Rhinesuchidae
Rhinesuchidae
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Guadalupiano-Triássico Inferior 279,3–247,2 Ma | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Gêneros | |||||||||||||
| |||||||||||||
| Sinónimos | |||||||||||||
| |||||||||||||
Rhinesuchidae é uma família de tetrápodes que viveu principalmente no período Permiano. Eles pertenciam ao amplo grupo Temnospondyli, uma coleção diversa e bem-sucedida de tetrápodes semiaquáticos dos quais os anfíbios modernos provavelmente descendem. Os membros da família Rhinesuchidae podem ser diferenciados de outros integrantes de Temnospondyli por detalhes de seus crânios, mais notavelmente a estrutura interna de seus entalhes óticos na parte posterior do crânio. Eles estavam entre os membros mais basais do grupo Stereospondyli, um subgrupo de Temnospondyli com cabeças achatadas e hábitos aquáticos. Embora os membros de Stereospondyli mais avançados tenham evoluído para alcançar uma distribuição mundial no Triássico, os membros da família Rhinesuchidae viveram principalmente em ambientes de alta latitude de Gondwana (hoje América do Sul e África) durante as épocas Guadalupiano e Lopingiano do Permiano. A taxonomia dessa família tem sido complicada, com mais de vinte espécies nomeadas no passado; uma revisão de 2017 reconheceu apenas oito delas (distribuídas entre sete gêneros) como válidas. Embora vários supostos membros desse grupo tenham sido relatados como vivendo no Triássico, a maioria é duvidosa ou não pertence ao grupo. No entanto, pelo menos um gênero válido é conhecido do Triássico Inferior, um pequeno membro chamado Broomistega [en]. A definição formal mais recente de Rhinesuchidae, defendida por Mariscano et al. (2017), é a de um clado contendo todos os táxons mais próximos de Rhinesuchus whaitsi [en] do que de Lydekkerina huxleyi [en] ou Peltobatrachus pustulatus [en].[1] Uma definição alternativa semelhante é que Rhinesuchidae é um clado contendo todos os táxons mais próximos de Uranocentrodon senekalensis [en] do que de Lydekkerina huxleyi, Trematosaurus brauni [en] ou Mastodonsaurus giganteus [en].[2]
Descrição
Os membros da família Rhinesuchidae geralmente tinham um tipo de corpo convencional para tetrápodes, com quatro membros e uma cauda moderadamente longa. Além disso, seus corpos eram um pouco alongados, e seus membros eram pequenos e fracos, mas ainda relativamente bem desenvolvidos. Alguns eram muito grandes, alcançando até 3 metros de comprimento. Como a maioria dos membros de Stereospondyli, seus crânios eram achatados e triangulares, com olhos voltados para cima. A maioria dos membros da família Rhinesuchidae tinha focinhos relativamente curtos, embora o focinho de Australerpeton [en] fosse muito longo e fino. Os únicos outros anfíbios gigantes de focinho longo do Permiano eram membros da família Archegosauridae, como Prionosuchus e Konzhukovia [en].
Entalhe ótico
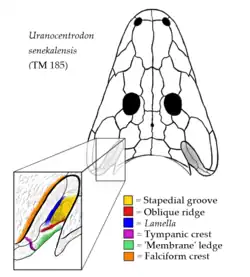
Como a maioria dos anfíbios antigos, os membros da família Rhinesuchidae tinham um par de indentações na borda posterior do crânio conhecidas como entalhes óticos. Embora às vezes consideradas como abrigando órgãos auditivos, como um tímpano (membrana timpânica), essas notas provavelmente continham espiráculos, orifícios carnudos usados para respiração. Os membros da família Rhinesuchidae podem ser caracterizados por um sistema único de cristas e sulcos dentro da cavidade interna de cada entalhe ótico. As paredes da cavidade do entalhe ótico (às vezes referidas como cavidade timpânica) são formadas principalmente pelo ramo ascendente dos ossos pterigoides. No entanto, a borda interna de cada cavidade é formada por um osso tabular. Os ossos tabulares são um par de ossos triangulares ao longo da borda posterior do crânio que formam estruturas pontiagudas conhecidas como chifres tabulares. A parte superior da parede externa da cavidade também é formada parcialmente pelos ossos esquamosais, que ocupam principalmente a face superior plana do crânio. A porção do esquamosal que forma a parede da cavidade é separada da porção externa da cavidade por um limite pronunciado conhecido como crista falciforme.
