Predação de ovos

A predação de ovos ou ovivoria é uma estratégia de alimentação em diversos grupos de animais (ovívoros) na qual eles consomem ovos. Como um ovo fertilizado representa um organismo completo em uma etapa de seu ciclo de vida, comer um ovo é uma forma de predação, ou seja, a eliminação de outro organismo para obtenção de alimento.
A predação de ovos é amplamente observada no reino animal, incluindo peixes, aves, serpentes, mamíferos e artrópodes. Algumas espécies são predadoras especializadas de ovos, enquanto muitas outras são generalistas, consumindo ovos quando a oportunidade surge.
Os seres humanos introduziram, acidentalmente ou intencionalmente, predadores de ovos, como ratos, em locais antes livres desses animais, causando prejuízos a espécies nativas, como aves marinhas que nidificam no solo. Aves predadoras, como corvos e gaivotas, também se expandiram, ameaçando aves que nidificam no solo como o tetrazes [en] e os esternídeos. Medidas para controlar esses predadores incluem o uso de iscas envenenadas.
Definições
Um animal ovívoro é aquele que se alimenta de ovos, termo derivado do latim ovum (ovo) e vorare (devorar).[1] Um ovívoro obrigatório ou predador exclusivo de ovos é um animal que se alimenta exclusivamente de ovos.[2] Isso difere de um parasita de ovo, como uma vespa parasitoide, que se desenvolve dentro do ovo de outro inseto.[3]
Relação ecológica
A predação de ovos é uma relação ecológica na qual um animal (predador) caça e consome os ovos de outra espécie (presa). Isso reduz a aptidão evolutiva dos pais cujos ovos são predados.[4]
Predadores generalistas de ovos

Os predadores generalistas podem ter um efeito substancial sobre as aves que fazem ninhos no solo, como a tarambola-dourada, Pluvialis apricaria: na Noruega, 78,2% dos ninhos dessa espécie sofreram predação. A remoção experimental de dois predadores de ninhos e ovos, a raposa-vermelha e a gralha-preta, aumentou a porcentagem de casais que conseguiram criar filhotes de cerca de 18% para cerca de 75%. O aumento populacional de predadores generalistas, como gaviões, texugos, corvos, martas, corvos-marinhos e raposas-vermelhas na Escócia, contribuiu para o declínio de várias espécies de aves que nidificam no solo, ao consumirem ovos, filhotes e fêmeas chocando.[6]
Comportamentos aprendidos para predação de ovos
Corvídeos, como corvos, são inteligentes e capazes de desenvolver comportamentos novos de forrageamento. No século XXI, corvos-pequenos aprenderam a saquear ninhos de pinguins-azuis na Ilha Phillip, no sudeste da Austrália, para acessar seus ovos. Cerca de um quarto dos ataques ocorre pela entrada do ninho (apenas em tocas curtas); o restante é feito por meio de buracos cavados no teto da toca. Os corvos predaram 61% dos ninhos monitorados.[7]
_(detail).jpg)
A primatóloga Jane Goodall observou que algumas aves e mamíferos utilizam ferramentas para quebrar ovos. Os abutres-do-egito jogam ovos pequenos para quebrá-los e atiram pedras em ovos de avestruz, que são grandes demais para serem levantados. Algumas espécies de mangustos jogam ovos contra rochas ou os pegam e os deixam cair sobre rochas.[8]
Predadores especializados de ovos
Algumas serpentes são especializadas na predação de ovos, como Oligodon formosanus [en], a serpente-marinha-marmoreada (Aipysurus eydouxii [en]), as cobras do gênero Dasypeltis [en] e a Boiga westermanni [en].[9][10][11][12] Essas serpentes possuem adaptações para sua dieta, como dentes e glândulas de peçonha atrofiados, que não são mais necessários para capturar presas (embora os poucos dentes de B. westermanni ainda sejam usados para segurar os ovos ao engoli-los).[10][11] A serpente-marinha-marmoreada também apresenta uma mutação de deleção em seu gene de toxina de três dedos [en], reduzindo a toxicidade de seu veneno entre 50 e 100 vezes.[9][10] Nas cobras africanas e indianas comedoras de ovos, há hipapófises (protrusões) nas vértebras usadas para quebrar os ovos engolidos.[11][12]
