Thescelosaurus
| Thescelosaurus | |
|---|---|

| |
| Esqueleto montado no Museu de História Natural de Burpee em Rockford, Estados Unidos | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Classe: | Reptilia |
| Clado: | Dinosauria |
| Clado: | †Ornithischia |
| Família: | †Thescelosauridae |
| Subfamília: | †Thescelosaurinae |
| Gênero: | †Thescelosaurus Gilmore, 1913 |
| Espécie-tipo | |
| †Thescelosaurus neglectus Gilmore, 1913
| |
| Espécies | |
| |
| Sinónimos | |
Thescelosaurus é um gênero de dinossauro ornitísquio que viveu durante o período Cretáceo Superior no oeste da América do Norte. Foi nomeado e descrito em 1913 pelo paleontólogo Charles W. Gilmore; a espécie-tipo é T. neglectus. Duas outras espécies, T. garbanii e T. assiniboiensis, foram nomeadas em 1976 e 2011, respectivamente. Espécies adicionais foram sugeridas, mas atualmente não são aceitas. Thescelosaurus é o membro epônimo de sua família, Thescelosauridae. Os tescelossaurídeos são considerados ornitópodes basais ("primitivos") ou são colocados fora deste grupo dentro do grupo mais amplo Neornithischia.
O Thescelosaurus adulto teria medido aproximadamente 3 a 4 m de comprimento e provavelmente pesava de 200 a 300 kg. Movia-se sobre duas pernas e seu corpo era contrabalançado por sua longa cauda, que compunha metade do comprimento do corpo e era enrijecida por tendões ossificados em forma de bastão. O animal tinha um focinho longo e baixo que terminava em um bico sem dentes. Possuía mais dentes do que gêneros relacionados, e os dentes eram de tipos diferentes.
O Thescelosaurus foi encontrado em uma ampla distribuição geográfica no oeste da América do Norte. Os primeiros espécimes foram descobertos na Formação Lance, em Wyoming, mas descobertas subsequentes foram feitas na Dakota do Norte, Dakota do Sul, Montana, Alberta e Saskatchewan, em formações geológicas como a Formação Frenchman, a Formação Hell Creek e a Formação Scollard. Era relativamente comum e pode ter sido o dinossauro mais comum na Formação Frenchman. Vivendo no final do Maastrichtiano, foi um dos últimos dinossauros não aviários antes da extinção de todo o grupo durante o evento de extinção do Cretáceo-Paleogeno, há cerca de 66 milhões de anos.
Descoberta
T. neglectus e seu espécime tipo


Os primeiros espécimes do que mais tarde seria chamado de Thescelosaurus foram descobertos durante a Guerra dos Ossos, uma rivalidade acirrada entre os paleontólogos Edward Drinker Cope e Othniel Charles Marsh. Em julho de 1891, o caçador de fósseis John Bell Hatcher, contratado por Marsh, e seu assistente William H. Utterback descobriram um esqueleto quase completo de um pequeno dinossauro herbívoro ao longo do Riacho Doegie, no Condado de Niobrara, Wyoming, em rochas da Formação Lance. O esqueleto foi encontrado deitado sobre o lado esquerdo e em grande parte em articulação natural, com apenas a cabeça e o pescoço perdidos devido à erosão. Foi levado ao Museu Nacional de História Natural (USNM) do Instituto Smithsoniano,[a] onde permaneceu em sua caixa original, sem rótulo. Em 1903, a Marinha dos EUA contratou o paleontólogo Charles W. Gilmore para trabalhar na extensa coleção reunida sob a direção de Marsh, falecido em 1899. Foi somente em 1913 que Gilmore abriu a caixa e, para sua surpresa, encontrou o esqueleto de uma nova espécie de dinossauro.[2][3][4] Em 1913, Gilmore publicou uma descrição preliminar nomeando o novo gênero e espécie Thescelosaurus neglectus. Além do espécime de Hatcher (USNM 7757), que se tornou o espécime-tipo da nova espécie, Gilmore atribuiu um segundo esqueleto, mais fragmentário, de Lance Creek, também no Condado de Niobrara, à espécie (parátipo, USNM 7758). O nome genérico deriva das palavras gregas θέσκελος (theskelos), 'maravilhoso', e σαυρος (sauros), 'réptil' ou 'lagarto'.[2][5] O nome específico, neglectus, é latim para 'negligenciado' ou 'esquecido', já que o espécime tipo ficou abandonado por muito tempo.[4]
Gilmore publicou uma descrição abrangente em 1915, após o espécime-tipo estar totalmente preparado.[2][3] Ele identificou mais seis espécimes, incluindo uma escápula com coracoide, uma vértebra cervical e um osso do pé, bem como três esqueletos parciais que haviam sido coletados por Barnum Brown e estavam armazenados no Museu Americano de História Natural (AMNH). O pescoço e o crânio permaneceram desconhecidos, no entanto, e Gilmore restaurou essas partes ausentes com base no Hypsilophodon, que ele considerava um parente próximo, em suas reconstruções esqueléticas e de vida. Para a exibição do espécime-tipo no museu, Gilmore manteve sua postura original e incompleta. Apenas a perna direita, que estava ligeiramente deslocada, foi ajustada em posição, e alguns pequenos danos aos ossos foram restaurados, mas pintados mais claros do que os ossos originais para que as partes reais e reconstruídas pudessem ser distinguidas visualmente.[3] Em 1963, a exibição foi incluída em uma montagem de parede ao lado dos ornitísquios Edmontosaurus e Corythosaurus e do terópode Gorgosaurus. Em 1981, a exposição foi reorganizada, colocando o Thescelosaurus em um local mais alto e fora de vista. As reformas da exposição entre 2014 e 2019 removeram o Thescelosaurus e outros dinossauros em exposição, substituindo-os por moldes de gesso para que os fósseis originais pudessem ser preparados e estudados posteriormente.[6]
