Teoria da seleção r/K
A teoria da seleção r/K, modelo evolutivo r/K ou estratégia de reprodução r/K refere-se a um modelo de seleção de características biológicas que promovem o sucesso em determinados ambientes. Esta teoria teve origem nos estudos de biogeografia insular dos ecólogos Robert MacArthur e Edward Osborne Wilson.[1]
Na teoria da seleção r/K, põe-se a hipótese de as pressões seletivas orientarem a evolução para duas direções estereotipadas: à seleção r ou à seleção K.[2]
Contexto histórico e definição
%252C_Anaxyrus_americanus_ssp._americanus%252C_Fairfax_VA_(22833803).jpg)
O desenvolvimento da Teoria da Seleção r/K ocorreu no contexto de expansão dos modelos matemáticos em ecologia de populações durante o pós-guerra, especialmente a partir da década de 1960. Esse ambiente acadêmico, favorecido pelo avanço tecnológico e computacional, possibilitou a formulação de teorias ecológicas mais quantitativas.[1]
_female_with_six-week-old_baby.jpg)
Nesse cenário, Robert MacArthur e Edward O. Wilson desenvolveram uma teoria que buscava explicar a dinâmica de populações e as estratégias de história de vida das espécies, particularmente no contexto da ocupação de ilhas e ambientes insulares. Os autores propuseram que, em ambientes novos ou instáveis — como ilhas recém-colonizadas, onde os recursos são inicialmente abundantes e a competição é reduzida — a seleção natural tenderia a favorecer espécies com alta taxa de crescimento populacional, rápida maturação e grande número de descendentes: os chamados estrategistas-r. Por outro lado, em ambientes mais previsíveis e estáveis, onde as populações se aproximam da capacidade de suporte (K) do ambiente, a seleção tenderia a favorecer espécies mais competitivas, com menor número de descendentes, maior investimento parental e estratégias de reprodução mais lentas: os estrategistas-K.[1][2]
Durante as décadas de 1970 e 1980, autores como Pianka e Parry expandiram e refinaram esses conceitos, relacionando-os a aspectos de história de vida, como tamanho da prole, idade de maturação e investimento parental. Assim, definiu-se que espécies associadas à seleção-r tendem a apresentar rápida maturação, alta fecundidade e baixo cuidado parental, enquanto espécies sob seleção-K investem mais em cada descendente, apresentam ciclos de vida mais longos e menor taxa de fecundidade.[2][3]
Atualmente, reconhece-se que a dicotomia r/K funciona como um modelo conceitual útil para alguns contextos, mas simplificado frente à complexidade ecológica real. Muitos estudos contemporâneos demonstram que as espécies podem apresentar características mistas ou contextualmente variáveis, e a compreensão das estratégias de história de vida passou a se basear em abordagens multivariadas e contínuas, considerando os trade-offs entre crescimento, reprodução e sobrevivência em diferentes ambientes.[1][3]
Contexto e aplicação matemática

A teoria de seleção r e K, proposta no século XX, fundamenta-se em modelos matemáticos de dinâmica populacional, que utilizam equações diferenciais.[4] Em 1798, Thomas Malthus propôs que a população humana tende a crescer em progressão geométrica, em um modelo que não considera os fatores limitantes do ambiente [5]. Posteriormente, o matemático belga Pierre-François Verhulst propôs em 1838 uma equação para modelar o crescimento populacional de forma mais realista do que o modelo de crescimento exponencial de Malthus (1798), incorporando a noção de capacidade de suporte ambiental, limitada por recursos como alimento, espaço e predadores.[5][6] A equação logística proposta por Pierre-François Verhulst em 1838, tornou-se um marco para a compreensão de dinâmicas populacionais e serviu de base conceitual e matemática para o desenvolvimento, mais tarde, da teoria de seleção r/K.[2]
A equação abaixo descreve um crescimento populacional que é inicialmente exponencial, mas que desacelera à medida que a população se aproxima de K, estabilizando-se eventualmente. As variáveis da equação são r, a taxa intrínseca de crescimento; N, o número de indivíduos; e K, historicamente referido como o limite superior de crescimento ou capacidade suporte, fator que não era considerado nas equações de Malthus[7].