Entre os invertebrados, a sanguessuga aquática Cystobranchus virginicus é uma predadora de ovos. Pode ser uma alimentadora obrigatória de ovos, pois não foi observada se alimentando de adultos, mas foi encontrada nos ninhos de várias espécies de peixes de água doce norte-americanos dos gêneros Campostoma e Moxostoma.[13] Uma espécie de tripes, Mirothrips arbiter, do Brasil, é uma predadora obrigatória de ovos; ela se reproduz em colônias de vespas (Polistinae), e tanto suas larvas quanto seus adultos se alimentam dos ovos das vespas.[14]
Estratégias contra a predação de ovos
_%E9%95%B7%E5%B0%BE%E5%8D%97%E8%9C%A52.jpg)
A teoria da seleção r/K implica duas estratégias amplas para sobreviver à predação: reproduzir-se rapidamente (estrategistas r) para que os predadores não eliminem a presa; ou fornecer cuidado suficiente (estrategistas K) a um número menor de filhotes para que alguns sobrevivam até a idade adulta.[16] No caso dos ovos, isso significa que estrategistas r põem grande quantidade de ovos, enquanto estrategistas K tomam cuidado para proteger um número menor de ovos. Percevejos do gênero Corythucha estão sujeitos à predação de ovos por predadores obrigatórios, como mirídeos, percevejos e tripes, e respondem de maneiras variadas. As fêmeas de C. solani defendem seus ovos dos predadores, enquanto C. marmorata enterra seus ovos dentro de folhas e os distribui no espaço e no tempo.[2]

Os ninhos de aves são vulneráveis à predação de ovos, especialmente para espécies como os êiders, que nidificam no solo. Em resposta ao roubo de ovos de seus ninhos, metade dos indivíduos começa uma nova ninhada em um novo ninho, sempre evitando a área ao redor do ninho roubado.[4] Aves que nidificam em árvores também sofrem predação por serpentes, mamíferos e aves, particularmente em florestas tropicais. Na Costa Rica, a taxa de predação de ninhos artificiais foi maior em altitudes intermediárias (entre 500 e 650 m), com uma diminuição em altitudes mais elevadas, até 2.740 m. Isso pode explicar por que muitas espécies de aves migram para altitudes mais altas para se reproduzir.[17] A predação de ovos por serpentes raramente é diretamente confrontada, mas o lagarto Eutropis longicaudata protege agressivamente seus ovos da cobra Oligodon formosanus.[15][18][19]
Os ovos das aves são coloridos e padronizados, aparentemente para camuflagem, enganando os olhos dos predadores. Por exemplo, os maçaricos-reais nidificam entre gramíneas altas e possuem ovos verdes e manchados, semelhantes ao ambiente, além de serem defendidos pelos adultos. Em contraste, os ovos dos borrelhos-pequenos-de-coleira, depositados em praias de seixos, são pálidos e salpicados, difíceis de distinguir entre as pedras.[20]
Registro fóssil
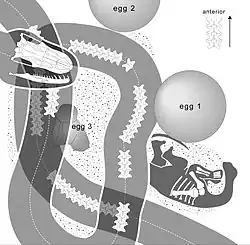
A predação de ovos pode ser uma estratégia de alimentação antiga. Um fóssil da serpente do Cretáceo Superior Sanajeh, do oeste da Índia, encontrado enrolado em torno de um ovo e um filhote de dinossauro saurópode, provavelmente era um predador de ninhos de saurópodes, incluindo ovos. A Sanajeh tinha cerca de 3,5 m de comprimento; seu crânio media 95 mm.[21] O Oviraptor era um dinossauro do Cretáceo Superior; seu nome, que significa "ladrão de ovos", foi dado inicialmente por se acreditar que era um predador de ovos. Mais tarde, descobriu-se que ele chocava seus próprios ovos, e suas mandíbulas sem dentes foram reinterpretadas como adaptadas a uma dieta diferente, possivelmente de folhas.[22][23][24][25]