T. edmontonensis, revisão e T. garbanii
Em 1926, William Parks descreveu a nova espécie T. warreni a partir de um esqueleto bem preservado de Alberta, Canadá, do que era então conhecido como Formação Edmonton.[7] Este esqueleto tinha diferenças notáveis de T. neglectus, e então Charles M. Sternberg o colocou em um novo gênero, Parksosaurus, em 1937.[8] Em 1940, Sternberg nomeou uma espécie adicional, T. edmontonensis, com base em outro esqueleto articulado (CMN 8537) que ele havia descoberto na Formação Edmonton de Rumsey, Alberta. Sternberg já havia mencionado este espécime em 1926, embora ainda não estivesse preparado naquela época.[9][10] Ele preserva a maior parte da coluna vertebral, pélvis, pernas, escápula, coracoide, braço e, mais significativamente, vários ossos do teto do crânio e uma mandíbula completa, a primeira conhecida de Thescelosaurus.[10] A geologia mais recente separou a Formação Edmonton em quatro formações, com Parksosaurus da mais antiga Formação Horseshoe Canyon e Thescelosaurus edmontonensis da mais jovem Formação Scollard.[11]


Em 1974, Peter M. Galton revisou o Thescelosaurus e descreveu espécimes adicionais, resultando em um total de 15 espécimes conhecidos. Estes incluem quatro espécimes da Formação Hell Creek coletados por Barnum Brown em Montana, EUA, em 1906 e 1909, alguns dos quais já haviam sido mencionados por Gilmore em 1915; um espécime encontrado em 1892 por Wortman e Peterson em um local incerto; dois espécimes encontrados em 1921 por Levi Sternberg na Formação Frenchman de Rocky Creek, Saskatchewan e dois ossos isolados, também de Saskatchewan. Um dos espécimes de Brown, AMNH 5034, foi encontrado apenas 1,5 m abaixo da Formação Fort Union, na localidade mais jovem onde dinossauros foram encontrados. Galton concluiu que T. edmontonensis era simplesmente um indivíduo mais robusto de T. neglectus (possivelmente o sexo oposto do indivíduo-tipo).[12]
William J. Morris descreveu três esqueletos parciais adicionais em 1976, dois encontrados na Formação Hell Creek, no Condado de Garfield, Montana, pelo preparador Harli Garbani, e um de um local desconhecido no Condado de Harding, Dakota do Sul. O primeiro espécime (LACM 33543) preserva partes da coluna vertebral e da pelve, além de ossos do crânio ainda não conhecidos do Thescelosaurus, como as jugais e a caixa craniana. O segundo espécime (LACM 33542) inclui vértebras do pescoço e das costas, e uma perna quase completa com um fêmur parcial. Morris concluiu que a anatomia do tornozelo e o tamanho maior eram únicos e, portanto, nomeou a nova espécie Thescelosaurus garbanii, em homenagem ao descobridor Garbani. Morris também argumentou que o tornozelo do T. edmontonensis, que Galton alegou estar danificado e mal interpretado, era realmente diferente do T. neglectus e mais semelhante ao T. garbanii. Portanto, ele sugeriu que T. edmontonensis e T. garbanii poderiam eventualmente ser separados de Thescelosaurus como um novo gênero. O terceiro espécime (SDSM 7210) inclui uma grande parte do crânio, algumas vértebras parciais das costas e dois ossos dos dedos, partes que não se sobrepõem às regiões diagnósticas do espécime tipo T. neglectus, impedindo comparações. Morris atribuiu provisoriamente o espécime a Thescelosaurus, mas sugeriu que ele poderia representar uma nova espécie; essa espécie potencial foi posteriormente chamada de "hipsilofodontídeo de Hell Creek".[13][14]
Bugenasaura e o espécime "Willo"

Galton revisou Thescelosaurus pela segunda vez em 1995. Ele argumentou que as características supostamente diagnósticas do tornozelo do espécime de T. edmontonensis são o resultado de quebra, como indicado pelo tornozelo esquerdo previamente não descrito daquele espécime que apresentava a mesma anatomia de T. neglectus. Consequentemente, ele sinonimizou T. edmontonensis com T. neglectus. Galton determinou que Morris interpretou corretamente o tornozelo de T. garbanii e sugeriu que a espécie poderia ser elevada a um gênero próprio. Havia também a possibilidade de que o membro posterior de T. garbanii pertencesse ao paquicefalossaurídeo Stygimoloch, que também é conhecido da Formação Hell Creek e para o qual o membro posterior era desconhecido. Galton também concluiu que o crânio de SDSM 7210, o terceiro dos espécimes descritos por Morris, era distinto de Thescelosaurus e, portanto, nomeou o novo táxon Bugenasaura infernalis. O nome é uma combinação do latim bu, "grande", e gena, "bochecha", bem como do grego antigo saura, "lagarto". O nome específico, do latim infernalis, "pertencente às regiões mais baixas", é uma referência aos níveis mais baixos da Formação Hell Creek, de onde é conhecido. Galton também atribuiu provisoriamente LACM 33543, o tipo de T. garbanii, à nova espécie, observando que material adicional é necessário para determinar se a referência está correta e que o nome garbanii deve ter prioridade se este for o caso.[15]