A parede externa da cavidade tem um sulco longo e pronunciado, conhecido como sulco estapédico, que se estende longitudinalmente ao longo da parede. A borda inferior do sulco é formada por uma crista conhecida como crista oblíqua. A borda superior do sulco estapédico é formada por outra crista que margeia o osso esquamosal, que Eltink et al. (2016) nomearam 'crista pterigoide dorsal'.[3] No entanto, Mariscano et al. (2017) preferiram usar o nome "lamella" para essa estrutura, para que não fosse confundida com uma crista diferente presente em membros da família Lydekkerinidae, que às vezes é chamada de "crista oblíqua do pterigoide", mas mais comumente chamada de "crista timpânica".[1][3] Confusamente, muitos membros da família Rhinesuchidae também possuem uma crista timpânica. Essa crista estava posicionada mais atrás do que as outras cristas (perto da interseção dos ossos pterigoide, quadrado e esquamosal) e se estende para baixo ao longo da face posterior da bochecha. A borda interna da parede externa da cavidade era formada por uma borda que a maioria dos estudos simplesmente chama de "membrana". Essa convenção existe devido à antiga e provavelmente incorreta hipótese de que os entalhes óticos abrigavam tímpanos.[4] Sob essa hipótese, a borda interna pode ter se fixado a uma membrana que se estendia pela cavidade interna do ouvido.[3]
Essa combinação de sulcos e cristas na cavidade ótica é única para os membros da família. A lamella e o sulco estapédico são desconhecidos em outros grupos, embora estejam presentes em praticamente todos os integrantes de Rhinesuchidae (exceto Broomistega, que não possui lamella). A crista timpânica está presente na maioria dos integrantes, mas ausente em alguns, e também está presente na família Lydekkerinidae. A crista oblíqua e a crista falciforme estão presentes na maioria dos outros membros de Stereospondyli (embora a primeira seja menos desenvolvida), enquanto a borda de "membrana" está presente em praticamente todos os membros de Stereospondylomorpha [en].[3]
Palato e caixa craniana
Vários ossos e aberturas compunham o palato (teto da boca) nos membros da família Rhinesuchidae, como em outros anfíbios. Na ponta do palato estava o vômer, enquanto as áreas próximas à borda da boca eram feitas de ossos palatinos e ectopterigoides. No meio da parte posterior da boca havia um osso retangular conhecido como paraesfenóide. A maior parte do paraesfenóide formava a face inferior da caixa craniana achatada, embora também possuísse uma haste fina projetada para a frente, conhecida como processo cultriforme, que se estende ao longo da linha mediana do crânio até encontrar o vômer. Na parte posterior da boca, havia os ossos pterigoides com várias ramificações em cada lado do crânio. Cada pterigoide tinha vários ramos, incluindo o ramo posterior, que se estende para trás e para o lado do crânio, o ramo medial curto, que se estende para dentro e se conecta ao osso paraesfenóide, um ramo ascendente que se projeta para cima para formar o entalhe ótico, e finalmente o ramo anterior, que se estende para a frente ao longo do palatino e ectopterigoide. Os pterigoides da maioria dos membros da família Rhinesuchidae têm ramos anteriores muito longos. Na maioria dos membros dessa família, o ramo anterior vai até o vômer, embora Australerpeton tenha ramos anteriores relativamente curtos. Um par de grandes aberturas preenche as áreas entre esses ossos, tornando a maior parte do palato um espaço aberto.[1][3]
Quando visto de trás, os ramos superiores da caixa craniana (processos paraoccipitais) se estendem de lado a lado, ocultando parcialmente o ramo ascendente dos pterigoides. Cada processo paraoccipital também é perfurado por um pequeno orifício, conhecido como fenestra pós-temporal. Esses orifícios são muito finos nos membros da família Rhinesuchidae. Acima desses processos paraoccipitais estão os entalhes óticos e os ossos tabulares. Os processos paraoccipitais também apontam para trás em certa medida, formando chifres que, em alguns membros da família Rhinesuchidae, são ligeiramente mais longos que os dos tabulares. Quando visto de baixo, a porção mais proeminente da caixa craniana é o osso paraesfenóide. Os cantos posteriores do paraesfenóide têm pequenos "bolsos" delimitados por cristas (conhecidas como crista muscularis).[1] Essas cristas podem ter ancorado músculos capazes de movimentar a cabeça no pescoço.[4]