Dinossauros saurópodes, alguns dos maiores animais que já existiram, parecem, surpreendentemente, ter seguido uma estratégia reprodutiva do tipo r, produzindo grande quantidade de ovos com casca dura. Isso contrasta com a estratégia K das baleias, mamíferos marinhos de tamanho comparável. As baleias produzem poucos ovos que se desenvolvem internamente, recebendo alto investimento parental. Uma possível causa é que o tamanho dos ovos é limitado: extrapolando a partir do tamanho dos ovos de aves em relação ao peso corporal do adulto, um saurópode de 10 toneladas produziria ovos pesando cerca de 333 kg, muito acima do limite (cerca de 10 kg) que uma casca de ovo poderia suportar. Se isso estiver correto, os saurópodes inevitavelmente tinham que seguir uma estratégia r com muitos ovos relativamente pequenos, não especificamente como resposta à predação de ovos.[26]
Interação com humanos
Danos à pesca comercial
Entre os peixes, a predação de ovos por espécies como o hadoque (Melanogrammus aeglefinus) pode contribuir para o declínio de populações de peixes comercialmente importantes, como o arenque do Atlântico (Clupea harengus). Esse efeito pode ser relevante em esforços para restaurar pescarias danificadas pela pesca predatória.[27]
Espécies introduzidas e invasoras
_(cropped).jpg)
Espécies invasoras frequentemente predam ovos e filhotes de espécies nativas. A joaninha-asiática Harmonia axyridis [en] consome ovos de outras espécies, incluindo outras joaninhas, como a Adalia bipunctata. As fêmeas da espécie predada põem ovos com maior quantidade de alcaloides defensivos quando ocorre a predação de ovos.[28]
A predação de ovos é uma ameaça especialmente grave para colônias de aves marinhas que nidificam no solo. Essas aves frequentemente escolhem ilhas afastadas como locais de nidificação, pois historicamente essas ilhas tinham menos predadores que o continente. Introduções acidentais de espécies predadoras perturbaram a reprodução de aves marinhas, já que os predadores encontram uma oferta concentrada de alimento na forma de ovos no solo ou em tocas, podendo se multiplicar rapidamente. Populações de ilhas na Australásia foram amplamente afetadas por espécies exóticas, como ratos, trazidos por navios da Eurásia. Espécies nativas, como lagartos Tiliqua nigrolutea [en] e ratos Hydromys chrysogaster, também podem impactar aves marinhas, como a pardela-de-cauda-curta (Ardenna tenuirostris), em ilhas da Tasmânia, embora as taxas de predação sejam relativamente baixas. Os ovos geralmente são tomados quando as tocas estão desprotegidas, sugerindo que os pais conseguem defender seus ovos eficazmente contra esses predadores.[29]
Controle de predadores de ovos
Onde populações de interesse estão ameaçadas por predadores de ovos, os conservacionistas podem tentar controlar os predadores para permitir a recuperação da espécie presa. No caso de predadores aviários, uma abordagem é utilizar ovos-isca tratados com o avicida de ação lenta DRC-1339. Isso, por exemplo, controlou corvos que ameaçavam o tetraz que nidifica no solo,[30] e, entre aves marinhas, gaivotas que ameaçavam colônias de esternídeos.[31] Tartarugas marinhas se reproduzem depositando e enterrando seus ovos em praias de nidificação, de modo que o controle de predadores de ovos nesses locais pode ser eficaz para auxiliar a recuperação das populações de tartarugas.[32]