Em sua revisão de 1995, Galton também reatribuiu dentes isolados da Formação Judith River, do Campaniano de Montana, ao gênero relacionado Orodromeus. Esses dentes haviam sido atribuídos a Thescelosaurus cf. neglectus por Ashok Sahni em 1972, o que teria sido a ocorrência mais antiga de Thescelosaurus.[15][16]Em um estudo de 1999 sobre a anatomia de Bugenasaura, Galton atribuiu um dente da coleção do Museu de Paleontologia da Universidade da Califórnia (UCMP 49611) a este último. Significativamente, esse dente supostamente veio da Formação Kimmeridge Clay, do Jurássico Superior, em Weymouth, Inglaterra, e, portanto, é cerca de 70 milhões de anos mais antigo que o espécime tipo Bugenasaura e de outro continente. Galton argumentou que ele possivelmente havia sido rotulado incorretamente e era, na verdade, da Formação Lance, em Wyoming, mas o dente foi coletado pela primeira vez antes de o museu estar ativo na região de Lance.[17] A falta de características diagnósticas levou Paul M. Barrett e Susannah Maidment a classificar o dente como um ornitísquio indeterminado em 2011.[18]
T. assiniboiensis e outras descobertas
Outra espécie, T. assiniboiensis, foi nomeada por Caleb M. Brown e colegas em 2011 com base em um espécime (RSM P 1225.1) encontrado em 1968 por Albert Swanston, técnico de museu no Museu Real de Saskatchewan. O nome específico, assiniboiensis, deriva do histórico Distrito de Assiniboia, que abrangia a região sul de Saskatchewan, onde a Formação Frenchman está exposta, que por sua vez recebe o nome dos povos Assiniboine. Quando descoberto, o espécime era articulado, com a cauda saindo da encosta de uma colina. É um espécime pequeno, incluindo um crânio fragmentado, a maior parte da coluna vertebral, as cinturas pélvicas e os membros posteriores. A localização do espécime, conforme relatado originalmente, estava incorreta, pois uma revisita ao vale do Rio Frenchman por Tim Tokaryk na década de 1980 revelou que a escavação, identificável por restos de osso e gesso, na verdade ocorreu no lado norte do vale, aproximadamente na metade do argilito exposto. Isso situa o espécime na Formação Frenchman.[19]
Os espécimes só podem ser comparados diretamente se preservarem os mesmos ossos, mas material sobreposto frequentemente não está disponível – a atribuição da maioria dos espécimes de Thescelosaurus a qualquer uma das três espécies reconhecidas, portanto, permaneceu incerta. Essa situação melhorou em 2014, quando Boyd e colegas relataram um novo espécime da Formação Hell Creek, no Condado de Dewey, Dakota do Sul (TLAM.BA.2014.027.0001), que foi coletado em terras privadas por Bill Alley antes de ser doado ao Museu Timber Lake and Area. Este espécime ainda não havia sido totalmente preparado, mas inclui um crânio quase completo, porém ligeiramente esmagado, e grande parte do esqueleto. Esta descoberta permitiu a atribuição deste espécime e do espécime "Willo" a T. neglectus.[20] Em 2022, a mídia noticiou que um espécime de Thescelosaurus foi encontrado no sítio fóssil de Tanis em Dakota do Norte, que se acredita mostrar sinais diretos do impacto do asteroide Chicxulub no Golfo do México que resultou na extinção K-Pg.[21][22]
Descrição

A anatomia esquelética deste gênero é bem documentada em geral, e restaurações foram publicadas em vários artigos, incluindo restaurações esqueléticas[23][3][12][24] e modelos.[10][3] O esqueleto é conhecido bem o suficiente para que uma reconstrução detalhada dos músculos do quadril e dos membros posteriores tenha sido feita.[25] O maior thescelossaurídeo conhecido,[26][19] seu comprimento corporal foi estimado em 2,5–4,0 m[12] e o peso em 200–300 kg,[27] com o grande espécime tipo de T. garbanii estimado em 4–4,5 m de comprimento.[13] Pode ter sido sexualmente dimórfico, com um sexo maior que o outro.[12]
Crânio
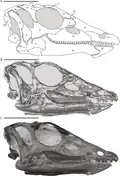


O crânio é mais conhecido de T. neglectus, principalmente graças ao espécime "Willo" excelentemente preservado, que foi submetido a tomografia computadorizada para revelar seus detalhes internos. Um crânio fragmentário também é conhecido de T. assiniboiensis (RSM P 1225.1). A maioria das autapomorfias – características distintivas que não são encontradas em gêneros relacionados – são encontradas no crânio. O crânio também apresenta muitas plesiomorfias, características "primitivas" (basais) que são tipicamente encontradas em ornitísquios geologicamente muito mais antigos, mas também apresenta características derivadas (avançadas).[28]