Outras características do crânio e da mandíbula
Muitos ossos formavam a parte superior do crânio, embora um par específico de ossos tivesse um design particular nos membros da família Rhinesuchidae. Esses ossos eram os jugais e pré-frontais alongados, que formavam a borda anterior das órbitas (orifícios dos olhos). Na maioria dos membros da família, a borda entre os dois ossos tinha uma forma "escalonada", com uma extensão triangular externa do pré-frontal empurrando a sutura com o jugal para uma posição mais lateral (para fora). No entanto, a sutura é mais reta em Australerpeton, como em outros Stereospondyli.[1]
A mandíbula inferior tem um par de orifícios visíveis apenas na borda interna da mandíbula. O orifício maior na parte posterior do complexo ósseo, conhecido como sulco meckeliano, era fino e alongado nos membros da família Rhinesuchidae. Um orifício adicional na parte inferior da articulação da mandíbula é visível apenas de baixo. Esse orifício, o forame da corda timpânica, era grande nessa família. Na parte superior da articulação da mandíbula, um sulco fino conhecido como sulco arcadiano se estende em direção ao lado lingual (língua) da mandíbula e separa outros relevos ósseos localizados entre a articulação da mandíbula. No geral, os sulcos e cristas da articulação da mandíbula eram pouco desenvolvidos nos membros da família Rhinesuchidae em comparação com muitos outros grupos de Stereospondyli, assemelhando-se à articulação simples de Archegosauridae, como Melosaurus [en].[1]
Paleobiologia
A maioria dos membros da família Rhinesuchidae é conhecida apenas por material craniano, embora alguns membros do grupo (Uranocentrodon, Broomistega e Australerpeton, por exemplo) incluam espécimes que preservam uma porção significativa do resto do esqueleto. Um espécime filhote de Broomistega tinha tornozelos e vértebras pouco ossificados, indicando que suas articulações tinham uma grande quantidade de material cartilaginoso para complementar a baixa quantidade de osso. Esse traço é frequentemente correlacionado com um estilo de vida aquático.[5] Características do crânio, como olhos voltados para cima, também apoiam essa hipótese.
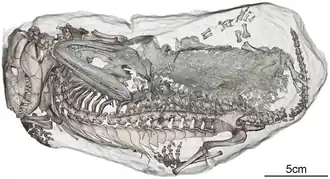
Apesar desse suporte para um estilo de vida aquático, outras evidências mostram que os membros da família Rhinesuchidae eram capazes de algum movimento terrestre. Embora os membros da família Rhinesuchidae não possuíssem adaptações para cavar, o espécime juvenil pouco ossificado de Broomistega foi encontrado em uma toca inundada que também era habitada por um Thrinaxodon. Várias condições da forma como esses animais foram preservados indicam que eles coabitavam a toca pacificamente, provavelmente para sobreviver a uma seca por estivação (permanecendo em estado dormente durante condições quentes e secas). O fato de um espécime de Broomistega ter conseguido entrar na toca de um animal terrestre como Thrinaxodon indica que os membros da família Rhinesuchidae não eram exclusivamente aquáticos.[5]
Além disso, foi observado que espécimes maiores do grupo Temnospondyli geralmente têm articulações mais bem ossificadas. Por exemplo, grandes espécimes de Australerpeton possuíam quadris robustos, vários ossos de tornozelo completamente ósseos e pleurocentros ossificados (parte das vértebras).[6] No entanto, esses esqueletos não eram tão robustos quanto os de Eryops (supostamente terrestre), com cinturas escapulares menores e locais menos proeminentes para fixação muscular. Dias & Schultz (2003) sugeriram que o estilo de vida de Australerpeton (e presumivelmente outros membros da família Rhinesuchidae) era o de um ictiófago semiaquático, preferindo caçar em corpos d'água rasos, mas mantendo a capacidade de caminhar em terra durante secas.[7]