Entender as variáveis da equação é fundamental para entender a dinâmica das populações. Quando o número de indivíduos (N) é pequeno em relação à capacidade de suporte (K), a razão, fazendo com que o valor da expressão . Portanto, em populações muito pequenas, o número de indivíduos em um determinado instante de tempo t depende apenas da taxa de crescimento (r) e do número de indivíduos (N), caracterizando um crescimento exponencial. Esse tipo de padrão é observado em estrategistas r, que vivem em ambientes instáveis ou buscam a colonização[6].



Porém, quando valor de N é próximo de K, ou seja, quando o número de indivíduos se aproxima da capacidade suporte do meio, a razão , o que resulta no valor de , representando justamente a tendência a estabilização do crescimento através da pressão ambiental, podendo atuar através da competição, limitação de espaço, doenças, predação ou outras formas de pressão.[6]


Assim, a taxa de crescimento populacional é máxima quando o número de indivíduos (N) é pequeno e vai diminuindo à medida que N se aproxima da capacidade suporte (K) até se estabilizar, refletindo o equilíbrio ecológico. Portanto, a equação logística não apenas descreve o comportamento dinâmico das populações, mas também oferece a fundamentação matemática para a distinção entre estratégias adaptativas em ambientes com diferentes graus de estabilidade e disponibilidade de recursos, conceito central para a teoria de seleção r/K [6][8].

Aplicação ecológica
Teoria r/K no crescimento populacional
Crescimento populacional é a variação no número de indivíduos de uma população ao longo do tempo, que pode ser influenciado por fatores como natalidade, mortalidade, imigração e emigração [1]. Estudar como e por que as populações aumentam ou diminuem é fundamental na ecologia, pois permite entender melhor as relações ecológicas, prever mudanças em comunidades naturais e planejar a gestão de recursos e conservação de espécies [9][10].
Em face deste equilíbrio entre os investimentos de energia dedicados a processos de crescimento, sobrevivência e reprodução, que pode ser decisivo para o provimento e manutenção de geração seguintes. Nesse contexto, os autores MacArthur e Wilson demonstraram que as populações não podem crescer exponencialmente devido às limitações existentes no meio, como por exemplo a falta de recursos como água e comida [1]. Essas limitações permitem definir a capacidade de suporte (K) do meio, que corresponde ao número máximo de indivíduos de uma população que um ambiente pode sustentar de forma estável, considerando os recursos (alimento, água, energia) e as condições (fatores abióticos como luminosidade e temperatura) [7][11].
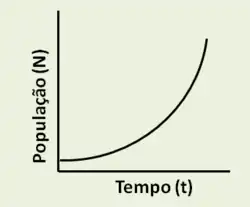
O crescimento populacional pode ser descrito por dois modelos principais: Crescimento logístico e crescimento exponencial [12]. No crescimento logístico, a população aumenta rapidamente no início, desacelerando progressivamente conforme se aproxima da capacidade de suporte, enquanto no crescimento exponencial o número de indivíduos cresce sem considerar essa limitação ambiental, podendo atingir níveis muito altos antes de sofrer um declínio brusco provocado por fatores limitantes [11][12]. Essas diferenças no tipo de crescimento permitem classificar as populações com base na sua estratégia de sobrevivência, sendo os estrategistas r, como já visto, as espécies que se reproduzem rapidamente, gerando grande número de descendentes com pouca ou nenhuma proteção parental e que tendem a colonizar ambientes instáveis ou imprevisíveis, onde a competição é baixa e as condições podem mudar rapidamente, podendo apresentar perfil de crescimento próximo ao exponencial [2]. Entretanto, os estrategistas K possuem menor taxa de reprodução, gerando poucos descendentes, mas com alto investimento parental, sendo espécies que vivem em ambientes mais estáveis, próximos da capacidade de suporte, onde a competição por recursos é intensa e o perfil de crescimento tende a ser do tipo logístico [1][2].