O crânio tinha um focinho longo e baixo que terminava em um bico sem dentes.[29][30] Como em outros dinossauros, era perfurado por várias fenestras, ou aberturas no crânio. Destas, a órbita ocular e a fenestra infratemporal (atrás da órbita) eram proporcionalmente grandes, enquanto a narina externa era pequena.[29] A narina externa era formada pela pré-maxila (o osso frontal da mandíbula superior) e o osso nasal, enquanto a maxila (o osso da "bochecha" com dentes) foi excluída.[31] Outra fenestra, a fenestra anterorbital, ficava entre a narina externa e a órbita e continha duas fenestras internas menores.[32] Ossos longos em forma de haste, chamados palpebrais, estavam presentes acima dos olhos, dando ao animal sobrancelhas pesadas e ósseas.[30] A pálpebra não estava alinhada com a margem superior da órbita como em alguns outros ornitísquios, mas projetava-se através dela.[33] Os ossos frontais, que formam o teto do crânio acima da órbita, eram mais largos no nível do meio da órbita e mais estreitos em suas extremidades posteriores (traseiras) - uma autapomorfia do Thescelosaurus.[34]
Vértebras e membros
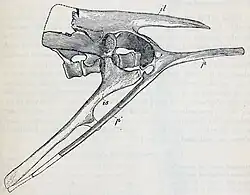
T. neglectus tinha seis vértebras sacrais ("quadril") e 27 vértebras pré-sacrais ("pescoço e costas").[13][19] O espécime tipo de T. assiniboiensis parece ter tido apenas cinco sacrais, mas é possível que este indivíduo ainda não estivesse totalmente maduro e que o último sacral ainda não estivesse fundido aos outros sacrais.[19] A cauda era longa e compunha metade do comprimento total do corpo. Era sustentada por tendões ossificados do meio à ponta, o que reduziria a flexibilidade da cauda.[3] A caixa torácica era larga, dando-lhe um dorso largo.[10] Grandes placas mineralizadas, finas e planas, foram encontradas próximas às laterais das costelas, as chamadas placas intercostais.[35] As costelas anteriores eram achatadas e côncavas, e as margens posteriores de suas extremidades inferiores tinham uma superfície áspera. Essas características são autapomorfias do Thescelosaurus e possivelmente adaptações que permitem a fixação das placas à caixa torácica.[36]
Os membros eram robustos.[10] O fêmur (osso da coxa) era mais longo que a tíbia (osso da canela), o que distingue o gênero de gêneros intimamente relacionados.[36] O Thescelosaurus tinha mãos curtas, largas e com cinco dedos. O segundo dígito era o mais longo, e o quinto dígito era bastante reduzido em tamanho. Apenas os três primeiros dígitos terminavam em ungueais semelhantes a cascos. Havia duas falanges (ossos dos dedos) no primeiro dígito, três no segundo, quatro no terceiro, três no quarto e dois no quinto.[3] O pé tinha cinco metatarsos, embora apenas os quatro primeiros possuíssem dígitos, sendo o quinto metatarso vestigial (reduzido a uma pequena tala). O primeiro metatarso tinha apenas metade do comprimento do terceiro, e seu dígito pode não ter tocado o solo regularmente. A maior parte do peso do animal era, portanto, suportada pelos três dígitos centrais, dos quais o do meio (terceiro) era o mais longo. O primeiro dígito tinha duas falanges, o segundo tinha três, o terceiro tinha quatro e o quarto tinha cinco.[19][3] Os dígitos eram mais curtos que os metatarsos, e suas falanges eram distintamente achatadas.[3][37] A espécie T. garbanii difere das outras espécies em seu tornozelo único, com o calcâneo sendo reduzido e não contribuindo para a articulação mediotarsal.[38][13]
Tegumento

Durante a maior parte de sua história, a natureza do tegumento deste gênero, sejam escamas ou algo mais, permaneceu desconhecida. Gilmore descreveu manchas de material carbonizado perto dos ombros como possível epiderme, com uma textura "perfurada", mas sem padrão regular,[3] enquanto Morris sugeriu que a armadura estava presente, na forma de pequenas placas que ele interpretou como localizadas pelo menos ao longo da linha média do pescoço de um espécime.[13] Placas não foram encontradas com outros espécimes articulados de Thescelosaurus, no entanto, e Galton argumentou em 2008 que as placas de Morris são de origem crocodiliana[39] Em 2022, a mídia noticiou que o espécime de Tanis preserva impressões de pele em uma perna que mostram que partes do animal estavam cobertas por escamas.[21]
Classificação
Em sua descrição de Thescelosaurus em 1913, Gilmore o considerou um membro de Camptosauridae, ao lado de Hypsilophodon, Dryosaurus e Laosaurus. Em 1915, ele o colocou dentro de Hypsilophodontidae, ao lado apenas de Hypsilophodon.[2][3] Muitos autores seguiram essa classificação dentro de Hypsilophodontidae.[40] Franz Nopcsa e Friedrich von Huene, por sua vez, mantiveram Thescelosaurus como parente de Camptosaurus em 1928 e 1956, respectivamente.[41][42] Em 1937, Sternberg separou Thescelosaurus e o relacionado Parksosaurus em uma família própria, Thescelosauridae, mas considerou ambos os gêneros como membros da subfamília Thescelosaurinae dentro de Hypsilophodontidae em 1940.[8][10] Anatoly Konstantinovich Rozhdestvensky e Richard A. Thulborn mantiveram Thescelosauridae como uma família separada em 1964 e 1974, respectivamente.[43][44] Galton classificou Thescelosaurus como um membro de Iguanodontidae com base nas proporções dos membros posteriores em 1974, mas esta família foi considerada polifilética (não um grupo natural);[12] ele, portanto, retornou à classificação de hipsilofodontídeos em 1995.[15]
