimais. Possui grandes processos paroccipitais e projeções mais curtas do processo pós-orbital do osso lacrimal.[59][60]
O crânio de Anoplotherium é estreito e alongado, com um osso pós-orbital constrito, indicando um desenvolvimento cerebral pobre. Apresenta cristas sagital e nucal [en] robustas, a primeira com elevações altas e emergindo de cristas pós-orbitais baixas, e a última com deslocamentos de elevação complicados. A parte de trás tem um forame magno circular e grandes côndilos occipitais. A parte inferior tem um palato alongado com superfícies glenoides e fortes processos pós-glenoides do osso esquamosal.[61]
Os ossos do crânio são robustos, com o osso esponjoso diploë sendo muito desenvolvido. A força do crânio é atribuída a músculos temporais maciços como parte de uma constituição corporal geral forte. O crânio tem uma sela túrcica rasa, uma fossa craniana em forma de pêra, ossos parietais extensos, osso esquamosal grande, osso occipital estreito e dois pequenos tubérculos occipitais para fixação muscular. Muitas características cranianas vistas em Anoplotherium também são encontradas no parente próximo Diplobune.[61][62]
Na região auditiva (incluindo os ossos temporais), o osso periótico do ouvido interno é extenso, sendo as aberturas do meato auditivo interno [en] e do canal facial [en] do osso temporal visíveis na área triangular inferior do osso periótico. A parte timpânica do osso temporal [en] está conectada parcialmente ao osso esquamosal, permanece separada do osso periótico e consiste em uma bula auditiva pequena, mas espessa (estrutura óssea oca da região auditiva), que se projeta por baixo da parte petrosa do osso temporal [en].[61]
Num fragmento de crânio de A. laurillardi com alvéolos de incisivos e caninos, o comprimento conhecido da região nasal é grande, medindo 38,1 mm.[45] A característica de nasais grandes é semelhante ao que foi observado num crânio de Diplobune secundaria, que são registados como maciços, alongados e conectados entre si e à maxila. Cyril Gagnaison e Jean-Jacques Leroux propuseram, no caso de D. secundaria, que a região nasal alongada suporta a presença de uma língua muito afilada, que, semelhante às girafas, pode tê-lo permitido puxar ramos de plantas.[63]
Anatomia do endocasto

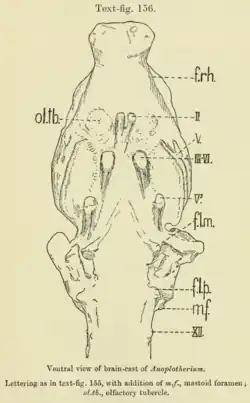
Em 1913, R.W. Palmer conduziu estudos sobre o molde do cérebro de um crânio de Anoplotherium commune, originário das fosfóritas de Quercy [en], das coleções do Museu Britânico (o endocasto está agora no Museu Nacional de História Natural, França, como o espécime BMNH 3753). O indivíduo em questão foi estimado a pesar 80 kg na sua morte, semelhante às lhamas existentes, pesando consideravelmente menos do que as estimativas típicas de um Anoplotherium adulto. O comprimento total do cérebro é inferior a 10 cm, e seu volume mede aproximadamente 230 ml.[61][64][45]
A forma do cérebro é naturalmente estreita e alongada.[61][12] O cerebelo e o cérebro estão ambos em posições altas em comparação com os ungulados modernos, que têm os hemisférios cerebrais localizados acima do cerebelo. Palmer notou que o cérebro era semelhante ao de Orycteropus afer. O cérebro altamente desenvolvido que permite um forte olfato em Anoplotherium o torna macrosmático (derivado no olfato), como também indicado pelos bulbos olfativos aumentados e pelo pequeno tamanho do neocórtex.[61] Tanto em Anoplotherium quanto em Diplobune, a fissura rinal divide o hemisfério cerebral horizontalmente e em metades iguais. O vérmis cerebelar do cerebelo é dividido quase igualmente pela fissura primária do cerebelo [en] (ou "fissura prima").[65]
Além disso, os bulbos olfativos são espessos, e os tubérculos olfativos [en] assumem a forma de elevações circulares lisas que são mais curvadas para trás do que no oricterope e são facilmente perceptíveis.[61] Em outro endocasto de Anoplotherium, os bulbos olfativos compõem 7,5% do volume total do cérebro, acima da média para artiodáctilos extintos e existentes.[12]
A área do neocórtex do cérebro, responsável pela percepção sensorial e outras funções cerebrais sensoriais, cobre 28% da área de superfície do endocasto de um A. commune de tamanho médio.[64] Outro endocasto, que pertence a Anoplotherium sp., mede 7173,92 mm² na superfície do cérebro, 4419,56 mm² na superfície do neopálio e 416,09 cm³ no volume endocraniano. Os dois primeiros dados, quando calculados juntos (superfície do neopálio/superfície do cérebro), compõem 61,6% da área de superfície neocortical total do cérebro, o que significa que Anoplotherium adulto tem medidas maciças de superfície cerebral e neocortical em comparação com a maioria dos artiodáctilos do Paleogeno, sendo a última medida igual ou menor que a dos artiodáctilos modernos.[12]
Anoplotherium e outros anoploteriídeos compartilham características de sulcos [en] geralmente alongados e paralelos no córtex cerebral, bem como um sulco vertical (cordial) correspondente ao sulco lateral. As fissuras na superfície da área central do cérebro mostram formações claras de um sulco lateral complexo (também conhecido como fissura de Sylvius) em um processo conhecido como operculização.[12] A operculização do cérebro dos anoploteriídeos é semelhante à de Anthracotheriidae, mas não indica qualquer relação filogenética próxima, o que significa que as semelhanças são um exemplo de evolução paralela. As medidas dos endocastos de Anoplotherium são maiores que as de outros artiodáctilos do Paleogeno em um estudo de 2015 de Ghislain Thiery e Stéphane Ducrocq.[66]
Dentição

Ao contrário da maioria dos gêneros de mamíferos fósseis, Anoplotherium é diagnosticado principalmente com base na morfologia pós-craniana em vez da morfologia dentária, mas possui diagnósticos baseados nesta última.[45] A fórmula dentária de Anoplotherium e outros anoploteriídeos é para um total de 44 dentes, consistente com a fórmula dentária primitiva para mamíferos placentários do Paleogeno inicial-médio.[58][67] Os anoploteriídeos têm pré-molares (P/p) e molares (M/m) selenodontes (forma de crista em crescente) ou bunoselenodontes (bunodonte e selenodonte), feitos para dietas de ramoneio de folhas. Os caninos (C/c) de Anoplotheriidae são, em geral, indiferenciados dos incisivos (I/i). Os pré-molares inferiores da família são perfurantes e alongados. Os molares superiores são de forma bunoselenodonte, enquanto os molares inferiores têm cúspides labiais selenodontes e cúspides linguais bunodontes (ou arredondadas). A subfamília Anoplotheriinae difere de Dacrytheriinae pelos pré-molares molariformes com paracônulos em forma de crescente e os molares inferiores que não possuem uma terceira cúspide entre o metacônido e o entocônido.[45]

Os molares superiores de Anoplotherium são caracterizados por contornos trapezoidais em vistas oclusais (ou vistas superiores do esmalte do dente), ectolofos em forma de W (cristas ou sulcos dos dentes molares superiores) e diferenças específicas nas cúspides. Mais especificamente, os molares superiores do gênero contêm cúspides protocônicas quase centrais e cônicas, alinhadas de perto com as cúspides do mesostilo, paracônulos cônicos que estão conectados ao parastilo por cristas posteriores, e parastilos e mesostilos comprimidos. Os molares inferiores do anoploteriídeo contêm as cúspides do paracônido e do metacônido, que têm separações pronunciadas por um vale entre elas.[58][45]
Vértebras e costelas

Anoplotherium tem um total de 7 vértebras cervicais para uma série de C1-C7, típica da maioria dos mamíferos. O atlas (C1) é semelhante ao de camelídeos como Lama em forma, bem como na posição dos "forames alares" em associação com as conexões das facetas articulares envolvendo o áxis (C2).[11][23] Um áxis que foi atribuído a A. commune (mas também possivelmente pertencente a seu parente próximo Diplobune secundaria) é alongado em comprimento e tem um processo espinhoso diminuído. As vértebras C3-C7 são análogas a Cainotherium. A vértebra C4 parece inclinada, o que sugere que o pescoço muda de orientação da vértebra C3 para a C4 como uma potencial curvatura na área frontal do pescoço, semelhante aos ursos modernos. Como resultado da morfologia das vértebras do pescoço, Anoplotherium provavelmente tinha uma posição inclinada e ascendente do pescoço.[23]
Anoplotherium também tinha 12 vértebras torácicas, 6 vértebras lombares e 3 vértebras sacrais. As vértebras lombares, especialmente L4-L6, contêm processos transversos que são largos, longos e apontam ligeiramente para a frente. As 3 vértebras sacrais são robustas e contêm apófises para fortes fixações à longa cauda. As vértebras do gênero anoploteriídeo são construídas para o movimento típico dos ungulados.[11][23]
O aspecto pós-craniano mais incomum de Anoplotherium em comparação com outros artiodáctilos é a cauda longa e espessa, que é composta por 22 vértebras caudais para um forte suporte muscular. As vértebras frontais tinham um processo bem pronunciado, e todas as vértebras, exceto as mais distais, têm arcos hemais.[23]