Teoria r/K e estratégias de sucessão ecológica
Seja na ocupação de um novo habitat ainda não colonizado - ilhas inabitadas - ou no restabelecimento da fauna e flora de uma área afetada por alguma perturbação, como incêndios, o processo de sucessão ecológica estará atuando [1]. A sucessão ecológica é justamente o processo de mudança gradual na composição e estrutura de comunidades ecológicas ao longo do tempo, após uma perturbação ou formação de um novo habitat [1][13].
Dentro desse contexto, a teoria r/K auxilia na compreensão das estratégias de colonização e permanência das espécies ao longo das diferentes fases da sucessão [1]. Espécies com altas taxas de reprodução e crescimento tendem a sobreviver mais facilmente nos estágios iniciais de colonização de ambientes, quando a densidade populacional é baixa, o que muda quando a densidade populacional aumenta nos estágios mais avançados da colonização, onde a pressão seletiva tende a favorecer espécies com menor potencial de crescimento e maiores especializações para sobrevivência competitiva na maior densidade [14]. Portanto, nos estágios iniciais, o ambiente é instável, com poucos recursos e alta disponibilidade de espaço, favorecendo espécies estrategistas r, caracterizadas por elevada taxa de reprodução, desenvolvimento rápido e ampla capacidade de dispersão, mas baixo investimento em cuidados parentais e menor competitividade [1][2]. Essas espécies são pioneiras, dominam rapidamente os ambientes recém-abertos, mas tendem a ser substituídas conforme a densidade populacional aumenta e a competição se intensifica.[14] À medida que a sucessão progride e a comunidade ecológica se torna mais estruturada e estável, as condições ambientais mudam, aumentando a competição por recursos e a densidade populacional. Nesse cenário, passam a predominar espécies estrategistas K, que apresentam menor taxa de reprodução, maior longevidade e alto investimento parental, sendo mais eficientes em ambientes saturados e competitivos [1][2][15].
MacArthur & Wilson apontam que as seleções r e K são forças seletivas opostas que constituem extremos de um gradiente, ou seja, nenhum organismo seria completamente r-selecionado ou K-selecionado, mas se situaria estrategicamente sempre em um ambiente em particular, em um dado momento no tempo, e em relação a outro organismo comparativamente. Ainda assim, comparativamente, algumas características podem ser associadas a cada uma destas estratégias.[1]
Esse padrão, identificado tanto em sucessões vegetais quanto animais, reforça o papel da teoria r/K como ferramenta para explicar não apenas o crescimento populacional, mas também a dinâmica de sucessão de comunidades ecológicas.[14] Assim, entender as estratégias ecológicas permite prever quais espécies tendem a colonizar áreas perturbadas e quais serão as características das espécies que ocuparão o ambiente nas fases finais da sucessão, contribuindo para práticas de conservação e restauração ecológica.
Seleção r/K em contextos globais atuais
Mudanças climáticas
As estratégias r e K, como já mencionado, são moldadas pelas condições ambientais de um determinado ecossistema, sendo que espécies r-estrategistas tendem a predominar em ambientes instáveis, enquanto espécies k-estrategistas são mais comuns em ambientes estáveis. Assim, mudanças drásticas nos ecossistemas podem gerar desequilíbrios nas densidades populacionais, e a estratégia de vida torna-se um fator essencial para determinar a resistência das espécies em sobreviver e gerar descendentes frente às condições impostas pelo meio. Atualmente, as mudanças climáticas globais e seus impactos, diretos e indiretos, sobre a biodiversidade têm sido foco de inúmeros estudos. Diversas linhas de pesquisa indicam que esse fenômeno pode ser um dos principais fatores de extinção de espécies ao longo do próximo século [16]. O impacto das mudanças climáticas são visíveis em muitas espécies no planeta, e algumas são potencialmente mais vulneráveis do que outras. Espécies com menor taxa reprodutiva, característica de k-estrategistas, podem ser consideradas mais suscetíveis às mudanças climáticas [17]. Por outro lado, espécies com maior taxa de reprodução, típicas de r-estrategistas, tendem a ser menos impactadas, pois possuem maior capacidade de expandir sua área de ocorrência para regiões menos afetadas pelas alterações climáticas [18].