Hypsilophodontidae incluiu apenas quatro gêneros em 1940: Hypsilophodon, Thescelosaurus, Parksosaurus e Dysalotosaurus.[10] Em 1966, Alfred Sherwood Romer atribuiu a maioria dos pequenos ornitópodes à família, seguido por Galton e autores posteriores, embora Thescelosaurus nem sempre tenha sido incluído na família. Como resultado, Hypsilophodontidae incluiu treze gêneros na primeira edição do livro The Dinosauria, em 1990.[45][12][46] Este conceito de Hypsilophodontidae como um grupo monofilético (natural) inclusivo foi apoiado pelos primeiros estudos cladísticos de Paul C. Sereno, David B. Weishampel e Ronald Heinrich, que descobriram que Thescelosaurus era o hipsilofodontídeo mais basal. A análise de Weishampel e Heinrich, em 1992, pode ser vista abaixo.[47][48]

| Hypsilophodontidae |
| ||||||||||||||||||||||||||||||||||||
O conceito de Hypsilophodontidae como um grupo monofilético caiu em desuso. Rodney Sheetz sugeriu em 1999 que "hipsilofodontídeos" eram simplesmente as formas primitivas de ornitópodes, o grupo maior ao qual eram comumente atribuídos. Scheetz descobriu que Thescelosaurus, Parksosaurus e Bugenasaura estavam sucessivamente mais próximos de Hypsilophodon e, posteriormente, de ornitópodes, mas não de um grupo próprio.[49] Outros estudos apresentaram resultados semelhantes, com Thescelosaurus ou Bugenasaura como ornitópodes primitivos próximos à origem do grupo, às vezes formando um clado com Parksosaurus.[50][51][52] Um problema com T. neglectus antes da revisão por Boyd e colegas em 2009 era a incerteza sobre os espécimes atribuídos, incluindo a separação de Bugenasaura e a questão não resolvida de se T. edmontonensis era distinto ou não.[52] Após a revisão taxonômica, as relações sistemáticas entre Thescelosaurus e os "hipsilofodontes" tornaram-se mais claras, e Boyd e colegas encontraram suporte para um grupo maior de ornitópodes primitivos, composto por Thescelosaurus, Parksosaurus, Zephyrosaurus, Orodromeus e Oryctodromeus.[23] Brown e colegas, ao descreverem T. assiniboiensis em 2011, chegaram a resultados semelhantes.[19] Os mesmos autores confirmaram esses resultados novamente em 2013, o que os levou a reintroduzir o nome Thescelosauridae para todo o grupo, que foi dividido na subfamília revisada Thescelosaurinae e na nova subfamília Orodrominae.[53][54]
Outros estudos não encontraram parentesco próximo entre Parksosaurus e Thescelosaurus, propondo, em vez disso, que ele fosse parente do Gasparinisaura sul-americano. No entanto, Boyd argumentou que a anatomia do Parksosaurus havia sido mal interpretada e que este e Thescelosaurus eram parentes muito próximos, se não parentes mais próximos um do outro.[20] Os clados Thescelosauridae (ou, alternativamente, Parksosauridae) e Thescelosaurinae foram confirmados por inúmeras análises filogenéticas,[53][40][55][56][57] embora não por todas.[58][59] Também há desacordo sobre se Thescelosaurus e tescelossaurídeos são membros de Ornithopoda ou mais basais. Boyd destacou em 2015 que muitos estudos filogenéticos que incluíram Thescelosaurus não incluem marginocefálos ou não são resolvidos, então não havia evidência definitiva de que Thescelosaurus era um ornitópode. Em sua análise, Thescelosaurus e Thescelosauridae estavam fora de Ornithopoda, formando um clado expansivo de neornithischianos não ornitópodes.[40] Alguns estudos concordam com essa colocação para tescelossaurídeos,[55][58] enquanto outros apoiam Thescelosaurus como um ornitópode,[59] e outros não estão resolvidos.[56][57] Fonseca e colegas deram o nome Pyrodontia ao clado que une Thescelosaurus com ornitísquios mais derivados quando Thescelosauridae está fora de Ornithopoda, referenciando a diversificação precoce e rápida de Thescelosauridae, Marginocephalia e Ornithopoda. Os resultados de Fonseca e colegas sobre thescelosaurids em 2024 podem ser vistos abaixo.[55]

| Thescelosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Os primeiros tescelossaurídeos conhecidos, Changchunsaurus e Zephyrosaurus, são do Cretáceo médio, cerca de 40 milhões de anos mais jovens do que quando o grupo teria evoluído, sugerindo uma longa linhagem fantasma (um período de tempo geológico durante o qual um grupo existiu, mas não deixou evidências fósseis).[40] Em 2024, André Fonseca e colegas recuperaram o Nanosaurus do Jurássico Superior como o primeiro tescelossaurídeo, o que encurtaria a linhagem fantasma.[55][58] Boyd concluiu em 2015 que a divisão entre Orodrominae e Thescelosaurinae ocorreu na América do Norte no estágio Aptiano, com Orodrominae se diversificando na América do Norte.[40] Thescelosaurinae pode ter se diversificado na América do Norte ou na Ásia;[40] o gênero Fona, descrito em 2024, sugere que Thescelosaurinae já estava estabelecido na América do Norte no início do Cretáceo Superior.[57]
Paleobiologia