Como o calicotério Chalicotherium e ao contrário de outros mamíferos como os caprinos do gênero Ovis e Cainotherium, as costelas se curvam em áreas mais largas e seus tubérculos não se projetam tanto na direção dorsal. As costelas de Anoplotherium formam um tronco em forma de barril, o que significa que a caixa torácica é muito mais larga que a dos ruminantes modernos. As costelas geralmente se projetam para os lados devido às suas posições muito curvas, à posição do tubérculo e às vértebras torácicas que se projetam nos lados superiores.[23]
Membros
Anoplotherium tem membros curtos e acredita-se que tenha sido ungulígrado nas posições dos membros, com a maioria das espécies tendo três dedos tanto nos membros dianteiros quanto nos traseiros. A. commune se diferencia do semelhante A. latipes por seus dígitos didáctilos ("de dois dedos") em oposição aos tridáctilos ("de três dedos").[23][45][68]
Membros dianteiros

A escápula (ou omoplata) tem uma borda coracoide convexa e é semelhante à de Diplobune. Semelhante aos camelos (Camelus), a fossa supraespinhosa [en] é mais larga que a fossa infraespinhosa [en], mas os camelos têm escápulas mais estreitas, especialmente nas extremidades distais (posteriores) da fossa supraespinhosa. A espinha da escápula é robusta, espessa e aumenta gradualmente em altura distalmente até atingir a borda das cavidades glenoides, como nos camelos, mas ao contrário da maioria dos outros artiodáctilos modernos. O processo coracoide (normalmente semelhante a uma pequena estrutura em forma de gancho) é reduzido a um nódulo rombo que se projeta apenas ligeiramente. A larga fossa supraespinhosa e a borda coracoide amplamente curvada das escápulas de Anoplotherium são diferentes de Cainotherium e Merycoidodon porque Anoplotherium não compartilha nem a forma triangular das omoplatas nem as fossas supraespinhosas estreitas.[23]
A morfologia do cotovelo de Anoplotherium, baseada nas estruturas e articulações dos ossos do cotovelo como o úmero, rádio e ulna, mostra evidências de adaptações para mover o cotovelo para cima e para baixo em rotações de supinação-pronação de no máximo 13°. Um cotovelo totalmente estendido poderia formar um ângulo entre a ulna e o úmero que mede aproximadamente 135°, indicando alta flexibilidade em comparação com outros artiodáctilos, incluindo os cotovelos já semiflexíveis de Cainotherium.[23]
Semelhante na morfologia do pulso aos porcos do gênero Sus, os cascos de Anoplotherium se abrem em ~16° quando para baixo, apoiado pela morfologia das pegadas. O pulso pode ter sido capaz de girar para cima e para baixo, mas apenas em um grau limitado e longe das morfologias de pulso flexíveis dos primatas, sugerindo que a adaptação não era uma característica principal do gênero de artiodáctilo, mas o resultado de recuperar uma característica primitiva.[23]
O carpo consiste no escafoide, semilunar, piramidal e pisiforme em sua primeira fileira e no trapézio, trapezoide, capitato e hamato em sua segunda. Anoplotherium tem quatro ossos dos dedos, mas os do dígito V e, no caso de A. commune, do dígito II são pouco desenvolvidos.[69] O segundo dedo (dígito II) de Anoplotherium não tem capacidade de rotação ou movimentos flexíveis, o que significa que não desempenha nenhum papel semelhante ao polegar, como nos primatas ou no panda-gigante.[23]
Membros traseiros

O ílio, parte do osso do quadril da bacia maior, é largo e tem uma crista ilíaca firmemente arredondada que se encontra com a borda inferior côncava em um ângulo agudo. O ílio de Anoplotherium pode ser diferenciado de Palaeotherium pelo corpo ilíaco mais curto, o ísquio (a área inferior e posterior do osso do quadril) mais longo e uma borda posterior da pelve mais reta que resulta em uma sínfise púbica mais longa. A região da fossa acetabular [en] da superfície do acetábulo da pelve é grande, com sua incisura acetabular [en] em uma posição posterior, semelhante à de Chalicotherium.[23]
O fêmur é maior que a tíbia, tem apenas dois trocânteres, semelhante a outros artiodáctilos basais, tem um espaço estreito entre sua cabeça femoral [en] e o trocanter maior [en], e tem um colo femoral [en] longo. A fossa trocantérica [en], uma cavidade na superfície do trocanter maior, é larga em profundidade e estreita em forma, aprofundando-se nas laterais. A tíbia é robusta, suportando fortemente as fixações musculares com base em suas cristas e processos. A extremidade distal da fíbula mais a proeminência do maléolo medial da tíbia envolvem a área central do astrágalo para evitar que ele se mova lateralmente.[23]
Os anoploteriídeos com fósseis pós-cranianos conhecidos têm astrágalos (ou talus ou osso do tornozelo) proporcionalmente largos, robustos e oblíquos, diferindo amplamente de outros artiodáctilos. A. latipes difere de A. commune em parte por morfologias das facetas e fossas do astrágalo e um calcâneo (osso do calcanhar) mais curto e mais robusto.[23][52] Os astrágalos dos anoplotérios compartilham níveis de elevações e posições de facetas específicas com os mericoidodontes que nenhum artiodáctilo moderno compartilha, possivelmente um exemplo de evolução convergente.[70][71] A faceta medial (sustentacular) de Anoplotherium e Diplobune é côncava, contrastando com a faceta plana a ligeiramente convexa de Dacrytherium [en].
O tarso consiste no navicular, três ossos cuneiformes e um osso cuboide. O pé de A. commune consistia em dois dedos, como indicado pelos ossos cuneiformes mais externos e médios relativamente pequenos.[69]
Pegadas
Pegadas de grande porte do sul da França e do norte da Espanha que datam do Eoceno Superior[72] podem ter sido de Anoplotherium. O icnogênero é chamado de Anoplotheriipus e foi descrito pela primeira vez no departamento de Gard, na França, por Paul Ellenberger em 1980. A derivação do nome do gênero refere-se ao fato de o icnotáxon ser o mais próximo em afinidade com Anoplotheriidae. O icnogênero é diagnosticado como pertencente a um artiodáctilo muito grande, a área do autópode excedendo a de A. commune em ~33%, a posição subparalela dos dois cascos e a área posterior da sola do pé sendo tão larga transversalmente quanto a área anterior da sola do pé.[73] Anoplotheriipus tem formato redondo a retangular com impressões de dígitos fendidos largas e pronunciadas anteriormente que se assemelham a rastros de camelo mal preservados.[74] O icnogênero de artiodáctilo semelhante, Diplartiopus, difere dele pelo paralelismo dos dois dedos, que são mais alongados.[75]
A espécie-tipo é Anoplotheriipus lavocati, que Ellenberger nomeou em homenagem ao paleontólogo René Lavocat [en] e considerou a "mais majestosa" das três icnoespécies devido à mobilidade específica exibida pelos metatarsos. Mede de 170 mm a 180 mm de comprimento e 120 mm de largura, tem formato robusto e mede 12° na divergência dos dedos. Os dois dedos são quase iguais em comprimento e, no mínimo, medem 115 mm sem os ossos metatarsais serem levados em conta e 225 mm com os metatarsos. As medidas são consideravelmente maiores que as medidas típicas dos dedos de A. commune, que são 85 mm sem os metatarsos e 170 mm com.[73]
Anoplotheriipus similicommunis, derivando na etimologia da espécie de "similis" (semelhante em latim) e A. commune, é semelhante à icnoespécie-tipo, mas é menor, correspondendo mais diretamente às medidas típicas de pé de A. commune por seu comprimento de 140 mm e largura de 105 mm. O ângulo de divergência entre os dois dedos principais é de 10°, e os comprimentos mínimos dos dedos são 90 mm sem os metatarsos e 180 mm com.[73]
Anoplotheriipus compactus é a terceira icnoespécie, que na etimologia da espécie deriva da palavra latina "compactus", que significa "compacto" em português, devido ao autópode curto e arredondado. Tem um diagnóstico menos definitivo em comparação com os outros dois icnotáxons, mas é semelhante em tamanho a A. similicommunis e tem uma sola do pé quase circular para suportar dedos ligeiramente mais curtos. Seu comprimento é de 120 mm, enquanto a largura é de 100 mm, e os comprimentos dos dedos medem 70 mm - 80 mm sem os metatarsos e 140 mm - 150 mm com. As pegadas podem ter sido produzidas por A. latipes, embora a resposta ainda seja incerta.[73]
Tamanho

As espécies de Anoplotherium eram particularmente grandes no Eoceno Superior, atingindo tamanhos incomuns para a maioria dos grupos de artiodáctilos do Paleogeno. As estimativas de grande porte começaram em 1995, quando Martinez e Sudre fizeram estimativas de peso de artiodáctilos do Paleogeno com base nas dimensões de seus astrágalos e dentes M1. Os astrágalos são ossos comuns em conjuntos fósseis devido à sua vulnerabilidade reduzida à fragmentação como resultado de sua forma robusta e estrutura compacta, explicando sua escolha para o uso. As duas medidas para A. commune produziram resultados diferentes, com o M1 dando a massa corporal de 312,075 kg e o astrágalo produzindo 265,967 kg. Essas estimativas são muito maiores que as da maioria dos outros artiodáctilos do Paleogeno no estudo, embora os pesquisadores tenham apontado que as medidas do M1 poderiam ser superestimadas em comparação com a estimativa do astrágalo.[52]