Invasões biológicas
A invasão biológica constitui uma das maiores ameaças à biodiversidade[19]. O sucesso da invasão por um ser vivo depende de dois fatores: as características da espécie não-nativa para invadir de forma eficiente um local e as propriedades de um ecossistema que o tornam suscetível à invasão[20]. O conhecimento da estratégia de vida de uma espécie é essencial para estimar suas chances de sobrevivência e estabelecimento em áreas fora de sua distribuição natural. Espécies de plantas r-estrategistas são mais prováveis de terem sucesso na invasão, visto que, normalmente as invasões ocorrem em ambientes com distúrbios naturais e/ou antrópicos.[21] Além disso, as características de reprodução de plantas com esse tipo de estratégia, como alta quantidade de produção de sementes, menor tamanho da semente e período juvenil mínimo baixo auxiliam no sucesso da invasão[20]. No entanto, também há espécies invasoras com estratégia K, que obtêm êxito por meio de alta capacidade competitiva em ambientes estáveis, muitas vezes superando espécies nativas[22]. Assim, o estudo das estratégias reprodutivas e de história de vida de espécies potencialmente invasoras é crucial para ações de prevenção e manejo, reduzindo riscos à biodiversidade nativa e à integridade dos ecossistemas.
Estratégia r/K em recursos econômicos vegetais
A castanha-do-pará (Bertholletia excelsa), também conhecida como castanha-do-brasil, é uma espécie arbórea da família Lecythidaceae, nativa da Amazônia, com ocorrência no Brasil, Bolívia e Peru. Suas sementes são altamente apreciadas, sendo tradicionalmente exploradas por povos locais há séculos[23][24] . A partir do século XX, essa exploração tornou-se mais intensa, com foco na comercialização e exportação. A coleta e venda das sementes de B. excelsa configuram-se como uma importante fonte de renda para comunidades extrativistas que vivem em regiões com alta densidade da espécie [25].

No contexto da exploração comercial de espécies vegetais e animais, torna-se essencial o conhecimento da estratégia de vida das espécies para embasar planos de manejo que previnam a super exploração e os consequentes impactos sobre as populações e os ecossistemas. A castanha-do-pará apresenta características típicas de espécies K-estrategistas, como maturação reprodutiva tardia, longa longevidade e baixa taxa reprodutiva, o que aumenta sua susceptibilidade à exploração humana [26]. Estudos demográficos indicam que a coleta intensiva de sementes reduz a presença de indivíduos jovens, comprometendo a regeneração natural e tornando indispensáveis o manejo sustentável e a implementação de políticas regulatórias [27][28].
Referências
- ↑ a b c d e f g h i j k l m MacArthur, R. e Edward Osborn Wilson (1967). The Theory of Island Biogeography, Princeton University Press (2001 reprint), ISBN 0-691-08836-5M.
- ↑ a b c d e f g h Pianka, E. R. (1970). On r and K selection. American Naturalist 104, 592-597.