Como outros ornitísquios, o Thescelosaurus provavelmente era herbívoro.[29] Os diferentes tipos de dentes, bem como o focinho estreito, sugerem que se alimentava seletivamente. O paquicefalossauro contemporâneo Stegoceras, em contraste, provavelmente se alimentava de forma mais indiscriminada, permitindo que ambos os animais compartilhassem o mesmo ambiente sem competir por alimento (particionamento de nicho).[60] Sabe-se que um espécime apresentava uma patologia óssea, com os ossos longos do pé direito fundidos na parte superior.[27]
Postura e locomoção


Em sua descrição de 1915, Gilmore sugeriu que o Thescelosaurus era um animal ágil, bípede (de duas pernas) e adaptado para correr. Ele também criou um modelo para representar sua aparência em vida, mostrando um corpo leve e ágil, construído com membros posteriores delgados.[3] Essas ideias foram contestadas por Sternberg em 1940, que argumentou que o esqueleto, e especialmente os membros, eram robustos. Seu próprio modelo, da espécie T. edmontonensis, consequentemente mostrou membros muito mais musculosos.[10] Outros estudos subsequentes discordaram da ideia de Gilmore de um corredor proficiente, dado o esqueleto robusto, o fêmur proporcionalmente longo e os ossos curtos da perna.[26] Galton, em 1974, chegou a sugerir que o Thescelosaurus poderia ter se movido ocasionalmente de quatro, dados seus braços bastante longos e mãos largas[12] Phil Senter e Jared Mackey, em 2024, concluíram que uma postura quadrúpede teria sido teoricamente possível, já que a coluna vertebral estava curvada para baixo, permitindo que a mão tocasse o solo mesmo com os membros posteriores retos. No entanto, em tal postura, os dedos teriam apontado para os lados em vez de para a frente e, consequentemente, não poderiam ter sido usados para impulsionar o animal para a frente; a locomoção quadrúpede, portanto, parece improvável.[61]
Um estudo de 2023 realizado por David Button e Lindsay Zanno concluiu que o Thescelosaurus era menos adaptado para correr do que outros tescelossaurídeos, mas, ainda assim, apresentava duas características comuns em corredores. A primeira delas é o quarto trocanter, uma crista óssea no fêmur que ancorava o principal músculo locomotor. Essa crista era relativamente proximal (mais próxima da extremidade superior do osso), permitindo movimentos mais rápidos em detrimento da potência. A segunda característica é encontrada no ouvido interno, que contém os três canais semicirculares que abrigam o senso de equilíbrio: um desses canais, o canal semicircular anterior, era bastante alargado, sugerindo sensibilidade aguda ao equilíbrio, o que, por sua vez, pode sugerir alta agilidade, mas também pode ser explicado por um possível comportamento de escavação.[26]
Ver também
Notas
- ↑ Na época conhecido como "Museu Nacional dos Estados Unidos"
Referências
- ↑ Boyd, Clint A.; Brown, Caleb M.; Scheetz, Rodney D.; Clarke, Julia A. (2009). «Taxonomic revision of the basal neornithischian taxa Thescelosaurus and Bugenasaura». Journal of Vertebrate Paleontology. 29 (3): 758–770. doi:10.1671/039.029.0328
- ↑ a b c d Gilmore, C.W. (1913). «A new dinosaur from the Lance Formation of Wyoming». Smithsonian Miscellaneous Collections. 61 (5): 1–5
- ↑ a b c d e f g h i j k l Gilmore, C.W. (1915). «Osteology of Thescelosaurus, an orthopodous dinosaur from the Lance Formation of Wyoming». Proceedings of the United States National Museum. 49 (2127): 591–616. doi:10.5479/si.00963801.49-2127.591
- ↑ a b Boyd, Clint A. (2016). «The "overlooked marvelous reptile"» (PDF). Geo News. 43 (1): 2–6. Consultado em 5 de dezembro de 2024. Cópia arquivada (PDF) em 21 de julho de 2024
- ↑ Lucas, S.G. (2016). Dinosaurs: The Textbook 6th ed. [S.l.]: Columbia University Press. p. XXIII. ISBN 978-0-231-54184-8
- ↑ Miller, B.H. (5 de maio de 2015). «Extinct Monsters: The Marsh Dinosaurs, Part III». Extinct Monsters. Consultado em 31 de outubro de 2024. Cópia arquivada em 1 de dezembro de 2023
- ↑ Parks, W.A. (1926). «Thescelosaurus warreni, a new species of orthopodous dinosaur from the Edmonton Formation of Alberta». University of Toronto Studies, Geological Series. 21: 1–42
- ↑ a b Sternberg, C.M. (1937). «Classification of Thescelosaurus, with a description of a new species». Geological Society of America, Proceedings of the Paleontological Society, 1936: 375
- ↑ Sternberg, C.M. (1926). «Notes on the Edmonton Formation of Alberta». The Canadian Field-Naturalist. 40 (5): 102–105. doi:10.5962/p.338660

- ↑ a b c d e f g h Sternberg, C.M. (1940). «Thescelosaurus edmontonensis, n. sp., and Classification of the Hypsilophodontidae». Journal of Paleontology. 14 (5): 481–494
- ↑ Eberth, D.A.; Kamo, S.L. (2019). «High-precision U-Pb CA-ID-TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River valley, Alberta, Canada». Canadian Journal of Earth Sciences. 57 (10): 1220–1237. doi:10.1139/cjes-2019-0019
- ↑ a b c d e f g Galton, P.M. (1974). «Notes on Thescelosaurus, a Conservative Ornithopod Dinosaur from the Upper Cretaceous of North America, with Comments on Ornithopod Classification». Journal of Paleontology. 48 (5): 1048–1067