Em 2014, Takehisa Tsubamoto reexaminou a relação entre o tamanho do astrágalo e a massa corporal estimada com base em estudos extensivos de mamíferos terrestres existentes, reaplicando os métodos a artiodáctilos do Paleogeno previamente testados por Sudre e Martinez. O pesquisador usou medidas lineares e seus produtos com fatores de correção ajustados. Os recálculos resultaram em estimativas um pouco mais baixas em comparação com os resultados de 1995 (com exceção de Diplobune minor, que tem uma proporção de astrágalo mais curta do que a maioria dos outros artiodáctilos), exibidas no gráfico abaixo:[76]
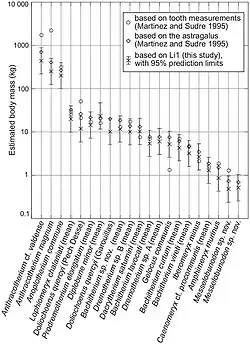
Em 2022, Weppe calculou a massa corporal de A. commune, resultando em 360 kg.[53] Em 2023, Ainara Badiola et al. estimaram que o peso de Anoplotherium varia entre 115 kg e 271 kg. Em seus cálculos, A. laurillardi era o anoploteriídeo menor que pesava em média 157 kg. A. latipes era maior e tem uma estimativa de peso médio de 229 kg, e A. commune tem as estimativas de peso mais pesadas, de 271 kg.[45]
Em 2007, Hooker fez estimativas de tamanho de A. latipes com base em um esqueleto incompleto de um indivíduo imaturo do Membro Hamstead da formação Bouldnor [en] na ilha de Wight, Reino Unido. O indivíduo reconstruído do nível 3 de Hamstead deu medidas de tamanho de 2 m de comprimento de cabeça e corpo. O úmero do indivíduo imaturo de Anoplotherium mede 330 mm de comprimento, então os úmeros de indivíduos maduros podem ter medido cerca de 410 mm de comprimento. Como resultado, A. latipes adulto pode ter medido 2,5 m de comprimento de cabeça e corpo e 1,25 m de altura no ombro. Ao ficar em pé bipedalmente em seus membros traseiros com as costas, pescoço e cabeça em um ângulo de cerca de 15°, o indivíduo do nível 3 de Hamstead pode ter alcançado 2,5 m ao se alimentar, enquanto indivíduos mais maduros de A. latipes podem ter ficado um pouco acima de 3 m.[23]
Paleobiologia

Desde 2007, acredita-se que Anoplotherium fosse um quadrúpede que poderia ter ficado em pé sobre as patas traseiras como um herbívoro bípede, graças à pélvis forte, à cauda longa e robusta para equilíbrio e às pernas traseiras abertas. As adaptações bípedes mostram algum exemplo de convergência com outros animais, como os calicotérios, vários gêneros de preguiças-gigantes, pandas-gigantes (Ailuropoda melanoleuca), gorilas (Gorilla) e o gerenuk (Litocranius walleri). De resto, a forma geral do corpo parece assemelhar-se à de Canidae. Como resultado da curvatura das vértebras cervicais C3-C4, o pescoço e a cabeça poderiam ter mantido orientações horizontais enquanto estivessem em pé bipedalmente. Os membros anteriores poderiam ter se estendido horizontalmente além do focinho enquanto o indivíduo estivesse em pé bipedalmente, embora não pudesse ter alcançado para cima e não tivesse garras ou órgãos preênseis na mão, ao contrário de Chalicotherium. Portanto, os antebraços podem não ter sido usados para rasgar e arrancar plantas, mas como apoio bípede. Pode ter se alimentado enquanto estava em pé em um ângulo mais acentuado, mais comparável ao gerenuk do que Chalicotherium.[23]
Seu grande tamanho e capacidade de se alimentar bipedalmente podem ter dado a Anoplotherium poucas fontes de competição terrestre, exceto por Palaeotherium magnum, um paleotério de grande porte com um pescoço longo que pode ter atingido 240,3 kg de massa corporal.[23][77] A subespécie P. magnum magnum teria atingido pouco mais de 2 m de altura de alimentação em postura quadrúpede, e não há evidências de qualquer adaptação bípede em paleotérios.[23] Anoplotherium provavelmente se engajou em graus de partição de nicho com os paleotérios do Eoceno Superior e Diplobune. Embora todos fossem herbívoros folívoros, os paleotérios Plagiolophus [en] e Palaeotherium podem ter tido pequenos graus de frugivoria, enquanto Diplobune era provavelmente adaptado ao arborealismo.[78][79][68] O quão bem adaptado o Anoplotherium era a folhas abrasivas e a condições mais secas, mas ainda subúmidas, no Eoceno Superior não é bem conhecido e requer pesquisas futuras na dentição para obter respostas.[23]

Hooker propôs a possibilidade de que os didáctilos A. commune e A. latipes possam ter sido dimorfos sexuais da mesma espécie (na qual A. latipes seria um sinônimo de A. commune). Existem poucas diferenças consistentes na morfologia dentária entre as duas espécies, com quaisquer pequenas diferenças podendo ser atribuídas a variações individuais. As diferenças no número de dedos entre as espécies podem ter refletido o fato de A. latipes ser tridáctilo e A. commune ser didáctilo. O paleontólogo explicou que, embora não haja evidências de que o dedo extra tocasse o chão enquanto o indivíduo caminhava, o dedo extra de A. latipes pode ter servido como equilíbrio adicional ao se alimentar bipedalmente.[23]
O terceiro dedo também pode ter servido como parte de disputas em competição intraespecífica entre indivíduos machos. No entanto, ele observou que, apesar da aparente "vantagem" de A. latipes na alimentação bípede, não há evidências de diferenças sexuais nos comportamentos ou preferências alimentares. Além disso, ambas as espécies são encontradas nas mesmas localidades de Bouldnor no Reino Unido, além de La Débruge e Montmartre na França, e embora A. latipes seja mais comum em La Débruge do que em Montmartre, isso pode ser resultado de fatores comportamentais e/ou tafonômicos.[23][45] Grégoire Métais expressou não estar convencido de que o terceiro dedo de A. latipes seja uma adaptação sexualmente dimórfica para a alimentação bípede, sugerindo, em vez disso, que eles eram usados em disputas entre machos, se A. latipes e A. commune fossem dimorfos sexuais.[68]
Algumas evidências das morfologias de Anoplotherium foram criticadas por algumas fontes. Em seu estudo da morfologia do gerenuk que permite a bipedia, os pesquisadores Matt Cartmill e Kaye Brown argumentaram que várias características pós-cranianas que supostamente eram adaptações de Litocranius e outros gêneros bípedes não distinguem o gerenuk de outros bovídeos.[80] Ciaran Clark et al. (incluindo J.J. Hooker) descobriram, a partir de tomografia computadorizada, que a ideia de Anoplotherium ser um navegador bípede facultativo [en] não era apoiada pela arquitetura trabecular da área proximal do fêmur. Isso pode ter sido resultado de dados de baixa qualidade das microtomografias e do tamanho menor da amostra, e dados de micro-CT de maior contraste podem fornecer melhores respostas sobre informações posturais.[81]
Os padrões de pegadas de Anoplotheriipus sugerem que os Anoplotherium caminhavam em velocidades de movimento muito semelhantes entre si. Com base nos agrupamentos do icnotáxon de pegadas na localidade de Fondota, no município de Abiego, na Espanha, Anoplotherium pode ter caminhado comumente em pequenos grupos, o que pode implicar algum comportamento gregário (ou sociável).[82]
Paleoecologia
Europa pré-Grande Coupure
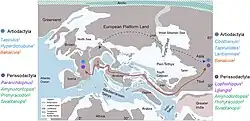
Durante a maior parte do Eoceno, prevaleceu um clima de estufa com ambientes húmidos e tropicais, com precipitações consistentemente altas. As ordens de mamíferos modernos, incluindo Perissodactyla, Artiodactyla e Primatas (ou a subordem Euprimates), já haviam aparecido no Eoceno Inferior, diversificando-se rapidamente e desenvolvendo dentições especializadas para a folivoria. As formas onívoras, na sua maioria, ou mudaram para dietas folívoras ou extinguiram-se no Eoceno Médio (47-37 milhões de anos atrás), juntamente com os "condilartros" arcaicos. No Eoceno Superior (aprox. 37-33 Ma), a maioria das dentições de formas unguladas mudou de cúspides bunodontes (ou arredondadas) para cristas cortantes (ou seja, lofos) para dietas folívoras.[83][84]
As conexões terrestres entre a Europa Ocidental e a América do Norte foram interrompidas por volta de 53 Ma. Desde o Eoceno Inferior até o evento de extinção Grande Coupure (56-33,9 Ma), a Eurásia Ocidental estava separada em três massas de terra: Europa Ocidental (um arquipélago), Balcanatolia (entre o Mar de Paratethys ao norte e o Oceano Neotethys ao sul) e Eurásia Oriental.[47] As faunas de mamíferos holárticos da Europa Ocidental estavam, portanto, em grande parte isoladas de outras massas de terra, incluindo a Gronelândia, África e Eurásia Oriental, permitindo o desenvolvimento de endemismo.[84] Portanto, os mamíferos europeus do Eoceno Superior (MP17-MP20 das zonas do Paleogeno de mamíferos) eram, na sua maioria, descendentes de grupos endêmicos do Eoceno Médio.[85]
O aparecimento de anoploteriíneos derivados por MP18 ocorreu muito depois da extinção da família de perissodáctilos endêmica europeia Lophiodontidae [en] em MP16, incluindo o maior lofiodonte, Lophiodon [en] lautricense, provavelmente resultado de uma mudança de ambientes húmidos e altamente tropicais para florestas mais secas e temperadas com áreas abertas e vegetação mais abrasiva. As faunas herbívoras sobreviventes mudaram suas dentições e estratégias alimentares de acordo para se adaptarem.[86][87] Os ambientes ainda eram subúmidos e cheios de florestas subtropicais perenes, no entanto. Palaeotheriidae era o único grupo de perissodáctilos europeu remanescente, e os artiodáctilos frugívoros-folívoros ou puramente folívoros tornaram-se o grupo dominante na Europa Ocidental.[88][89] MP16 também marcou os últimos aparecimentos da maioria dos crocodilomorfos europeus, dos quais o aligatoroide Diplocynodon [en] foi o único sobrevivente devido a aparentemente se adaptar ao declínio geral dos climas tropicais do Eoceno Superior.[90][91][92]