Um estudo de histologia de vários fósseis indeterminados de membros da família Rhinesuchidae (referidos a Rhinesuchus) indica que os membros da família cresciam sazonalmente, como os anfíbios modernos. Os indivíduos também tinham uma vida relativamente longa, com um espécime tendo de 30 a 35 anos na época de sua morte, com base no número de linhas de crescimento interrompido (anéis nos ossos usados para determinar a idade, como anéis de árvores) presentes em um fragmento de quadril. Algumas linhas de crescimento interrompido eram muito estreitas, indicando que os indivíduos podiam reduzir seu crescimento e metabolismo durante períodos de dificuldade. Essa capacidade pode ser a razão pela qual os membros da família Rhinesuchidae foram relativamente bem-sucedidos no final do Permiano, bem como por que alguns membros pequenos do grupo sobreviveram ao evento de extinção do Permiano-Triássico.[8]
Brânquias
Três fileiras de ossos minúsculos (ossículos branquiais) cobertos com estruturas finas semelhantes a dentes foram preservadas perto do pescoço de um espécime de Uranocentrodon. Esses ossos quase certamente estavam fixados aos arcos branquiais das brânquias enquanto o animal estava vivo.[9] Embora tais ossos sejam raros entre os integrantes de Stereospondyli e desconhecidos em outros membros da família Rhinesuchidae, isso pode ser simplesmente devido ao fato de que os ossos de outros gêneros foram preservados em sedimentos de grão mais grosseiro, onde ossos tão delicados poderiam ser quebrados ou difíceis de encontrar.[10]
Embora seja evidente que Uranocentrodon tinha brânquias de algum tipo, é difícil determinar que tipo de brânquias eram. Por um lado, poderiam ser brânquias internas, como as dos peixes, que mal eram visíveis do exterior do corpo. Por outro lado, poderiam ser brânquias externas em forma de haste, como as de larvas de salamandras modernas ou até mesmo salamandras adultas neotênicas, como espécies de Necturus ou o axolote. As brânquias externas devem ter evoluído a partir das brânquias internas em algum momento durante a evolução dos anfíbios, embora a localização precisa dessa transição seja controversa. Os ossos de suporte às brânquias preservados em anfíbios antigos mostram muitas semelhanças com os das brânquias de peixes e de salamandras. Paleontólogos que preferem comparar tetrápodes antigos com anfíbios modernos geralmente encontram muitas semelhanças entre os ossos fósseis e os ossos branquiais de salamandras modernas. Por outro lado, paleontólogos que comparam tetrápodes fósseis com peixes fósseis consideram que os ossos se correlacionam com brânquias internas. Esse dilema, conhecido como paradoxo de Bystrow, dificultou a avaliação das brânquias em anfíbios antigos como Uranocentrodon, pois diferentes paleontólogos chegam a conclusões diferentes com base em seu campo de estudo.[11]

O paradoxo de Bystrow foi finalmente resolvido por um estudo de 2010, que descobriu que estruturas ceratobranquiais sulcadas (componentes dos arcos branquiais) estão correlacionadas com brânquias internas. Os tetrápodes antigos que preservaram estruturas ceratobranquiais sulcadas, como Dvinosaurus [en], provavelmente tinham apenas brânquias internas como adultos. No entanto, brânquias externas foram diretamente preservadas como tecido mole em alguns membros de Temnospondyli. Contudo, essas situações ocorrem apenas em espécimes larvais ou membros de grupos especializados, como espécies da família Branchiosauridae [en]. Uma espécie viva de peixe dipnoico (Lepidosiren) tem brânquias externas como larva, que se transformam em brânquias internas como adulto. Apesar de espécimes adultos de Dvinosaurus terem características esqueléticas correlacionadas com brânquias internas, alguns espécimes larvais de outro