Reparação de incompatibilidades do ADN
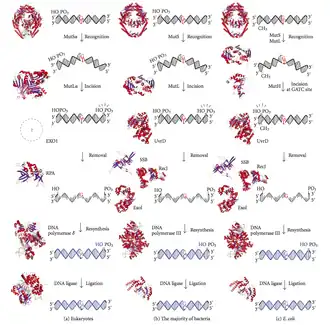
A reparação de incompatibilidades no ADN (abreviada como MMR, do inglês mismatch repair) é um sistema para o reconhecimento e reparação da inserção errada, deleção, ou incorreta incorporação de bases (sem respeitar a complementariedade), que se pode originar durante a replicação do ADN e a recombinação, e também para a reparação de algumas formas de danos no ADN.[1][2]
A reparação de incompatibilidades é específica de fita. Durante a síntese de ADN a fita nova sintetizada (fita filha) incluirá erros. Para começar a reparar-se, a maquinaria de reparação distingue a fita filha de nova síntese da fita molde (parental). Em bactérias gramnegativas, a hemimetilação transitória distingue as duas fitas, já que a parental está metilada e a filha não. Porém, noutros procariotas e eucariotas, o mecanismo exacto não está claro. Suspeita-se que, em eucariotas, a fita atrasada do ADN de nova síntese contém transitoriamente amossegas (ou rompimento de fita simples) (antes de que sejam seladas pela ADN ligase), o que proporciona um sinal que dirige os sistemas de correção de provas de incompatibilidades para a fita apropriada. Isto implica que estas amossegas devem estar presentes na fita guia, e encontraram-se recentemente provas sobre isto.[3] Em trabalhos recentes[4] demonstrou-se que as amossegas são sítios para a carga dependente de RFC da braçadeira deslizante de replicação PCNA, de uma maneira específica de orientação, de tal maneira que uma das faces da proteína com forma de donut se justapõe para o extremo 3'-OH na amossegas. A PCNA orientada dirige depois a ação da endonuclease MutLalpha para uma fita em presença de uma incompatibilidade e MutSalpha ou MutSbeta.
Qualquer evento mutacional que distorça a estrutura de super-hélice do ADN tem o potencial de comprometer a estabilidade genética da célula. O facto de que os sistemas de reparação e deteção de danos sejam tão complexos quanto a própria maquinaria de replicação sublinha a importância que na evolução se lhe deu à fidelidade na cópia do ADN.
Exemplos de bases discordantes são os emparelhamentos G/T ou A/C, que não respeitam o emparelhamento normal de bases (ver reparação do ADN). As incompatibilidade devem-se comunmente à tautomerização de bases durante a fase G2 do ciclo celular. Os danos são reparados por reconhecimento da deformidade causada pela incompatibilidade, o que determina as fitas molde e não molde, e excinde a base incorporada incorretamente e a substitua com o nucleótido correto. O processo de eliminação envolve geralmente mais nucleótidos do que o próprio nucleótido incompatibilidade. Podem eliminar-se junto com ele alguns pares de bases ou às vezes até milhares de pares de bases da fita de ADN de nova síntese.
Referências
- ↑ Iyer R, Pluciennik A, Burdett V, Modrich P (2006). «DNA mismatch repair: functions and mechanisms». Chem Rev. 106 (2): 302–23. PMID 16464007. doi:10.1021/cr0404794
- ↑ Larrea AA, Lujan SA, Kunkel TA (2010). «DNA mismatch repair». Cell. 141 (4). 730 páginas. PMID 20478261. doi:10.1016/j.cell.2010.05.002
- ↑ Heller RC, Marians KJ (2006). «Replisome assembly and the direct restart of stalled replication forks». Nat Rev Mol Cell Biol. 7 (12): 932–43. PMID 17139333. doi:10.1038/nrm2058
- ↑ Pluciennik; et al. (2010). «PCNA function in the activation and strand direction of MutLα endonuclease in mismatch repair.». PNAS. 107 (37): 16066–71. PMC 2941292
 . PMID 20713735. doi:10.1073/pnas.1010662107
. PMID 20713735. doi:10.1073/pnas.1010662107