Eoceno Superior
Após uma lacuna considerável nos fósseis de anoploteriíneos em MP17a e MP17b, os anoploteriíneos derivados Anoplotherium e Diplobune fizeram suas primeiras aparições conhecidas na unidade MP18.[50] Eles eram exclusivos do arquipélago da Europa Ocidental, mas suas origens exatas e rotas de dispersão são desconhecidas. Naquela época, Anoplotherium e Diplobune viviam na Europa Central (então uma ilha) e na Península Ibérica, sendo que apenas o primeiro gênero se dispersou mais tarde para o sul da Inglaterra por MP19, devido à aparente falta de barreiras oceânicas.[45][23]
Anoplotherium coexistiu com uma grande diversidade de artiodáctilos na Europa Ocidental por MP18, variando desde os mais difundidos Dichobunidae, Tapirulidae e Anthracotheriidae até muitas outras famílias endêmicas, como Xiphodontidae, Choeropotamidae [en], Cebochoeridae, Amphimerycidae [en] e Cainotheriidae.[48][54][93][94] Anoplotherium também coexistiu com os Palaeotheriidae, a família de perissodáctilos remanescente da Europa Ocidental.[85] Grupos europeus do Eoceno Superior do clado Ferae representavam predominantemente Hyaenodonta [en] (Hyaenodontinae [en], Hyainailourinae [en] e Proviverrinae [en]), mas também continham Carnivoramorpha (Miacidae) e Carnivora (Amphicyonidae de pequeno porte).[88] Outros grupos de mamíferos presentes no Eoceno Superior da Europa Ocidental representavam os leptictídeos (Pseudorhyncocyonidae),[95] primatas (Adapoidea e Omomyoidea),[96] eulipotiflanos (Nyctitheriidae),[97] quirópteros,[84] herpetoteriídeos [en],[98] apatotérios [en],[99] e roedores endêmicos (Pseudosciuridae, Theridomyidae e Gliridae).[100] O aligatoroide Diplocynodon, presente apenas na Europa desde o Paleoceno superior, coexistiu também com as faunas pré-Grande Coupure.[101] Além de cobras, sapos e salamandrídeos, uma rica assembleia de lagartos também é conhecida na Europa Ocidental de MP16-MP20, representando Iguanidae, Lacertidae, Gekkonidae, Agamidae, Scincidae, Helodermatidae e Varanoidea [en].[102]
Na localidade MP18 de Zambrana, na Espanha, foram encontrados restos de A. laurillardi e A. sp. com grupos indeterminados de sapos e escamados, o aligatoroide Diplocynodon, o herpetoteriídeo Peratherium [en], roedores (Theridomys, Elfomys, Pseudoltinomys [en], Remys), o omomiídeo Microchoerus [en], os carnivoraformes Quercygale [en] e Paramiacis [en], o dicobunídeo Dichobune, os xifodontes Xiphodon e Haplomeryx, e os paleotérios (Palaeotherium, Leptolophus, Iberolophus, Pachynolophus, Paranchilophus).[103]
Como parte de uma massa de terra separada na época, La Débruge, na França, datada de MP18, rendeu faunas ligeiramente diferentes que coexistiram com A. commune, A. latipes e A. laurillardi, nomeadamente o herpetoteriídeo Peratherium, roedores (Blainvillimys, Theridomys, Plesiarctomys, Glamys), hienodontes (Hyaenodon e Pterodon [en]), o anficíonídeo Cynodictis [en], paleotérios (Plagiolophus, Anchilophus, Palaeotherium), o dicobunídeo Dichobune, o queropotamídeo Choeropotamus [en], os ceboquerídeos Cebochoerus [en] e Acotherulum, os anoploteriídeos Dacrytherium e Diplobune, o tapirulídeo Tapirulus, os xifodontes Xiphodon e Dichodon [en], o cainotério Oxacron, o anfimerídeo Amphimeryx e o antracotério Elomeryx.[104]
Extinção

O evento Grande Coupure, durante o final do Eoceno até o início do Oligoceno (MP20-MP21), é uma das maiores e mais abruptas mudanças faunísticas do Cenozoico da Europa Ocidental e coincide com eventos de forçamento climático de climas mais frios e sazonais.[105] O evento levou à extinção de 60% das linhagens de mamíferos da Europa Ocidental, que foram subsequentemente substituídas por imigrantes asiáticos.[106][107][108] A Grande Coupure é frequentemente datada diretamente no limite Eoceno-Oligoceno, a 33,9 Ma, embora alguns estimem que o evento tenha começado um pouco mais tarde, a 33,6-33,4 Ma.[109][110] O evento ocorreu durante ou após a transição Eoceno-Oligoceno, uma mudança abrupta de um mundo de estufa quente que caracterizou grande parte do Paleogeno para um mundo de casa fria/casa de gelo a partir do Oligoceno Inferior. A queda maciça de temperaturas resulta da primeira grande expansão das camadas de gelo da Antártida, que causou diminuições drásticas de pCO2 e uma queda estimada de ~70 m no nível do mar.[111]
Muitos paleontólogos concordam que a glaciação e as consequentes quedas no nível do mar permitiram um aumento das migrações entre a Balcanatolia e a Europa Ocidental. O estreito de Turgai [en], que antes separava grande parte da Europa da Ásia, é frequentemente proposto como a principal barreira marítima europeia antes da Grande Coupure, mas alguns pesquisadores contestaram recentemente essa percepção, argumentando que ele já havia recuado completamente há 37 Ma, muito antes da transição Eoceno-Oligoceno. Em 2022, Alexis Licht et al. sugeriram que a Grande Coupure poderia possivelmente ter sido síncrona com a glaciação Oi-1 (33,5 Ma), que registra um declínio no CO2 atmosférico, impulsionando a glaciação antártica que já havia começado na transição Eoceno-Oligoceno.[47][112]
O evento Grande Coupure também marcou uma grande mudança faunística, com as chegadas de antracotérios posteriores, entelodontes, ruminantes (Gelocidae, Lophiomerycidae), rinocerotoides (Rhinocerotidae, Amynodontidae, Eggysodontidae [en]), carnívoros (Amphicyonidae posteriores, Amphicynodontidae, Nimravidae e Ursidae), roedores da Eurásia Oriental (Eomyidae [en], Cricetidae e Castoridae) e eulipotiflanos (Erinaceidae).[113][114][106][115]
A transição Eoceno-Oligoceno da Europa Ocidental, como resultado das condições climáticas globais, é marcada por uma transição de florestas tropicais e subtropicais para habitats mais abertos, temperados ou mistos decíduos, com adaptações a uma maior sazonalidade. Embora Anoplotherium não tenha durado muito no início do Oligoceno, há divergências sobre se ele sobreviveu à Grande Coupure ou se extinguiu no evento.[89][48] Embora as evidências apontem para a extirpação de Anoplotherium de áreas como a França e o Reino Unido pela Grande Coupure (últimas ocorrências em MP20),[104][106][53] a percepção é complicada pela aparente sobrevivência final de A. commune na localidade MP21 de Möhren 19, no sul da Alemanha (a borda da Europa Ocidental), juntamente com Palaeotherium medium e Diplobune quercyi (localidades ligeiramente mais jovens indicam suas extinções e substituições por imigrantes da Grande Coupure, como o antracotério Anthracotherium e o rinocerotídeo Epiaceratherium [en]).[116]
Hooker apontou que localidades como Möhren 19 abrangem tempos anteriores, onde as faunas endêmicas sobreviventes são acompanhadas por alguns imigrantes da Grande Coupure, mas ainda não foram unidas por certos imigrantes, como Anthracotherium. Além disso, os endêmicos sobreviventes da localidade estão ausentes de outras áreas datadas de MP21. Portanto, ele argumentou que certas localidades MP21 mais antigas com faunas endêmicas sobreviventes preenchem a longa lacuna entre o mais jovem membro Inferior de Hamstead, pré-Grande Coupure, e o mais jovem membro superior de Hamstead, pós-Grande Coupure, dentro da formação Bouldnor. Essa interpretação, explicou Hooker, significa que as localidades representaram momentos muito breves de sobrevivência de faunas endêmicas durante a Grande Coupure, apoiando assim a ideia de um evento de extinção e imigração faunística importante e rápido, incluindo a extinção de Anoplotherium no evento.[106][117]
As extinções da maioria dos artiodáctilos endêmicos, incluindo Anoplotherium, foram atribuídas à competição com faunas imigrantes, mudanças ambientais decorrentes do resfriamento climático, ou alguma combinação dos dois.[109] Sarah C. Joomun et al. determinaram que certas faunas podem ter chegado mais tarde e, portanto, podem não ter desempenhado um papel nas extinções. Eles concluíram que a mudança climática, que levou ao aumento da sazonalidade e a mudanças na disponibilidade de alimentos vegetais, fez com que os artiodáctilos se tornassem incapazes de se adaptar às grandes mudanças e se extinguissem.[118] Weppe fez argumentos semelhantes em relação à mudança climática como a principal causa do evento de extinção Grande Coupure, argumentando que os climas mais frios deslocaram os ambientes subtropicais anteriormente estáveis da Europa Ocidental e causaram um colapso na comunidade de artiodáctilos, que, após suas extinções, deixaram nichos ecológicos vazios que foram passivamente preenchidos por faunas imigrantes.[53]
Notas
- ↑ O latim commune é a forma neutra de communis, que se traduz em português como 'comum'.