Dvinosaurus, Isodectes [en], preservaram brânquias externas de tecido mole. Assim, o desenvolvimento das brânquias de Dvinosaurus (e presumivelmente de outros membros de Temnospondyli, como Uranocentrodon) espelhava o de Lepidosiren. Apesar de essa característica provavelmente ser um exemplo de evolução convergente (já que outros peixes dipnoicos possuem exclusivamente brânquias internas), ela ainda é uma referência útil para como as brânquias dos membros de Temnospondyli se desenvolveram. Os autores do estudo concluíram que as brânquias de Temnospondyli (incluindo Uranocentrodon e outros membros da família Rhinesuchidae que podem ter possuído brânquias) eram provavelmente internas (como as de um peixe) na fase adulta, mas externas (como as de uma salamandra) na fase larval.[11]
Armadura corporal
Um esqueleto de Uranocentrodon também preservou grandes áreas de escudos ou escamas ósseas ao redor do corpo. Os escudos que estariam na barriga do animal estavam dispostos em fileiras diagonais paralelas que convergiam na linha mediana do corpo e divergiam à medida que as fileiras se estendiam em direção à cauda. Cada escudo tinha uma crista ao longo da parte central, e os escudos mais próximos da linha mediana se sobrepunham aos mais afastados. Ao longo da linha mediana, uma fileira de escamas planas e largas se estendia da garganta até a cauda. Enquanto essas escamas ventrais eram feitas de osso, as escamas em outras partes do corpo tinham menos estrutura óssea e provavelmente eram feitas de queratina. As escamas nas laterais do corpo eram mais planas e menores que os escudos ventrais ósseos. Os escudos na parte traseira do corpo eram semelhantes, embora mais arredondados, com alguns escudos maiores perto da linha mediana. As escamas dos membros posteriores e da parte inferior da região do quadril eram semelhantes às da parte traseira, embora nenhum tegumento tenha sido preservado nos membros anteriores ou na cauda. Assim, é provável que pelo menos a cauda não fosse blindada e estivesse coberta apenas por pele nua.[9]
Escamas também foram preservadas em espécimes de Australerpeton. Elas são semelhantes em distribuição às de Uranocentrodon, mas geralmente são mais arredondadas. Também possuíam uma estrutura interna em forma de favo e características histológicas que indicam que estavam profundamente embutidas na pele. Portanto, é improvável que fossem visíveis do exterior do corpo. Não pode ser determinado se as escamas ou escudos dos membros da família Rhinesuchidae permitiam ou restringiam a respiração cutânea (respiração pela pele, como em anfíbios modernos). Outras aplicações potenciais das escamas incluíam proteção contra predadores, retenção de água durante secas e possivelmente até armazenamento de cálcio em condições adversas (uma técnica usada por fêmeas de crocodilos-do-nilo). Essa última hipótese é a menos provável, pois os membros da família Rhinesuchidae não botavam ovos de casca dura, que é a razão pela qual as crocodilos fêmeas precisam armazenar cálcio.[12]
Classificação
Quando a família foi nomeada pela primeira vez em 1919, Rhinesuchidae já era reconhecida como um grupo basal, uma posição que mantém até hoje. Entre os traços usados para apoiar essa posição está o fato de que a maioria dos membros da família Rhinesuchidae tinha ramos anteriores longos de seus pterigoides. Grupos mais avançados tinham ramos anteriores mais curtos. Em 1947, Alfred Sherwood Romer colocou a família (que ele acreditava incluir apenas Rhinesuchus) em uma ampla superfamília que ele chamou de Rhinesuchoidea.[13] Rhinesuchoidea foi concebida como parte de um grado de Temnospondyli que ligava "primitivos" do grupo Rhachitomi [en], como Eryops, a Stereospondyli "avançados", como os das famílias Metoposauridae [en] e Trematosauridae [en]. Esse grado foi separado em Capitosauroidea (que continha o grupo Capitosauria [en] e o gênero Benthosuchus [en]) e Rhinesuchoidea. Além de conter Rhinesuchidae, Rhinesuchoidea também incluía vários gêneros, bem como as famílias Lydekkerinidae [en], Sclerothoracidae e, finalmente, Uranocentrodontidae. Romer sentiu que certos táxons (isto é, Uranocentrodon e o gênero duvidoso possivelmente sinônimo "Laccocephalus"[1]) frequentemente considerados membros da família Rhinesuchidae eram melhor colocados na família separada Uranocentrodontidae, enquanto outros (isto é, Rhinesuchoides) não foram colocados em nenhuma família em particular.[13] Outras famílias foram posteriormente colocadas em Rhinesuchoidea, como Rhinecepidae em 1966 e Australerpetonidae em 1998.[1]