Referências
- ↑ «ovivorous (adj.)». Online Etymology Dictionary. Consultado em 26 de maio de 2025
- ↑ a b Tallamy, Douglas W.; Denno, Robert F. (1981). «Alternative Life History Patterns in Risky Environments: An Example from Lacebugs». Proceedings in Life Sciences. [S.l.: s.n.] pp. 129–147. ISBN 978-1-4612-5943-5. doi:10.1007/978-1-4612-5941-1_9
- ↑ «egg parasite». Merriam-Webster. Consultado em 26 de maio de 2025
- ↑ a b Hanssen, Sveinn Are; Erikstad, Kjell Einar (2012). «The long-term consequences of egg predation». Behavioral Ecology. 24 (2): 564–569. doi:10.1093/beheco/ars198

- ↑ Goodwin, D. (1983). Crows of the World. [S.l.]: Queensland University Press. p. 92. ISBN 0-7022-1015-3
- ↑ Ainsworth, Gill; Calladine, John; Martay, Blaise; Park, Kirsty; Redpath, Steve; Wernham, Chris; Wilson, Mark; Young, Juliette (2017). Understanding Predation: A review bringing together natural science and local knowledge of recent wild bird population changes and their drivers in Scotland. [S.l.]: Scotland's Moorland Forum. pp. 233–234
- ↑ Ekanayake, Kasun B.; Sutherland, Duncan R.; Dann, Peter; Weston, Michael A. (2015). «Out of sight but not out of mind: corvids prey extensively on eggs of burrow-nesting penguins». Wildlife Research. 42 (6): 509. doi:10.1071/wr15108
- ↑ Van Lawick-Goodall, Jane (1971). Tool-using in primates and other vertebrates. Col: Advances in the study of behavior. 3. [S.l.]: Academic Press. pp. 195–249. ISBN 9780120045037. doi:10.1016/S0065-3454(08)60157-6
- ↑ a b Pike, David A.; Clark, Rulon W.; Manica, Andrea; et al. (2016). «Surf and turf: predation by egg-eating snakes has led to the evolution of parental care in a terrestrial lizard». Scientific Reports. 6 (1): 22207. Bibcode:2016NatSR...622207P. PMC 4768160. PMID 26915464. doi:10.1038/srep22207
- ↑ a b c Li, Min; Fry, B.G.; Kini, R. Manjunatha (2005). «Eggs-Only Diet: Its Implications for the Toxin Profile Changes and Ecology of the Marbled Sea Snake (Aipysurus eydouxii)». Journal of Molecular Evolution. 60 (1): 81–89. Bibcode:2005JMolE..60...81L. PMID 15696370. doi:10.1007/s00239-004-0138-0
- ↑ a b c Gans, Carl (1952). «The functional morphology of the egg-eating adaptations in the snake genus Dasypeltis». Zoologica. 37 (18): 209–244
- ↑ a b Dandge, P. H.; Tiple, A. D. (2016). «Notes on Natural History, New Distribution Records and Threats of Indian Egg Eater Snake Elachistodon westermanni Reinhardt, 1863 (Serpentes: Colubridae): Implications for Conservation». Russian Journal of Herpetology. 23 (1): 55-62. doi:10.30906/1026-2296-2016-23-1-55-62
- ↑ Light, Jessica E.; Fiumera, Anthony C.; Porter, Brady A. (2005). «Egg-feeding in the freshwater piscicolid leech Cystobranchus virginicus (Annelida, Hirudinea)». Invertebrate Biology. 124 (1): 50–56. doi:10.1111/j.1744-7410.2005.1241-06.x
- ↑ Cavalleri, Adriano; de Souza, André R.; Prezoto, Fábio; Mound, Laurence A. (2013). «Egg predation within the nests of social wasps: a new genus and species of Phlaeothripidae, and evolutionary consequences of Thysanoptera invasive behaviour». Biological Journal of the Linnean Society. 109 (2): 332–341. doi:10.1111/bij.12057
- ↑ a b «Snakes meet their match in offspring-protecting lizards». Phys.org. 2013. Consultado em 26 de maio de 2025
- ↑ Verhulst, P.F. (1838). «Notice sur la loi que la population pursuit dans son accroissement». Corresp. Math. Phys. 10: 113–121
- ↑ Boyle, W. Alice (2008). «Can variation in risk of nest predation explain altitudinal migration in tropical birds?». Oecologia. 155 (2): 397–403. Bibcode:2008Oecol.155..397B. PMID 18188606. doi:10.1007/s00442-007-0897-6