- ↑ a b c d e Morris, W.J. (1976). «Hypsilophodont dinosaurs: a new species and comments on their systematics». In: Churcher, C.S. Athlon: Essays on Palaeontology in honour of Loris Shano Russell. [S.l.]: Royal Ontario Museum, Life Sciences Miscellaneous Publications. pp. 93–113. ISBN 978-0-88854-157-4
- ↑ Sues, H.-D. (1980). «Anatomy and relationships of a new hypsilophodontid dinosaur from the Lower Cretaceous of North America». Palaeontographica Abteilung A. 169 (1–3): 51–72
- ↑ a b c Galton, P.M. (1995). «The species of the basal hypsilophodontid dinosaur Thescelosaurus Gilmore (Ornithischia: Ornithopoda) from the Late Cretaceous of North America». Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 198 (3): 297–311. Bibcode:1995NJGPA.198..297G. doi:10.1127/njgpa/198/1995/297
- ↑ Sahni, A. (1972). «The vertebrate fauna of the Judith River Formation, Montana». Bulletin of the American Museum of Natural History. 147 (6): 321–412
- ↑ Galton, P.M. (1999). «Cranial anatomy of the hysilophodontid dinosaur Bugenasaura infernalis (Ornithischia: Ornithopoda) from the Upper Cretaceous of North America». Revue de Paléobiologie. 18 (2): 517–534
- ↑ Barrett, P.M.; Maidment, S.C.R. (2011). «Dinosaurs of Dorset: Part III, the ornithischian dinosaurs (Dinosauria, Ornithischia) with additional comments on the sauropods». Proceedings of the Dorset Natural History and Archaeological Society. 132: 145–163
- ↑ a b c d e f Brown, C. M.; Boyd, C.A.; Russell, A.P. (2011). «A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America». Zoological Journal of the Linnean Society. 163 (4): 1157–1198. doi:10.1111/j.1096-3642.2011.00735.x
- ↑ a b Boyd, C.A. (2014). «The cranial anatomy of the neornithischian dinosaur Thescelosaurus neglectus». PeerJ. 2: e669. PMC 4232843. PMID 25405076. doi:10.7717/peerj.669
- ↑ a b Amos, Jonathan (6 de abril de 2022). «Tanis: Fossil found of dinosaur killed in asteroid strike, scientists claim». BBC News (em inglês). Consultado em 25 de outubro de 2022. Cópia arquivada em 24 de outubro de 2022
- ↑ Martin, Saleen. «Fossil of dinosaur killed in asteroid strike discovered in North Dakota, scientists say». phys.org (em inglês). Consultado em 25 de outubro de 2022. Cópia arquivada em 25 de outubro de 2022
- ↑ a b Boyd, C.A.; Brown, C.M.; Scheetz, R.D.; Clarke, J.A. (2009). «Taxonomic revision of the basal neornithischian taxa Thescelosaurus and Bugenasaura». Journal of Vertebrate Paleontology. 29 (3): 758–770. Bibcode:2009JVPal..29..758B. doi:10.1671/039.029.0328
- ↑ Brett-Surman, Michael K. (1997). «Ornithopods». In: Farlow, James O.; Brett-Surman, Michael K. The Complete Dinosaur. Bloomington and Indianapolis: Indiana University Press. pp. 330–346. ISBN 978-0-253-33349-0
- ↑ Romer, Alfred S. (1927). «The pelvic musculature of ornithischian dinosaurs». Acta Zoologica. 8 (2–3): 225–275. doi:10.1111/j.1463-6395.1927.tb00653.x
- ↑ a b Erickson, Bruce R. (2003). Dinosaurs of the Science Museum of Minnesota. St. Paul, Minnesota: The Science Museum of Minnesota. p. 31
- ↑ Boyd 2014, p. 1-9
- ↑ a b c Norman, David B.; Sues, Hans-Dieter; Witmer, Larry M.; Coria, Rodolfo A. (2004). «Basal Ornithopoda». In: Weishampel, David B.; Dodson, Peter; Osmólska, Halszka. The Dinosauria 2nd ed. Berkeley: University of California Press. pp. 393–412. ISBN 978-0-520-24209-8
- ↑ a b Galton, P.M. (1997). «Cranial anatomy of the basal hypsilophodontid dinosaur Thescelosaurus neglectus Gilmore (Ornithischia: Ornithopoda) from the Late Cretaceous of North America». Revue de Paléobiologie. 16 (1): 231–258
- ↑ Boyd 2014, p. 18
- ↑ Boyd 2014, p. 20
- ↑ Boyd 2014, p. 55
- ↑ Boyd 2014, p. 6
- ↑ Fisher, P.E.; Russell, D.A.; Stoskopf, M.K.; Barrick, R.E.; Hammer, M.; Kuzmitz, A.A. (2000). «Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur». Science. 288 (5465): 503–505. Bibcode:2000Sci...288..503F. PMID 10775107. doi:10.1126/science.288.5465.503
- ↑ a b Boyd 2014, p. 7
- ↑ Horner, Jack (2001). Dinosaurs under the Big Sky. Missoula, Montana: Mountain Press Publications. p. 128. ISBN 9780878424450
- ↑ Boyd 2014, p. 8
- ↑ Butler, R.J.; Galton, P.M. (2008). «The 'dermal armour' of the ornithopod dinosaur Hypsilophodon from the Wealden (Early Cretaceous: Barremian) of the Isle of Wight: a reappraisal». Cretaceous Research. 29 (4): 636–642. Bibcode:2008CrRes..29..636B. doi:10.1016/j.cretres.2008.02.002
- ↑ a b c d e f Boyd, C.A. (2015). «The systematic relationships and biogeographic history of ornithischian dinosaurs». PeerJ. 3: e1523. PMC 4690359. PMID 26713260. doi:10.7717/peerj.1523
- ↑ Nopcsa, F. (1928). «The genera of reptiles». Palaeobiologica. 1: 163–188
- ↑ Huene, F.F.v. (1956). Paläontologie und Phylogenie der Niederen Tetrapoden. [S.l.]: Gustav Fischer Verlag. p. 536
- ↑ Rozhdestvensky, A.K. (1964). «Подотряд Ornithopoda, Птиценогие». In: Orlov, Y.A. Основы палеонтологии [Fundamentals of Paleontology]. [S.l.]: НАУКА. pp. 553–572