Referências
- ↑ Cuvier, Georges (1804). «Suite des Recherches: Sur les espèces d'animaux dont proviennent les os fossiles répandus dans la pierre à plâtre des environs de Paris.». Annales du Muséum National d'Histoire Naturelle, Paris (em francês). 3: 364–387. Consultado em 30 de agosto de 2023. Cópia arquivada em 27 de julho de 2023
- ↑ Rudwick, Martin J. S. (2022). «Georges Cuvier's appeal for international collaboration, 1800». History of Geology. 46 (1): 117–125. doi:10.18814/epiiugs/2022/022002

- ↑ Roberts, George (1839). An etymological and explanatory dictionary of the terms and language of geology (em inglês). London: Longman, Orme, Brown, Green, & Longmans. p. 8. Consultado em 29 de Dezembro de 2021
- ↑ Cuvier, Georges (1804). «Suite des Recherches: Suite de recherches sur les os fossiles de la pierre à plâtre des environs de Paris. Troisième mémoire. Restitution des pieds. Première section. Restitution des différens pieds de derrière.». Annales du Muséum National d'Histoire Naturelle, Paris (em francês). 3: 442–472. Consultado em 30 de agosto de 2023. Cópia arquivada em 27 de julho de 2023
- ↑ Cuvier, Georges (1805). «Troisième mémoire. Deuxième section. Restitution des différens pieds de devant.». Annales du Muséum National d'Histoire Naturelle, Paris (em francês). 6: 253–283. Consultado em 30 de agosto de 2023. Cópia arquivada em 10 de novembro de 2012
- ↑ Cuvier, Georges (1807). «Suite des recherches sur les os fossiles des environs de Paris. Troisième mémoire, troisième section, les phalanges. Quatrième mémoire sur les os des extrémités, première section, les os longs des extrémités postérieures.». Annales du Muséum d'Histoire Naturelle. 9: 10–44. Consultado em 30 de agosto de 2023. Cópia arquivada em 2 de setembro de 2023
- ↑ a b Cuvier, Georges (1807). «Suite des recherches sur les os fossiles des environs de Paris. Ve mémoire, IIe section, description de deux squelettes presque entiers d'Anoplotherium commune.». Annales du Muséum d'Histoire Naturelle (em francês). 9: 272–282. Consultado em 30 de agosto de 2023. Cópia arquivada em 2 de setembro de 2023
- ↑ Belhoste, Bruno (2017). «Chapter 10: From Quarry to Paper. Cuvier's Three Epistemological Cultures». In: Chemla, Karine; Keller, Evelyn Fox. Cultures without Culturalism: The Making of Scientific Knowledge. [S.l.]: Duke University Press. pp. 250–277
- ↑ Wallace, David Rains (2004). «Chapter 1: Pachyderms in the Catacombs». Beasts of Eden: Walking Whales, Dawn Horses, and Other Enigmas of Mammal Evolution. [S.l.]: University of California Press. pp. 1–13
- ↑ Allemand, Rémi (2017). Endocranial microtomographic study of marine reptiles (Plesiosauria and Mosasauroidea) from the Turonian (Late Cretaceous) of Morocco: palaeobiological and behavioral implications (PhD). Museu Nacional de História Natural, França
- ↑ a b c d Cuvier, Georges (1822). Recherches sur les ossemens fossiles, où l'on rétablit les caractères de plusieurs animaux dont les révolutions du globe ont détruit les espèces. 3. [S.l.]: G. Dufour and E. d'Ocagne. Consultado em 30 de agosto de 2023. Cópia arquivada em 19 de agosto de 2023
- ↑ a b c d e f Orliac, Maeva J.; Maugoust, Jacob; Balcarcel, Ana; Gilissen, Emmanuel (2023). «Paleoneurology of Artiodactyla, an Overview of the Evolution of the Artiodactyl Brain» (PDF). In: Dozo; Paulina-Carabajal, Ariana; Macrini, Thomas E.; Walsh, Stig. Paleoneurology of Amniotes. [S.l.]: Springer Cham. pp. 507–555. ISBN 978-3-031-13982-6. doi:10.1007/978-3-031-13983-3_13. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 29 de agosto de 2023
- ↑ Moodie, Roy Lee (1917). «Studies in Paleopathology. I. General Consideration of Evidence of Pathological Conditions Found among Fossil Animals». Annals of Medical History. 1 (4): 374–393. PMC 7927727. PMID 33943144
- ↑ Diéguez, Carmen; Isidro, Albert; Malgosa, Assumpció (1996). «An introduction to zoo-paleopathology and an update on fossil phyto-paleopathology from Spain». Journal of Paleopathology. 8 (3): 133–142
- ↑ Thomas, Richard (2012). «Chapter 66: NonHuman Paleopathology». In: Buikstra, Jane; Roberts, Charlotte. The Global History of Paleopathology: Pioneers and Prospects. [S.l.]: Oxford University Press. pp. 652–664. doi:10.1093/acprof:osobl/9780195389807.003.0066
- ↑ Cuvier, Geoges (1812). «Résumé général et rétablissement des Squelettes des diverses espèces». Recherches sur les ossemens fossiles de quadrupèdes: où l'on rétablit les caractères de plusieurs espèces d'animaux que les révolutions du globe paroissent avoir détruites (em francês). 3. [S.l.]: Chez Deterville. Consultado em 30 de agosto de 2023. Cópia arquivada em 31 de julho de 2023
- ↑ Rudwick, Martin J. S. (1997). «Chapter 6: The Animals from the Gypsum Beds around Paris». Georges Cuvier, Fossil Bones, and Geological Catastrophes: New Translations and Interpretations of the Primary Texts. [S.l.]: University of Chicago Press
- ↑ Manucci, Fabio; Romano, Marco (2022). «Reviewing the iconography and the central role of 'paleoart': four centuries of geo-palaeontological art». Historical Biology. 35 (1): 1–48. doi:10.1080/08912963.2021.2017919
- ↑ Rudwick, Martin J.S. (1992). «Chapter 2: Keyholes into the Past». Scenes from Deep Time: Early Pictorial Representations of the Prehistoric World. [S.l.: s.n.] pp. 27–58
- ↑ a b Gervais, Paul (1848–1852). «Note sur le genre Eurytherium, suivie d'une liste comparative des Mamifères observés dans les hassins de Paris et d'Apt, et de remarques sur les Ongulés observés en France.». Zoologie et paléontologie françaises (animaux vertébrés): ou nouvelles recherches sur les animaux vivants et fossiles de la France. 2. [S.l.]: Arthus Bertrand. Consultado em 30 de agosto de 2023. Cópia arquivada em 4 de agosto de 2023
- ↑ a b Schlosser, Max (1883). «Uebersicht der bekannten Anoplotherien und Diplobunen nebst Erläuterung der Beziehungen zwischen Anoplotherium und anderen Säugethierfamilien.». Neues Jahrbuch für Mineralogie, Geologie und Palaeontologie, Abhandlungen. 2. Consultado em 30 de agosto de 2023. Cópia arquivada em 4 de agosto de 2023
- ↑ Dor, M. (1938). «Sur la biologie de l'Anoplotherium (L'Anoplotherium était-il aquatique?)». Mammalia. 2: 43–48
- ↑ a b c d e f g h i j k l m n o p q r s t u v w x y Hooker, Jerry J. (2007). «Bipedal browsing adaptations of the unusual Late Eocene–earliest Oligocene tylopod Anoplotherium (Artiodactyla, Mammalia)». Zoological Journal of the Linnean Society. 151 (3): 609–659. doi:10.1111/j.1096-3642.2007.00352.x
- ↑ Davydenko, Svitozar; Gol’din, Pavel; Bosselaers, Mark; Vahldiek, Bernd; Vliet, Henk Jan van (2023). «Gross and microscopic anatomy of a tibia tentatively attributed to a cetacean from the Middle Eocene of Europe, with a note on the artiodactyl Anoplotherium and on the perissodactyl Lophiodon». Paläontologische Zeitschrift. 97 (3): 627–652. Bibcode:2023PalZ...97..627D. doi:10.1007/s12542-023-00653-x
- ↑ Phillips, Samuel; Shenton, Francis Kingston John (1860). Guide to the Crystal Palace and park. [S.l.: s.n.] Consultado em 30 de agosto de 2023. Cópia arquivada em 29 de agosto de 2023
- ↑ a b Witton, Mark P.; Michel, Ellinor (2022). «Chapter 4: The sculptures: mammals». The Art and Science of the Crystal Palace Dinosaurs. [S.l.]: The Crowood Press. pp. 68–91
- ↑ Pomel, Auguste (1848). «Recherches sur les caractères et les rapports entre eux des divers genres vivants et fossiles des Mammifères ongulés». Comptes rendus hebdomadaires des séances de l'Académie des sciences. 26: 686–688
- ↑ Gervais, Paul (1848–1852). «Diverses espèces d'Ongulés fossiles.». Zoologie et paléontologie françaises (animaux vertébrés): ou nouvelles recherches sur les animaux vivants et fossiles de la France. 2. [S.l.]: Arthus Bertrand. Consultado em 30 de agosto de 2023. Cópia arquivada em 4 de agosto de 2023
- ↑ a b c d Lydekker, Richard (1885). Catalogue of the fossil Mammalia in the British museum, (Natural History): Part II. Containing the Order Ungulata, Suborder Artiodactyla. [S.l.]: Order of the Trustees, London. Consultado em 30 de agosto de 2023. Cópia arquivada em 2 de agosto de 2023
- ↑ Saint-Hilaire, Étienne Geoffroy (1833). «Considérations sur ossements fossiles, la plus inconnus, trouvés et observés dans le l'Auvergne». Revue encyclopédique, ou Analyse raisonnée des productions les plus remarquables. 59. [S.l.]: Bureau Central de la Revue Encyclopédique. Consultado em 19 de setembro de 2023. Cópia arquivada em 25 de agosto de 2023
- ↑ a b Pomel, Auguste (1851). «Nouvelles observations sur la structure des pieds dans les animaux de la famille des Anoplotherium, et dans le genre Hyaemoschus.». Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences. 33: 16–17. Consultado em 30 de agosto de 2023. Cópia arquivada em 3 de agosto de 2023