A chegada da cladística no final do século XX fez com que os grados caíssem em desuso nos últimos anos, sendo substituídos por clados, que são definidos por relações próximas em vez de associações ancestrais. No entanto, a ideia básica por trás de Rhinesuchoidea, que afirma que Stereospondyli avançados descendem de animais semelhantes aos membros da família Rhinesuchidae, ainda é considerada válida. Rhinecepidae e Uranocentrodontidae foram consideradas sinônimas de Rhinesuchidae, segundo uma análise de 2000 por Schoch e Milner. Um estudo colocou Rhinesuchidae dentro da superfamília Capitosauroidea.[14] No entanto, essa interpretação não foi seguida por outros estudos que consideram os membros da família Rhinesuchidae mais basais que Capitosauria. Australerpetonidae, uma família monotípica contendo apenas o gênero Australerpeton, tem sido mais difícil de comparar com Rhinesuchidae. Alguns estudos colocam Australerpeton como Stereospondyli basal fora de Rhinesuchidae,[2][6] enquanto outros o consideram um membro de Archegosauridae fora de Stereospondyli completamente.[15]
Uma revisão abrangente de Australerpeton publicada por Eltink et al. (2016) favoreceu a hipótese de que ele estava profundamente aninhado dentro de Rhinesuchidae. Um estudo filogenético realizado como parte do estudo dividiu a família em dois clados. Um clado era uma subfamília chamada Rhinesuchinae. Rhinesuchinae contém Rhinesuchus e Rhineceps. Essa subfamília é caracterizada principalmente por características do palato, como um ramo anterior do pterigoide sem cristas e ossos palatinos cobertos por pequenos dentículos. O outro clado principal da família continha Uranocentrodon, bem como outra subfamília chamada Australerpetinae. Esse clado é unido pela presença de uma crista timpânica e um forame magno (o orifício para a medula espinhal na parte posterior da caixa craniana) que tem uma borda superior curvada. Australerpetinae é uma versão modificada de Australerpetonidae, que foi reduzida ao status de subfamília para se encaixar dentro de Rhinesuchidae. Essa subfamília contém Australerpeton, Broomistega, Laccosaurus e Rhinesuchoides. Membros dessa subfamília tinham focinhos um pouco mais longos e afilados que outros membros da família Rhinesuchidae, embora (segundo Eltink et al.) seus pterigoides tivessem ramos anteriores curtos. A árvore mais parcimoniosa (evolutivamente mais simples) encontrada por Eltink et al. (2016) é vista abaixo:[3]
| Temnospondyli |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
A estrutura de Rhinesuchidae após o estudo de Eltink et al. foi contestada por um estudo diferente sobre membros da família Rhinesuchidae publicado menos de um ano depois. Esse estudo, Mariscano et al. (2017), concordou que Australerpeton era um integrante de Rhinesuchidae, mas o considerou o membro mais basal da família. Eles discordam do reconhecimento de Eltink et al. de ramos anteriores curtos do pterigoide em vários gêneros. Segundo sua análise, apenas Australerpeton possuía esse traço, a principal característica que o separa do resto de Rhinesuchidae. Outros traços que apoiam essa separação incluem o fato de que outros membros da família Rhinesuchidae têm contato jugal-pré-frontal escalonado e ossos coronoides sem dentes na mandíbula inferior. O resto da família foi pouco resolvido em sua análise filogenética, embora três clados tivessem valores de suporte Bremer moderados de 2.