- ↑ a b Parry, Gregory D. (1 de março de 1981). «The meanings of r- and K-selection». Oecologia (em inglês) (2): 260–264. ISSN 1432-1939. doi:10.1007/BF00347974. Consultado em 21 de junho de 2025
- ↑ Getz, Wayne M. (1 de maio de 1993). «Metaphysiological and evolutionary dynamics of populations exploiting constant and interactive resources:R—K selection revisited». Evolutionary Ecology (em inglês) (3): 287–305. ISSN 1573-8477. doi:10.1007/BF01237746. Consultado em 14 de junho de 2025
- ↑ a b Sojecka, Agata Angelika; Drozd-Rzoska, Aleksandra (18 de junho de 2024), Verhulst Equation and the Universal Pattern for the Global Population Growth, doi:10.48550/arXiv.2406.13016, consultado em 14 de junho de 2025
- ↑ a b c d Bacaër, Nicolas (2011). Bacaër, Nicolas, ed. «Verhulst and the logistic equation (1838)». London: Springer (em inglês): 35–39. ISBN 978-0-85729-115-8. doi:10.1007/978-0-85729-115-8_6. Consultado em 14 de junho de 2025
- ↑ a b Chapman, Eric J.; Byron, Carrie J. (1 de janeiro de 2018). «The flexible application of carrying capacity in ecology». Global Ecology and Conservation: e00365. ISSN 2351-9894. doi:10.1016/j.gecco.2017.e00365. Consultado em 14 de junho de 2025
- ↑ Boyce, Mark S. (1984). «Restitution of r- and K-Selection as a Model of Density-Dependent Natural Selection». Annual Review of Ecology and Systematics: 427–447. ISSN 0066-4162. Consultado em 21 de junho de 2025
- ↑ Nichols, James D.; Breininger, David R.; Armstrong, Doug P.; Lacy, Robert C. (28 de outubro de 2024). «Habitat management decisions for conservation: a conceptual framework». Frontiers in Conservation Science (em inglês). ISSN 2673-611X. doi:10.3389/fcosc.2024.1450000. Consultado em 14 de junho de 2025
- ↑ Lande, Russell; Engen, Steinar; Sæther, Bernt-Erik (2003). Stochastic Population Dynamics in Ecology and Conservation (em inglês). [S.l.]: Oxford University Press. Consultado em 14 de junho de 2025
- ↑ a b Bradshaw, Corey J. A.; Herrando-Pérez, Salvador (2023). «Logistic-growth models measuring density feedback are sensitive to population declines, but not fluctuating carrying capacity». Ecology and Evolution (em inglês) (4): e10010. ISSN 2045-7758. PMC 10131297
 . PMID 37122772. doi:10.1002/ece3.10010. Consultado em 14 de junho de 2025
. PMID 37122772. doi:10.1002/ece3.10010. Consultado em 14 de junho de 2025
- ↑ a b DEWAN, Nalin. Population Modelling–Exponential and Logistic Models. 2021.
- ↑ Connell, Joseph H.; Slatyer, Ralph O. (novembro de 1977). «Mechanisms of Succession in Natural Communities and Their Role in Community Stability and Organization». The American Naturalist (982): 1119–1144. ISSN 0003-0147. doi:10.1086/283241. Consultado em 14 de junho de 2025
- ↑ a b c Reznick, David; Bryant, Michael J.; Bashey, Farrah (2002). «r- AND K-SELECTION REVISITED: THE ROLE OF POPULATION REGULATION IN LIFE-HISTORY EVOLUTION». Ecology (em inglês) (6): 1509–1520. ISSN 1939-9170. doi:10.1890/0012-9658(2002)083[1509:RAKSRT]2.0.CO;2. Consultado em 14 de junho de 2025
- ↑ Grime, J. P. (novembro de 1977). «Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory». The American Naturalist (982): 1169–1194. ISSN 0003-0147. doi:10.1086/283244. Consultado em 14 de junho de 2025
- ↑ Pacifici, Michela; Foden, Wendy B.; Visconti, Piero; Watson, James E. M.; Butchart, Stuart H. M.; Kovacs, Kit M.; Scheffers, Brett R.; Hole, David G.; Martin, Tara G. (março de 2015). «Assessing species vulnerability to climate change». Nature Climate Change (em inglês) (3): 215–224. ISSN 1758-6798. doi:10.1038/nclimate2448. Consultado em 22 de junho de 2025