- ↑ Berthet, Didier (2003). «Le genre Cainotherium (Mammalia, Artiodactyla): étude morphométrique, révision systématique, implications évolutives et paléogéographiques, extinction». Travaux et Documents des Laboratoires de Géologie de Lyon. 159 (159): 3–205. Consultado em 30 de agosto de 2023. Cópia arquivada em 6 de agosto de 2023
- ↑ Lartet, Édouard (1837). «Note sur les ossements fossiles des terrains tertiaires de Simorre, de Sansan, etc., dans le département du Gers, et sur la découverte récente d'une mâchoire de singe fossile.». Comptes Rendus de l'Académie des Sciences. 4: 85–93. Consultado em 30 de agosto de 2023. Cópia arquivada em 13 de junho de 2020
- ↑ Lartet, Édouard (1839). «Notice géologique.». Extrait de l'Annuaire du Département du Gers
- ↑ Anquetin, Jérémy; Antoine, Pierre-Olivier; Tassy, Pascal (2007). «Middle Miocene Chalicotheriinae (Mammalia, Perissodactyla) from France, with a discussion on chalicotheriine phylogeny». Zoological Journal of the Linnean Society. 151 (3): 577–608. doi:10.1111/j.1096-3642.2007.00327.x
- ↑ Cautley, Proby T.; Falconer, Hugh (1837). «Notice on the Remains of a Fossil Monkey from the Tertiary Strata of the Sewalik Hills in the North of Hindoostan.». Transactions of the Geological Society: 499–504. Consultado em 30 de agosto de 2023. Cópia arquivada em 2 de agosto de 2023
- ↑ Lydekker, Richard (1886). Catalogue of the fossil Mammalia in the British museum, (Natural History): Part III. Containing the Order Ungulata, Suborders Perissodactyla, Toxondontia, Condylarthra, and Amblypoda. [S.l.]: Order of the Trustees, London. Consultado em 30 de agosto de 2023. Cópia arquivada em 2 de agosto de 2023
- ↑ Colbert, Edwin H. (1935). «The Proper Use of the Generic Name Nestoritherium». Journal of Mammalogy. 16 (3): 233–234. doi:10.1093/jmammal/16.3.233
- ↑ Buffetaut, Eric (2016). «From Charles Darwin's comments to the first mention of South American giant fossil birds: Auguste Bravard's catalogue of fossil species from Argentina (1860) and its significance». Bulletin de la Société Géologique de France. 187 (1): 41–53. doi:10.2113/gssgfbull.187.1.41
- ↑ Filhol, Henri (1877). «Recherches sur les Phosphorites du Quercy. Etude des fossiles qu'on y rencontre et spécialement des mammiféres.». Annales des Sciences Géologiques de Paris. Consultado em 30 de agosto de 2023. Cópia arquivada em 4 de agosto de 2023
- ↑ Rütimeyer, Ludwig (1862). «Eocaene Säugethiere aus dem Gebiet des schweizerischen Jura.». Neue Denkschriften der Schweizerischen Naturforschenden Gesellschaft. 19: 1–98. Consultado em 30 de agosto de 2023. Cópia arquivada em 4 de agosto de 2023
- ↑ von Fraas, Oscar Friedrich (1870). «Diplobune bavaricum.». Palaeontographica. 17: 177–184. Consultado em 30 de agosto de 2023. Cópia arquivada em 4 de agosto de 2023
- ↑ Stehlin, Hans Georg (1910). «Die Säugertiere des schweizerischen Eocaens. Sechster Teil: Catodontherium – Dacrytherium – Leptotherium – Anoplotherium – Diplobune – Xiphodon – Pseudamphimeryx – Amphimeryx – Dichodon – Haplomeryx – Tapirulus – Gelocus. Nachträge, Artiodactyla incertae sedis, Schlussbetrachtungen über die Artiodactylen, Nachträge zu den Perissodactylen.». Abhandlungen der Schweizerischen Paläontologischen Gesellschaft. 36. Consultado em 30 de agosto de 2023. Cópia arquivada em 5 de agosto de 2023
- ↑ Dietrich, Wilhelm Otto (1922). «Beitrag zur Kenntnis der säugetierführenden Bohnerzformation in Schwaben. 1. Ein vergessenes, neu erschlossenes Höhlenvorkommen terrestrischen Eozäns auf der Ulmer Alb.». Zentralblatt für Mineralogie, Geologie und Paläontologie. 19: 209–224
- ↑ a b c d e f g h i j k l m n Badiola, Ainara; De Vicuña, Nahia Jiménez; Perales-Gogenola, Leire; Gómez-Olivencia, Asier (2023). «First clear evidence of Anoplotherium (Mammalia, Artiodactyla) in the Iberian Peninsula: an update on the Iberian anoplotheriines». The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. PMC 12062563. PMID 37221992. doi:10.1002/ar.25238
- ↑ de Bonis, Louis (1964). «Étude de quelques mammifères du Ludien de La Débruge (Vaucluse)». Annales de Paléontologie: 121–154
- ↑ a b c Licht, Alexis; Métais, Grégoire; Coster, Pauline; İbilioğlu, Deniz; Ocakoğlu, Faruk; Westerweel, Jan; Mueller, Megan; Campbell, Clay; Mattingly, Spencer; Wood, Melissa C.; Beard, K. Christopher (2022). «Balkanatolia: The insular mammalian biogeographic province that partly paved the way to the Grande Coupure». Earth-Science Reviews. 226: 103929. Bibcode:2022ESRv..22603929L. doi:10.1016/j.earscirev.2022.103929
- ↑ a b c d Erfurt, Jörg; Métais, Grégoire (2007). «Endemic European Paleogene Artiodactyls». In: Prothero, Donald R.; Foss, Scott E. The Evolution of Artiodactyls. [S.l.]: Johns Hopkins University Press. pp. 59–84
- ↑ Orliac, Maeva; Gilissen, Emmanuel (2012). «Virtual endocranial cast of earliest Eocene Diacodexis (Artiodactyla, Mammalia) and morphological diversity of early artiodactyl brains». Proceedings of the Royal Society B. 279 (1743): 3670–3677. PMC 3415922. PMID 22764165. doi:10.1098/rspb.2012.1156
- ↑ a b c d Cuesta, Miguel-Ángel; Badiola, Ainara (2009). «Duerotherium sudrei gen. et sp. nov., a New Anoplotheriine Artiodactyl from the Middle Eocene of the Iberian Peninsula». Journal of Vertebrate Paleontology. 29 (1): 303–308. Bibcode:2009JVPal..29..303C. JSTOR 20491092. doi:10.1671/039.029.0110. Consultado em 30 de agosto de 2023. Cópia arquivada em 10 de agosto de 2023
- ↑ Schmidt-Kittler, Norbert; Godinot, Marc; Franzen, Jens L.; Hooker, Jeremy J. (1987). «European reference levels and correlation tables». Münchner geowissenschaftliche Abhandlungen A10. [S.l.]: Pfeil Verlag, München. pp. 13–31
- ↑ a b c Sudre, Jean; Martinez, Jean-Noël (1995). «The astragalus of Paleogene artiodactyls: comparative morphology, variability and prediction of body mass». Lethaia. 28 (3): 197–209. Bibcode:1995Letha..28..197M. doi:10.1111/j.1502-3931.1995.tb01423.x
- ↑ a b c d e f Weppe, Romain (2022). Déclin des artiodactyles endémiques européens, autopsie d'une extinction (Tese) (em francês). University of Montpellier. Consultado em 30 de agosto de 2023. Cópia arquivada em 11 de agosto de 2023
- ↑ a b c Luccisano, Vincent; Sudre, Jean; Lihoreau, Fabrice (2020). «Revision of the Eocene artiodactyls (Mammalia, Placentalia) from Aumelas and Saint-Martin-de-Londres (Montpellier limestones, Hérault, France) questions the early European artiodactyl radiation». Journal of Systematic Palaeontology. 18 (19): 1631–1656. Bibcode:2020JSPal..18.1631L. doi:10.1080/14772019.2020.1799253
- ↑ a b Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Pélissié, Thierry; Antoine, Pierre-Olivier; Orliac, Maëva Judith (2020). «Cainotheriidae (Mammalia, Artiodactyla) from Dams (Quercy, SW France): phylogenetic relationships and evolution around the Eocene–Oligocene transition (MP19–MP21)» (PDF). Journal of Systematic Palaeontology. 18 (7): 541–572. Bibcode:2020JSPal..18..541W. doi:10.1080/14772019.2019.1645754. Consultado em 19 de setembro de 2023. Cópia arquivada (PDF) em 7 de março de 2022
- ↑ Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Pélissié, Thierry; Orliac, Maëva Judith (2020). «A new Cainotherioidea (Mammalia, Artiodactyla) from Palembert (Quercy, SW France): Phylogenetic relationships and evolutionary history of the dental pattern of Cainotheriidae». Palaeontologia Electronica (23(3):a54). doi:10.26879/1081
- ↑ Gentry, Alan W.; Hooker, Jerry J. (1988). «The phylogeny of the Artiodactyla». The Phylogeny and Classification of the Tetrapods: Volume 2: Mammals (The Systematics Association Special Volume, No. 35B). [S.l.]: Oxford University Press. pp. 235–272
- ↑ a b c von Zittel, Karl Alfred (1925). Schlosser, Max, ed. Text-Book of Paleontology. Volume III. Mammalia. [S.l.]: Macmillan and Co. Limited. pp. 179–180. Consultado em 30 de agosto de 2023. Cópia arquivada em 14 de agosto de 2023
- ↑ Beddard, Frank Evers (1902). Harmer, Sidney Frederic; Shipley, Arthur Everett, eds. The Cambridge Natural History: Mammalia. [S.l.]: Macmillan and Co. Limited. pp. 332–333. Consultado em 30 de agosto de 2023. Cópia arquivada em 17 de agosto de 2023
- ↑ Scott, William B. (1945). «The Mammalia of the Duchesne River Oligocene». Transactions of the American Philosophical Society. 34 (3): 209–253. JSTOR 1005542. doi:10.2307/1005542
- ↑ a b c d e f g Palmer, R.W. (1913). «The Brain and Brain-Case of a Fossil Ungulate of the Genus Anoplotherium». Proceedings of the Zoological Society of London. 83 (4): 878–893. doi:10.1111/j.1096-3642.1913.tb01994.x. Consultado em 19 de setembro de 2023. Cópia arquivada em 13 de outubro de 2023