O suporte Bremer é medido contando o número de vezes que os traços analisados são adquiridos, perdidos ou readquiridos dentro de uma árvore genealógica. Algumas árvores genealógicas incluem mais dessas transições do que outras, o que significa que algumas árvores possíveis assumem que mais do que a quantidade mínima de evolução ocorreu. A árvore genealógica com o menor número dessas "etapas" (transições) é provavelmente a mais precisa, com base no princípio da navalha de Occam (a resposta mais simples é a mais precisa). O suporte Bremer é usado para indicar o quão bem suportados são os clados, analisando como eles são distribuídos entre alternativas mais complexas à árvore mais parcimoniosa. Clados que não existem em uma árvore genealógica que é apenas uma etapa mais complexa que a árvore mais parcimoniosa têm um suporte Bremer de 1, significando que a existência do clado é muito incerta. Mesmo que a árvore mais parcimoniosa da análise atual apoie sua existência, novos dados podem tornar uma árvore genealógica concorrente mais parcimoniosa, dissolvendo clados que são suportados apenas na árvore mais parcimoniosa atual. Outros clados podem ter valores de suporte Bremer muito mais altos, indicando que suposições mais drásticas precisam ser formuladas para tornar o clado inválido. Rhinesuchidae como um todo, por exemplo, tem um suporte Bremer de 6 em Mariscano et al. (2017), que é considerado alto suporte. Um suporte Bremer de 2, como é o caso de três clados específicos nesta análise, é considerado moderado. Um desses clados incluía as duas espécies válidas de Rhinesuchoides, enquanto outro clado conectava Rhineceps e Uranocentrodon, e o último continha Rhinesuchus e Laccosaurus. A disposição desses clados (bem como a colocação de Broomistega) não pôde ser resolvida com confiança absoluta, com valores de suporte Bremer de apenas 1, independentemente de onde os três clados foram colocados entre os Rhinesuchidae não-Australerpeton. A árvore mais parcimoniosa encontrada por Mariscano et al. (2017) é vista abaixo:[1]
| Temnospondyli |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Galeria
-
![Rhinesuchus whaitsi [en], do Permiano Médio (Guadalupiano) ao Permiano Superior (Lopinguiano) da África do Sul.](./_assets_/0c70a452f799bfe840676ee341124611/Rhinesuchus1DB.jpg) Rhinesuchus whaitsi [en], do Permiano Médio (Guadalupiano) ao Permiano Superior (Lopinguiano) da África do Sul.
Rhinesuchus whaitsi [en], do Permiano Médio (Guadalupiano) ao Permiano Superior (Lopinguiano) da África do Sul. -
-
![Uranocentrodon senekalensis [en], do Permiano Superior da África do Sul.](./_assets_/0c70a452f799bfe840676ee341124611/Uranocentr10_copy.jpg) Uranocentrodon senekalensis [en], do Permiano Superior da África do Sul.
Uranocentrodon senekalensis [en], do Permiano Superior da África do Sul. -
![Rhineceps nyasaensis [en], do Permiano Superior do Malawi.](./_assets_/0c70a452f799bfe840676ee341124611/Rhineceps_nyasaensis.jpg)
-
![Broomistega putterilli [en], do Triássico inferior da África do Sul.](./_assets_/0c70a452f799bfe840676ee341124611/Broomistega_putterilli.jpg) Broomistega putterilli [en], do Triássico inferior da África do Sul.
Broomistega putterilli [en], do Triássico inferior da África do Sul.