- ↑ Huang, W.-S.; Pike, D. A. (2011). «Does maternal care evolve through egg recognition or directed territoriality?». Journal of Evolutionary Biology. 24 (9): 1984–1991. PMID 21649766. doi:10.1111/j.1420-9101.2011.02332.x
- ↑ Huang, Wen-San; Lin, Si-Min; Dubey, Sylvain; Pike, David A. (2012). Coulson, Tim, ed. «Predation drives interpopulation differences in parental care expression». Journal of Animal Ecology. 82 (2): 429–437. PMID 23237108. doi:10.1111/1365-2656.12015
- ↑ Stoddard, Mary Caswell; Marshall, Kate L.A.; Kilner, Rebecca M. (2011). «Imperfectly Camouflaged Avian Eggs: Artefact or Adaptation?». Avian Biology Research. 4 (4): 196–213. doi:10.3184/175815511x13207484398647
- ↑ Wilson, J.A.; Mohabey, D.M.; Peters, S.E.; Head, J.J. (2010). Benton, Michael J., ed. «Predation upon Hatchling Dinosaurs by a New Snake from the Late Cretaceous of India». PLOS Biology. 8 (3): e1000322. PMC 2830453. PMID 20209142. doi:10.1371/journal.pbio.1000322
- ↑ Norell, M. A.; Clark, J. M.; Dashzeveg, D.; Barsbold, R.; Chiappe, L. M.; Davidson, A. R.; McKenna, M. C.; Altangerel, P.; Novacek, M. J. (1994). «A theropod dinosaur embryo and the affinities of the Flaming Cliffs Dinosaur eggs». Science. 266 (5186): 779−782. Bibcode:1994Sci...266..779N. JSTOR 2885545. PMID 17730398. doi:10.1126/science.266.5186.779
- ↑ Norell, M. A.; Clark, J. M.; Chiappe, L. M.; Dashzeveg, D. (1995). «A nesting dinosaur». Nature. 378 (6559): 774−776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0
- ↑ Dong, Z.; Currie, P. J. (1996). «On the discovery of an oviraptorid skeleton on a nest of eggs at Bayan Mandahu, Inner Mongolia, People's Republic of China». Canadian Journal of Earth Sciences. 33 (4): 631−636. Bibcode:1996CaJES..33..631D. doi:10.1139/e96-046
- ↑ Smith, D. K. (1990). «Osteology of Oviraptor philoceratops, a possible herbivorous theropod from the Upper Cretaceous of Mongolia». Journal of Vertebrate Paleontology. 10 (supp. 003): 42A. doi:10.1080/02724634.1990.10011841
- ↑ Sander, P. Martin; Peitz, Christian; Jackson, Frankie D.; Chiappe, Luis M. (2008). «Upper Cretaceous titanosaur nesting sites and their implications for sauropod dinosaur reproductive biology». Palaeontographica, Abteilung A. 284 (4–6): 69–107. Bibcode:2008PalAA.284...69S. doi:10.1127/pala/284/2008/69
- ↑ Richardson, D. E.; Hare, J. A.; Fogarty, M. J.; Link, J. S. (2011). «Role of egg predation by haddock in the decline of an Atlantic herring population». Proceedings of the National Academy of Sciences. 108 (33): 13606–13611. Bibcode:2011PNAS..10813606R. PMC 3158212. PMID 21825166. doi:10.1073/pnas.1015400108
- ↑ Paul, Sarah C.; Stevens, Martin; Burton, Jake; et al. (2018). «Invasive Egg Predators and Food Availability Interactively Affect Maternal Investment in Egg Chemical Defense». Frontiers in Ecology and Evolution. 6. doi:10.3389/fevo.2018.00004. hdl:10871/30982
- ↑ Carey, Mark J. (2010). «Predation of short-tailed shearwater eggs on Great Dog Island, Tasmania». Australian Field Ornithology. 27 (2): 59–64
- ↑ Spencer, Jack O. (2002). «DRC-1339 use and control of common ravens». Proceedings of the Vertebrate Pest Conference. 20. doi:10.5070/v420110074
- ↑ Blodget, Bradford G.; Henze, Laura (1991). «Use of DRC-1339 to eliminate gulls and re-establish a tern nesting colony in Buzzards Bay, Massachusetts». Fifth Eastern Wildlife Damage Control Conference. article 5
- ↑ McCullough, Dale (1992). Wildlife 2001 : populations. Dordrecht: Springer. p. 352. ISBN 978-94-011-2868-1. OCLC 840309647