- ↑ Pearson, Helga Sharpe (1927). «On the Skulls of Early Tertiary Suidae, together with an Account of the Otic Region in Some Other Primitive Artiodactyla». Philosophical Transactions of the Royal Society of London. Series B, Containing Papers of a Biological Character. 215 (421–430): 440–445. doi:10.1098/rstb.1927.0009
- ↑ Gagnaison, Cyril; Leroux, Jean-Jacques (2013). «Un crâne de Diplobune secundaria Cuvier, 1822 de Saint-Capraise-d'Eymet (Dordogne)». Symbioses (em francês). 29: 43–46
- ↑ a b Jerison, Harry J. (2009). «Chapter 10: How Can Fossils Tell us About the Evolution of the Neocortex?». In: Kass, Jon H.; Striedter, Georg F.; Rubenstein, John L.R.; Bullock, Theodore H.; Krubitzer, Leah; Preuss, Todd. Evolutionary Neuroscience. [S.l.]: Academic Press. pp. 497–508
- ↑ Edinger, Tilly (1948). «Evolution of the Horse Brain». 25 : Evolution of the Horse Brain. Col: Geological Society of America Memoirs. 25. [S.l.]: Geological Society of America. pp. 1–178. doi:10.1130/MEM25-p1. Consultado em 19 de setembro de 2023. Cópia arquivada em 13 de outubro de 2023
- ↑ Thiery, Ghislain; Ducrocq, Stéphane (2015). «Endocasts and brain evolution in Anthracotheriidae (Artiodactyla, Hippopotamoidea)». Journal of Anatomy. 227 (3): 277–285. PMC 4560562. PMID 26278931. doi:10.1111/joa.12348
- ↑ Lihoreau, Fabrice; Boisserie, Jean-Renaud; Viriot, Laurent; Brunet, Michel (2006). «Anthracothere dental anatomy reveals a late Miocene Chado-Libyan bioprovince». Proceedings of the National Academy of Sciences. 103 (23): 8763–8767. Bibcode:2006PNAS..103.8763L. PMC 1482652. PMID 16723392. doi:10.1073/pnas.0603126103
- ↑ a b c Métais, Grégoire (2014). On the "thumb" of anoplotheriins: a 3D comparative study of the hand of Anoplotherium and Diplobune. Encontro Suíço de Geociências 2014
- ↑ a b Abusch-Siewert, Susanne (1989). «Bemerkungen zu den Anoplotherien (Artiodactyla, Mammalia) der Pariser Gipse». Münchner Geowissenschaftliche Abhandlungen A. 15: 55–78
- ↑ Heissig, Kurt (1993). «The Astragalus in Anoplotheres and Oreodonts, Phylogenetical and Paleogeographical Implications». Kaupia. 3: 173–178
- ↑ Prothero, Donald R.; Foss, Scott E., eds. (2007). «Summary». The Evolution of Artiodactyls. [S.l.]: Johns Hopkins University Press. 307 páginas
- ↑ de Carvalho, Carlos Neto; Muñiz, Fernando; Cáceres, Luis M.; Belaústegui, Zain; Rodríguez-Vidal, Joaquín; Belo, João; Moreira, Noel; Cachão, Mário; Cunha, Pedro P.; Figueiredo, Silvério; et al. (2022). «Aurochs roamed along the SW coast of Andalusia (Spain) during Late Pleistocene». Scientific Reports. 12 (9911): 9911. Bibcode:2022NatSR..12.9911D. PMC 9198092. PMID 35701579. doi:10.1038/s41598-022-14137-6
- ↑ a b c d Ellenberger, Paul (1980). «Sur les empreintes de pas des gros mammifères de l'Eocène supérieur de Garrigues-ste-Eulalie (Gard)». Palaeovertebrata. 9: 37–78. Consultado em 30 de agosto de 2023. Cópia arquivada em 29 de agosto de 2023
- ↑ Lucas, Spencer G.; Hunt, Adrian P. (2007). «Ichnotaxonomy of Camel Footprints». In: Lucas, Spencer G.; Spielmann, Justin A.; Lockley, Martin G. Cenozoic Vertebrate Tracks and Traces: Bulletin 42. [S.l.]: Museu de História Natural e Ciência do Novo México
- ↑ Montes, Martín Linares (2020). Paleontología e interpretación medioambiental de las icnitas de mamíferos del yacimiento de Fondota (Paleógeno, Abiego, Huesca) (PDF) (Tese). University of Zaragoza. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 19 de outubro de 2023
- ↑ Tsubamoto, Takehisa (2014). «Estimating body mass from the astragalus in mammals». Acta Palaeontologica Polonica. 59 (2): 259–265. doi:10.4202/app.2011.0067
- ↑ MacLaren, Jamie Alexander; Nauwelaerts, Sandra (2020). «Modern Tapirs as Morphofunctional Analogues for Locomotion in Endemic Eocene European Perissodactyls» (PDF). Journal of Mammalian Evolution. 27 (2): 245–263. doi:10.1007/s10914-019-09460-1. hdl:10067/1580640151162165141
- ↑ Joomun, Sarah C.; Hooker, Jerry J.; Collinson, Margaret E. (2009). Differences in the Dietary Responses of the Perissodactyl Plagiolophus and the Artiodactyl Diplobune to the Eocene/Oligocene Transition Events in Europe (PDF). 69th Annual Meeting Society of Vertebrate Paleontology and the 57th Symposium of Vertebrate Palaeontology and Comparative Anatomy (SVPCA). 29. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 31 de maio de 2023
- ↑ Joomun, Sarah C.; Hooker, Jerry J.; Collinson, Margaret E. (2010). «Changes in Dental Wear of Plagiolophus Minor (Mammalia: Perissodactyla) Across the Eocene-Oligocene Transition». Journal of Vertebrate Paleontology. 30 (2): 563–576. Bibcode:2010JVPal..30..563J. JSTOR 40666176. doi:10.1080/02724631003618124. Consultado em 30 de agosto de 2023. Cópia arquivada em 29 de agosto de 2023
- ↑ Cartmill, Matt; Brown, Kaye (2017). «Posture, Locomotion and Bipedality: The Case of the Gerenuk (Litocranius walleri)». In: Marom, Assaf; Hovers, Erella. Human Paleontology and Prehistory. Col: Vertebrate Paleobiology and Paleoanthropology. [S.l.]: Springer, Cham. pp. 53–70. ISBN 978-3-319-46644-6. doi:10.1007/978-3-319-46646-0_6
- ↑ Clark, Ciaran; Janis, Christine M.; Hooker, Jerry J.; Rayfield, Emily J. (2018). Posture in the fossil record: can bone microstructure be used to more accurately reconstruct past behaviour? (PDF). Associação Paleontológica: 62ª Reunião Anual; 14 a 17 de Dezembro de 2018; Universidade de Bristol. 72 páginas. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 18 de junho de 2022
- ↑ Linares Montes, Martín; Canudo, José Ignacio; Luzón, María Aránzazu; Castanera, Diego (2021). «Un excepcional registro paleoicnológico de artiodáctilos en el Oligoceno inferior de Abiego (Huesca, España)» (PDF). Comunicações Geológicas (em espanhol). 108: 103–107. doi:10.34637/6f8b-cj24. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 29 de agosto de 2023
- ↑ Eronen, Jussi T.; Janis, Christine M.; Chamberlain, Charles Page; Mulch, Andreas (2015). «Mountain uplift explains differences in Palaeogene patterns of mammalian evolution and extinction between North America and Europe». Proceedings of the Royal Society B. 282 (1809). PMC 4590438. PMID 26041349. doi:10.1098/rspb.2015.0136
- ↑ a b c Maitre, Elodie (2014). «Western European middle Eocene to early Oligocene Chiroptera: systematics, phylogeny and palaeoecology based on new material from the Quercy (France)». Swiss Journal of Palaeontology. 133 (2): 141–242. Bibcode:2014SwJP..133..141M. doi:10.1007/s13358-014-0069-3
- ↑ a b Badiola, Ainara; Perales-Gogenola, Leire; Astibia, Humberto; Suberbiola, Xabier Pereda (2022). «A synthesis of Eocene equoids (Perissodactyla, Mammalia) from the Iberian Peninsula: new signs of endemism». Historical Biology. 34 (8): 1623–1631. Bibcode:2022HBio...34.1623B. doi:10.1080/08912963.2022.2060098
- ↑ Robinet, Céline; Remy, Jean Albert; Laurent, Yves; Danilo, Laure; Lihoreau, Fabrice (2015). «A new genus of Lophiodontidae (Perissodactyla, Mammalia) from the early Eocene of La Borie (Southern France) and the origin of the genus Lophiodon Cuvier, 1822». Geobios. 48 (1): 25–38. Bibcode:2015Geobi..48...25R. doi:10.1016/j.geobios.2014.11.003
- ↑ Perales-Gogenola, Leire; Badiola, Ainara; Gómez-Olivencia, Asier; Pereda-Suberbiola, Xabier (2022). «A remarkable new paleotheriid (Mammalia) in the endemic Iberian Eocene perissodactyl fauna». Journal of Vertebrate Paleontology. 42 (4). Bibcode:2022JVPal..42E9447P. doi:10.1080/02724634.2023.2189447
- ↑ a b Solé, Floréal; Fischer, Valentin; Le Verger, Kévin; Mennecart, Bastien; Speijer, Robert P.; Peigné, Stéphane; Smith, Thierry (2022). «Evolution of European carnivorous mammal assemblages through the Paleogene». Biological Journal of the Linnean Society. 135 (4): 734–753. doi:10.1093/biolinnean/blac002
- ↑ a b Blondel, Cécile (2001). «The Eocene-Oligocene ungulates from Western Europe and their environment» (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 168 (1–2): 125–139. Bibcode:2001PPP...168..125B. doi:10.1016/S0031-0182(00)00252-2. Consultado em 30 de agosto de 2023. Cópia arquivada (PDF) em 22 de agosto de 2017
- ↑ Martin, Jeremy E.; Pochat-Cottilloux, Yohan; Laurent, Yves; Perrier, Vincent; Robert, Emmanuel; Antoine, Pierre-Olivier (2022). «Anatomy and phylogeny of an exceptionally large sebecid (Crocodylomorpha) from the middle Eocene of southern France». Journal of Vertebrate Paleontology. 42 (4). Bibcode:2022JVPal..42E3828M. doi:10.1080/02724634.2023.2193828
- ↑ Martin, Jeremy E. (2015). «A sebecosuchian in a middle Eocene karst with comments on the dorsal shield in Crocodylomorpha». Acta Palaeontologica Polonica. 60 (3): 673–680. doi:10.4202/app.00072.2014
- ↑ Antunes, Miguel Telles (2003). «Lower Paleogene Crocodilians from Silveirinha, Portugal». Palaeovenebrata. 32: 1–26. Consultado em 30 de agosto de 2023. Cópia arquivada em 29 de agosto de 2023