Referências
- ↑ a b c d e f g h i Mariscano, C.A.; Latimer, E.; Rubidge, B.; Smith, R.M.H. (2017). «The Rhinesuchidae and early history of the Stereospondyli (Amphibia: Temnospondyli) at the end of the Palaeozoic». Zoological Journal of the Linnean Society. 181 (2): 357–384. doi:10.1093/zoolinnean/zlw032. hdl:11336/105150

- ↑ a b Schoch, Rainer R. (2013). «The evolution of major temnospondyl clades: an inclusive phylogenetic analysis». Journal of Systematic Palaeontology. 11 (6): 673–705. Bibcode:2013JSPal..11..673S. doi:10.1080/14772019.2012.699006
- ↑ a b c d e f Eltink, Estevan; Dias, Eliseu V.; Dias-da-Silva, Sérgio; Schultz, Cesar L.; Langer, Max C. (2016). «The cranial morphology of the temnospondyl Australerpeton cosgriffi (Tetrapoda: Stereospondyli) from the Middle-Late Permian of Paraná Basin and the phylogenetic relationships of Rhinesuchidae». Zoological Journal of the Linnean Society (em inglês). 176 (4): 835–860. ISSN 0024-4082. doi:10.1111/zoj.12339
- ↑ a b Watson, D.M.S. (1962). «The Evolution of the Labyrinthodonts». Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 245 (723): 219–265. Bibcode:1962RSPTB.245..219W. JSTOR 2416605. doi:10.1098/rstb.1962.0010
- ↑ a b Fernandez, Vincent; Abdala, Fernando; Carlson, Kristian J.; Cook, Della Collins; Rubidge, Bruce S.; Yates, Adam; Tafforeau, Paul (2013). «Synchrotron Reveals Early Triassic Odd Couple: Injured Amphibian and Aestivating Therapsid Share Burrow». PLOS ONE (em inglês). 8 (6): e64978. Bibcode:2013PLoSO...864978F. ISSN 1932-6203. PMC 3689844. PMID 23805181. doi:10.1371/journal.pone.0064978
- ↑ a b Eltink, Estevan; Langer, Max C. (2014). «A New Specimen of the Temnospondyl Australerpeton cosgriffi from the Late Permian of Brazil (Rio Do Rasto Formation, Paraná Basin): Comparative Anatomy and phylogenetic relationships». Journal of Vertebrate Paleontology. 34 (3): 524–538. Bibcode:2014JVPal..34..524E. JSTOR 24523275. doi:10.1080/02724634.2013.826667
- ↑ Dias, E.V.; Schultz, C.L. (2003). «The first Paleozoic temnospondyl postcranial skeleton from South America» (PDF). Revista Brasileira de Paleontologia. 6: 29–42. Consultado em 19 de junho de 2025
- ↑ McHugh, Julia B. (2014). «Paleohistology and histovariability of the Permian stereospondyl Rhinesuchus». Journal of Vertebrate Paleontology. 34 (1): 59–68. Bibcode:2014JVPal..34...59M. doi:10.1080/02724634.2013.787429. Consultado em 19 de junho de 2025
- ↑ a b Van Hoepen, E.C.N. (1915). «Stegocephalia of Senekal». Annals of the Transvaal Museum. 5 (2): 125–149. Consultado em 19 de junho de 2025
- ↑ Schoch, Rainer R. (2002). «The evolution of metamorphosis in temnospondyls». Lethaia (em inglês). 35 (4): 309–327. ISSN 1502-3931. doi:10.1111/j.1502-3931.2002.tb00091.x
- ↑ a b Schoch, Rainer R.; Witzmann, Florian (2011). «Bystrow's Paradox - gills, fossils, and the fish-to-tetrapod transition». Acta Zoologica (em inglês). 92 (3): 251–265. ISSN 1463-6395. doi:10.1111/j.1463-6395.2010.00456.x. Consultado em 19 de junho de 2025
- ↑ Dias, E.V.; Richter, M. (2002). «On the squamation of Australerpeton cosgriffi Barberena, a temnospondyl amphibian from the Upper Permian of Brazil» (PDF). Anais da Academia Brasileira de Ciências. 74 (3): 477–490. ISSN 0001-3765. doi:10.1590/S0001-37652002000300010. Consultado em 19 de junho de 2025
- ↑ a b Romer, Alfred Sherwood (1947). «Review of the Labyrinthodontia». Bulletin of the Museum of Comparative Zoology at Harvard College. 99 (1): 7–368. Consultado em 19 de junho de 2025
- ↑ Shishkin, M. A.; Rubidge, B. S. (2000). «A Relict Rhinesuchid (Amphibia: Temnospondyli) From The Lower Triassic Of South Africa». Palaeontology (em inglês). 43 (4): 653–670. Bibcode:2000Palgy..43..653S. ISSN 1475-4983. doi:10.1111/1475-4983.00144
- ↑ Schoch, R. R., and Milner, A. R. 2000. Stereospondyli, stem-Stereospondyli, Rhinesuchidae, Rhytidostea, Trematosauroidea, Capitosauroidea. Handbuch der Paläoherpetologie, Munich, 3b:1-203.