- ↑ Thulborn, R.A. (1974). «A new heterodontosaurid dinosaur (Reptilia: Ornithischia) from the Upper Triassic Red Beds of Lesotho». Zoological Journal of the Linnean Society. 55 (2): 151–175. doi:10.1111/j.1096-3642.1974.tb01591.x
- ↑ Romer, A.S. (1966). Vertebrate Paleontology 3rd ed. [S.l.]: The University of Chicago Press. p. 370
- ↑ Sues, H.-D.; Norman, D.B. (1990). «Hypsilophodontidae, Tenontosaurus, Dryosauridae». In: Weishampel, D.B.; Dodson, P.; Osmólska, H. The Dinosauria 1st ed. Berkeley: University of California Press. pp. 498–509. ISBN 978-0-520-06727-1
- ↑ Sereno, P.C. (1986). «Phylogeny of the Bird-Hipped Dinosaurs (Ornithischia)». National Geographic Research. 2 (2): 234–256
- ↑ Weishampel, D.B.; Heinrich, R.E. (1992). «Systematics of Hypsilophodontidae and Basal Iguanodontia (Dinosauria: Ornithopoda)». Historical Biology. 6 (3): 159–184. Bibcode:1992HBio....6..159W. doi:10.1080/10292389209380426
- ↑ Scheetz, R.D. (1999). «Osteology of Orodromeus makelai and the phylogeny of basal ornithopod dinosaurs». Unpublished Dissertation, Montana State University: 1–189
- ↑ Weishampel, D.B.; Jianu, C.-M.; Csiki, Z.; Norman, D.B. (2003). «Osteology and phylogeny of Zalmoxes (n.g.), an unusual euornithopod dinosaur from the latest Cretaceous of Romania». Journal of Systematic Palaeontology. 1 (2): 1–56. Bibcode:2003JSPal...1...65W. doi:10.1017/S1477201903001032
- ↑ Varricchio, D.J.; Martin, A.J.; Katsura, Y. (2007). «First trace and body fossil evidence of a burrowing, denning dinosaur». Proceedings of the Royal Society B: Biological Sciences. 274 (1616): 1361–1368. PMC 2176205. PMID 17374596. doi:10.1098/rspb.2006.0443
- ↑ a b Butler, R.J.; Upchurch, P.; Norman, D.B. (2008). «The phylogeny of the ornithischian dinosaurs». Journal of Systematic Palaeontology. 6 (1): 1–40. Bibcode:2008JSPal...6....1B. doi:10.1017/S1477201907002271
- ↑ a b Brown, C.M.; Evans, D.C.; Ryan, M.J.; Russell, A.P. (2013). «New data on the diversity and abundance of small-bodied ornithopods (Dinosauria, Ornithischia) from the Belly River Group (Campanian) of Alberta». Journal of Vertebrate Paleontology. 33 (3): 495–520. Bibcode:2013JVPal..33..495B. doi:10.1080/02724634.2013.746229
- ↑ Han, F.; Barrett, P.M.; Butler, R.J.; Xu, X. (2012). «Postcranial anatomy of Jeholosaurus shangyuanensis (Dinosauria, Ornithischia) from the Lower Cretaceous Yixian Formation of China». Journal of Vertebrate Paleontology. 32 (6): 1370–1395. Bibcode:2012JVPal..32.1370H. doi:10.1080/02724634.2012.694385
- ↑ a b c d Fonseca, A.O.; Reid, I.J.; Venner, A.; Duncan, R.J.; Garcia, M.S.; Müller, R.T. (2024). «A comprehensive phylogenetic analysis on early ornithischian evolution». Journal of Systematic Palaeontology. 22 (1). 2346577 páginas. Bibcode:2024JSPal..2246577F. doi:10.1080/14772019.2024.2346577
- ↑ a b Sues, H.-D.; Evans, D.C.; Galton, P.M.; Brown, C.M. (2023). «Anatomy of the neornithischian dinosaur Parksosaurus warreni (Parks, 1926) from the Upper Cretaceous (lower Maastrichtian) Horseshoe Canyon Formation of Alberta, Canada». Cretaceous Research. 141. Bibcode:2023CrRes.14105369S. doi:10.1016/j.cretres.2022.105369
- ↑ a b c Avrahami, H.M.; Makovicky, P.J.; Tucker, R.T.; Zanno, L.E. (2024). «A new semi-fossorial thescelosaurine dinosaur from the Cenomanian-age Mussentuchit Member of the Cedar Mountain Formation, Utah». The Anatomical Record (em inglês). 307 (12): 3717–3781. ISSN 1932-8486. doi:10.1002/ar.25505
- ↑ a b c Herne, M.C.; Nair, J.P.; Evans, A.R.; Tait, A.M. (2019). «New small-bodied ornithopods (Dinosauria, Neornithischia) from the Early Cretaceous Wonthaggi Formation (Strzelecki Group) of the Australian-Antarctic rift system, with revision of Qantassaurus intrepidus Rich and Vickers-Rich, 1999». Journal of Paleontology. 93 (3): 543–584. Bibcode:2019JPal...93..543H. doi:10.1017/jpa.2018.95
- ↑ a b Dieudonné, P.-E.; Cruzado-Caballero, P.; Godefroit, P.; Tortosa, T. (2021). «A new phylogeny of cerapodan dinosaurs». Historical Biology. 33 (10): 2335–2355. Bibcode:2021HBio...33.2335D. doi:10.1080/08912963.2020.1793979
- ↑ Hudgins, M.N.; Currie, P.J.; Sullivan, C. (2021). «Dental assessment of Stegoceras validum (Ornithischia: Pachycephalosauridae) and Thescelosaurus neglectus (Ornithischia: Thescelosauridae): paleoecological inferences». Cretaceous Research. 120. doi:10.1016/j.cretres.2021.105058
- ↑ Senter, Philip; Mackey, Jared (2023). «Forelimb motion and orientation in the ornithischian dinosaurs Styracosaurus and Thescelosaurus, and its implications for locomotion and other behavior». Palaeontologia Electronica (em inglês). doi:10.26879/1289
Bibliografia
- Boyd, C.A. (2014). «The cranial anatomy of the neornithischian dinosaur Thescelosaurus neglectus». PeerJ. 2: e669. PMC 4232843. PMID 25405076. doi:10.7717/peerj.669
Ligações externas
 O Wikispecies possui informações sobre: Thescelosaurus
O Wikispecies possui informações sobre: Thescelosaurus Media relacionados com Thescelosaurus no Wikimedia Commons
Media relacionados com Thescelosaurus no Wikimedia Commons
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||