- ↑ Bai, Bin; Wang, Yuan-Qing; Theodor, Jessica M.; Meng, Jin (2023). «Small artiodactyls with tapir-like teeth from the middle Eocene of the Erlian Basin, Inner Mongolia, China». Frontiers in Earth Science. 11: 1–20. Bibcode:2023FrEaS..1117911B. doi:10.3389/feart.2023.1117911
- ↑ Kostopoulos, Dimitris S.; Koufos, George D.; Christanis, Kimon (2012). «On some anthracotheriid (Artiodactyla, Mammalia) remains from northern Greece: comments on the palaeozoogeography and phylogeny of Elomeryx». Swiss Journal of Palaeontology. 131 (2): 303–315. Bibcode:2012SwJP..131..303K. doi:10.1007/s13358-012-0041-z
- ↑ Hooker, Jerry J. (2013). «Origin and evolution of the Pseudorhyncocyonidae, a European Paleogene family of insectivorous placental mammals». Palaeontology. 56 (4): 807–835. Bibcode:2013Palgy..56..807H. doi:10.1111/pala.12018
- ↑ Marigó, Judit; Susanna, Ivette; Minwer-Barakat, Raef; Malapeira, Joan Madurell; Moyà-Solà, Salvador; Casanovas-Vilar, Isaac; Gimenez, Jose Maria Robles; Alba, David M. (2014). «The primate fossil record in the Iberian Peninsula». Journal of Iberian Geology. 40 (1): 179–211. doi:10.5209/rev_JIGE.2014.v40.n1.44094
- ↑ Manz, Carly; Bloch, Jonathan Ivan (2014). «Systematics and Phylogeny of Paleocene-Eocene Nyctitheriidae (Mammalia, Eulipotyphla?) with Description of a new Species from the Late Paleocene of the Clarks Fork Basin, Wyoming, USA». Journal of Mammalian Evolution. 22 (3): 307–342. doi:10.1007/s10914-014-9284-3
- ↑ Badiola, Ainara; Cuesta, Miguel-Ángel (2006). «Los marsupiales del yacimiento del Eoceno Superior de Zambrana (Álava, Región Vasco-Cantábrica)». Estudios Geológicos (em espanhol). 62 (1): 349–358. doi:10.3989/egeol.0662130
- ↑ Sigé, Bernard (1997). «Les mammiféres insectivoresdes nouvelles collections de Sossís et sites associes (Éocène supérieur, Espagne)». Geobios. 30 (1): 91–113. Bibcode:1997Geobi..30...91S. doi:10.1016/S0016-6995(97)80260-4
- ↑ Dawson, Mary R. (2003). «Paleogene rodents of Eurasia». Distribution and migration of tertiary mammals in Eurasia. 10. [S.l.: s.n.] pp. 97–127
- ↑ Chroust, Milan; Mazuch, Martin; Luján, Àngel Hernández (2019). «New crocodilian material from the Eocene-Oligocene transition of the NW Bohemia (Czech Republic): an updated fossil record in Central Europe during the Grande Coupure». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 293 (1): 73–82. doi:10.1127/njgpa/2019/0832
- ↑ Rage, Jean-Claude (2012). «Amphibians and squamates in the Eocene of Europe: what do they tell us?». Palaeobiodiversity and Palaeoenvironments. 92 (4): 445–457. Bibcode:2012PdPe...92..445R. doi:10.1007/s12549-012-0087-3
- ↑ Minwer-Barakat, Raef; Badiola, Ainara; Marigó, Judit; Moyà-Solà, Salvador (2013). «First record of the genus Microchoerus (Omomyidae, Primates) in the western Iberian Peninsula and its palaeobiogeographic implications». Journal of Human Evolution. 65 (3): 313–321. Bibcode:2013JHumE..65..313M. PMID 23916791. doi:10.1016/j.jhevol.2013.07.002
- ↑ a b Aguilar, Jean-Pierre; Legendre, Serge; Michaux, Jacques (1997). «Synthèses et tableaux de corrélations». Actes du Congrès Bio-chroM'97. Mémoires et Travaux de l'EPHE Institut de Montpellier 21 (em francês). [S.l.]: École Pratique des Hautes Études-Sciences de la Vie et de la Terre, Montpellier. pp. 769–850
- ↑ Sun, Jimin; Ni, Xijun; Bi, Shundong; Wu, Wenyu; Ye, Jie; Meng, Jin; Windley, Brian F. (2014). «Synchronous turnover of flora, fauna, and climate at the Eocene-Oligocene Boundary in Asia». Scientific Reports. 4 (7463): 7463. Bibcode:2014NatSR...4.7463S. PMC 4264005. PMID 25501388. doi:10.1038/srep07463
- ↑ a b c d Hooker, Jerry J.; Collinson, Margaret E.; Sille, Nicholas P. (2004). «Eocene–Oligocene mammalian faunal turnover in the Hampshire Basin, UK: calibration to the global time scale and the major cooling event» (PDF). Journal of the Geological Society. 161 (2): 161–172. Bibcode:2004JGSoc.161..161H. doi:10.1144/0016-764903-091. Consultado em 31 de agosto de 2023. Cópia arquivada (PDF) em 8 de agosto de 2023
- ↑ Legendre, Serge; Mourer-Chauviré, Cécile; Hugueney, Marguerite; Maitre, Elodie; Sigé, Bernard; Escarguel, Gilles (2006). «Dynamique de la diversité des mammifères et des oiseaux paléogènes du Massif Central (Quercy et Limagnes, France)». STRATA. 1 (em francês). 13: 275–282
- ↑ Escarguel, Gilles; Legendre, Serge; Sigé, Bernard (2008). «Unearthing deep-time biodiversity changes: The Palaeogene mammalian metacommunity of the Quercy and Limagne area (Massif Central, France)». Comptes Rendus Geoscience. 340 (9–10): 602–614. Bibcode:2008CRGeo.340..602E. doi:10.1016/j.crte.2007.11.005. Consultado em 19 de setembro de 2023. Cópia arquivada em 13 de outubro de 2023
- ↑ a b Costa, Elisenda; Garcés, Miguel; Sáez, Alberto; Cabrera, Lluís; López-Blanco, Miguel (2011). «The age of the "Grande Coupure" mammal turnover: New constraints from the Eocene–Oligocene record of the Eastern Ebro Basin (NE Spain)». Palaeogeography, Palaeoclimatology, Palaeoecology. 301 (1–4): 97–107. Bibcode:2011PPP...301...97C. doi:10.1016/j.palaeo.2011.01.005. hdl:2445/34510
- ↑ Hutchinson, David K.; Coxall, Helen K.; Lunt, Daniel J.; Steinthorsdottir, Margret; De Boer, Agatha M.; Baatsen, Michiel L.J.; Von der Heydt, Anna S.; Huber, Matthew; Kennedy-Asser, Alan T.; Kunzmann, Lutz; Ladant, Jean-Baptiste; Lear, Caroline; Moraweck, Karolin; Pearson, Paul; Piga, Emanuela; Pound, Matthew J.; Salzmann, Ulrich; Scher, Howie D.; Sijp, Willem P.; Śliwińska, Kasia K; Wilson, Paul A.; Zhang, Zhongshi (2021). «The Eocene-Oligocene transition: A review of marine and terrestrial proxy data, models and model-data comparisons». Climate of the Past. 17 (1): 269–315. Bibcode:2021CliPa..17..269H. doi:10.5194/cp-17-269-2021. hdl:11250/3135351
- ↑ Toumoulin, Agathe; Tardif, Delphine; Donnadieu, Yannick; Licht, Alexis; Ladant, Jean-Baptiste; Kunzmann, Lutz; Dupont-Nivet, Guillaume (2022). «Evolution of continental temperature seasonality from the Eocene greenhouse to the Oligocene icehouse –a model–data comparison». Climate of the Past. 18 (2): 341–362. Bibcode:2022CliPa..18..341T. doi:10.5194/cp-18-341-2022
- ↑ Boulila, Slah; Dupont-Nivet, Guillaume; Galbrun, Bruno; Bauer, Hugues; Châteauneuf, Jean-Jacques (2021). «Age and driving mechanisms of the Eocene–Oligocene transition from astronomical tuning of a lacustrine record (Rennes Basin, France)». Climate of the Past. 17 (6): 2343–2360. Bibcode:2021CliPa..17.2343B. doi:10.5194/cp-17-2343-2021
- ↑ Rivals, Florent; Belyaev, Ruslan I.; Basova, Vera B.; Prilepskaya, Natalya E. (2023). «Hogs, hippos or bears? Paleodiet of European Oligocene anthracotheres and entelodonts». Palaeogeography, Palaeoclimatology, Palaeoecology. 611: 111363. Bibcode:2023PPP...61111363R. doi:10.1016/j.palaeo.2022.111363
- ↑ Becker, Damien (2009). «Earliest record of rhinocerotoids (Mammalia: Perissodactyla) from Switzerland: systematics and biostratigraphy». Swiss Journal of Geosciences. 102 (3): 489–504. Bibcode:2009SwJG..102..489B. doi:10.1007/s00015-009-1330-4
- ↑ Solé, Floréal; Fischer, Fischer; Denayer, Julien; Speijer, Robert P.; Fournier, Morgane; Le Verger, Kévin; Ladevèze, Sandrine; Folie, Annelise; Smith, Thierry (2020). «The upper Eocene-Oligocene carnivorous mammals from the Quercy Phosphorites (France) housed in Belgian collections». Geologica Belgica. 24 (1–2): 1–16. doi:10.20341/gb.2020.006
- ↑ Kocsis, László; Ozsvárt, Péter; Becker, Damien; Ziegler, Reinhard; Scherler, Laureline; Vlad, Codrea A. (2014). «Orogeny forced terrestrial climate variation during the late Eocene–early Oligocene in Europe» (PDF). Geology. 42 (8): 727–730. Bibcode:2014Geo....42..727K. doi:10.1130/G35673.1. Consultado em 19 de setembro de 2023. Cópia arquivada (PDF) em 21 de setembro de 2017
- ↑ Hooker, Jerry J. (2010). «The 'Grande Coupure' in the Hampshire Basin, UK: taxonomy and stratigraphy of the mammals on either side of this major Palaeogene faunal turnover». In: Whittaker, John E.; Hart, Malcolm B. Micropalaeontology, Sedimentary Environments and Stratigraphy: a Tribute to Dennis Curry (1912–2001). 4. [S.l.]: Geological Society of London. pp. 147–215. ISBN 9781862396227. doi:10.1144/TMS004.8
- ↑ Joomun, Sarah C.; Hooker, Jerry J.; Collinson, Margaret E. (2010). Climate Change Versus Competition as the Cause of Ungulate Extinction at the Grande Coupure (Early Oligocene, Europe) (PDF). Congresso Internacional de Paleontologia: Londres 2010. 221 páginas. Consultado em 31 de agosto de 2023. Cópia arquivada (PDF) em 11 de junho de 2020
Ligações externas
 Media relacionados com Anoplotherium no Wikimedia Commons
Media relacionados com Anoplotherium no Wikimedia Commons