- ↑ Pacifici, Michela; Visconti, Piero; Butchart, Stuart H. M.; Watson, James E. M.; Cassola, Francesca M.; Rondinini, Carlo (março de 2017). «Species' traits influenced their response to recent climate change». Nature Climate Change (em inglês) (3): 205–208. ISSN 1758-6798. doi:10.1038/nclimate3223. Consultado em 22 de junho de 2025
- ↑ Angert, Amy L.; Crozier, Lisa G.; Rissler, Leslie J.; Gilman, Sarah E.; Tewksbury, Josh J.; Chunco, Amanda J. (2011). «Do species' traits predict recent shifts at expanding range edges?». Ecology Letters (em inglês) (7): 677–689. ISSN 1461-0248. doi:10.1111/j.1461-0248.2011.01620.x. Consultado em 22 de junho de 2025
- ↑ Simberloff, Daniel; Martin, Jean-Louis; Genovesi, Piero; Maris, Virginie; Wardle, David A.; Aronson, James; Courchamp, Franck; Galil, Bella; García-Berthou, Emili (1 de janeiro de 2013). «Impacts of biological invasions: what's what and the way forward». Trends in Ecology & Evolution (em inglês) (1): 58–66. ISSN 0169-5347. PMID 22889499. doi:10.1016/j.tree.2012.07.013. Consultado em 22 de junho de 2025
- ↑ a b Closset-Kopp, Déborah; Chabrerie, Olivier; Valentin, Bertille; Delachapelle, Hermine; Decocq, Guillaume (15 de agosto de 2007). «When Oskar meets Alice: Does a lack of trade-off in r/K-strategies make Prunus serotina a successful invader of European forests?». Forest Ecology and Management (1): 120–130. ISSN 0378-1127. doi:10.1016/j.foreco.2007.04.023. Consultado em 22 de junho de 2025
- ↑ Rejmánek, Marcel; Richardson, David M. (1996). «What Attributes Make Some Plant Species More Invasive?». Ecology (6): 1655–1661. ISSN 0012-9658. doi:10.2307/2265768. Consultado em 22 de junho de 2025
- ↑ Duyck, Pierre-François; David, Patrice; Quilici, Serge (2007). «Can more K-selected species be better invaders? A case study of fruit flies in La Réunion». Diversity and Distributions (em inglês) (5): 535–543. ISSN 1472-4642. doi:10.1111/j.1472-4642.2007.00360.x. Consultado em 22 de junho de 2025
- ↑ Mori, Scott A.; Prance, Ghillean T. (1990). «Taxonomy, Ecology, and Economic Botany of the Brazil Nut (Bertholletia excelsa Humb. & Bonpl.: Lecythidaceae)». Advances in Economic Botany: 130–150. ISSN 0741-8280. Consultado em 22 de junho de 2025
- ↑ Wadt, Lúcia H. O.; Kainer, Karen A.; Gomes-Silva, Daisy A. P. (20 de junho de 2005). «Population structure and nut yield of a Bertholletia excelsa stand in Southwestern Amazonia». Forest Ecology and Management (3): 371–384. ISSN 0378-1127. doi:10.1016/j.foreco.2005.02.061. Consultado em 22 de junho de 2025
- ↑ Zuidema, Pieter (2003). Ecology and management of the Brazil nut tree (Bertholletia excelsa) (PDF). [S.l.]: PROMAB. 112 páginas. ISBN 90-393-3390-4
- ↑ Brook, Barry W.; Sodhi, Navjot S.; Bradshaw, Corey J. A. (1 de agosto de 2008). «Synergies among extinction drivers under global change». Trends in Ecology & Evolution (em inglês) (8): 453–460. ISSN 0169-5347. PMID 18582986. doi:10.1016/j.tree.2008.03.011. Consultado em 22 de junho de 2025
- ↑ Wadt, L. H. O.; Kainer, K. A.; Staudhammer, C. L.; Serrano, R. O. P. (1 de janeiro de 2008). «Sustainable forest use in Brazilian extractive reserves: Natural regeneration of Brazil nut in exploited populations». Biological Conservation (1): 332–346. ISSN 0006-3207. doi:10.1016/j.biocon.2007.10.007. Consultado em 22 de junho de 2025
- ↑ Peres, Carlos A.; Baider, Claudia; Zuidema, Pieter A.; Wadt, Lúcia H. O.; Kainer, Karen A.; Gomes-Silva, Daisy A. P.; Salomão, Rafael P.; Simões, Luciana L.; Franciosi, Eduardo R. N. (19 de dezembro de 2003). «Demographic Threats to the Sustainability of Brazil Nut Exploitation». Science (5653): 2112–2114. ISSN 0036-8075. doi:10.1126/science.1091698. Consultado em 22 de junho de 2025