Papa-mel-barulhento
Papa-mel-barulhento
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Subespécie leachi | |||||||||||||||||
| Estado de conservação | |||||||||||||||||
 Pouco preocupante (IUCN 3.1) [1] | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Nome binomial | |||||||||||||||||
| Manorina melanocephala (Latham, 1801) | |||||||||||||||||
| Distribuição geográfica | |||||||||||||||||
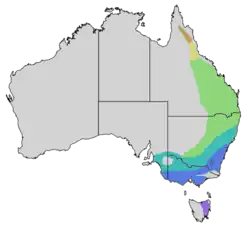 Área de distribuição do papa-mel-barulhento
ssp. titaniota ssp. lepidota ssp. melanocephala ssp. leachi zona intermediária zona intermediária | |||||||||||||||||
| Sinónimos | |||||||||||||||||
| |||||||||||||||||
O papa-mel-barulhento (Manorina melanocephala) é um pássaro da família Meliphagidae, e é endêmico do leste e sudeste da Austrália.[1] É um pássaro cinza, com cabeça preta, bico e pés amarelo-alaranjados, uma mancha amarela distinta atrás do olho e pontas brancas nas penas da cauda. A subespécie da Tasmânia tem detalhes amarelos mais intensos na asa e uma ponta branca mais larga na cauda. Os machos, as fêmeas e os filhotes têm aparência semelhante, embora as aves jovens sejam cinza-amarronzadas. Como o nome comum sugere, o papa-mel-barulhento é uma espécie vocal com uma grande variedade de cantos, chamadas, repreensões e alarmes, além de vocalizações quase constantes, principalmente das aves jovens. Uma das quatro espécies do gênero Manorina, o papa-mel-barulhento é dividido em quatro subespécies. A separação do M. m. leachi da Tasmânia é antiga, e as aves do continente foram divididas em 1999.
Encontrado em um amplo arco desde o extremo norte de Queensland, passando por Nova Gales do Sul e Vitória, até a Tasmânia e o sudeste da Austrália Meridional, o papa-mel-barulhento habita principalmente florestas secas e abertas de eucalipto que não possuem arbustos de sub-bosque. Isso inclui florestas dominadas por Corymbia maculata, bem como em bosques degradados em que o sub-bosque foi desmatado, como áreas queimadas recentemente, áreas agrícolas e de pastagem, reservas à beira de estradas e parques e jardins suburbanos com árvores e grama, mas sem arbustos densos. A densidade das populações de papa-mel-barulhento aumentou significativamente em muitos locais de sua área de distribuição, principalmente em habitats dominados por humanos. Acreditava-se que a popularidade das plantas de jardim produtoras de néctar, como as plantas do gênero Grevillea, desempenhava um papel em sua proliferação, mas estudos agora mostram que o papa-mel-barulhento se beneficiou principalmente das práticas de paisagismo que criam áreas abertas dominadas por eucaliptos.
O papa-mel-barulhento é gregário e territorial; eles se alimentam, tomam banho, se empoleiram, se reproduzem e defendem o território de forma comunitária, formando colônias que podem conter várias centenas de aves. Cada ave tem um “espaço de atividade”, e as aves com espaços de atividade sobrepostos formam associações, que são as unidades mais estáveis dentro da colônia. As aves também formam bandos temporários chamados de “coalizões” para atividades específicas, como atacar um predador (mobbing). A coesão do grupo é facilitada não apenas por vocalizações, mas também por exibições ritualizadas, que foram categorizadas como exibições de voo, exibições posturais e exibições faciais. O papa-mel-barulhento é um pássaro notavelmente agressivo, de modo que perseguições, bicadas, brigas, repreensões e ataques ocorrem durante todo o dia, tendo como alvo tanto os intrusos quanto os membros da colônia.
Forrageando na copa das árvores, em troncos e galhos e no solo, o papa-mel-barulhento se alimenta principalmente de néctar, frutas e insetos. Passa a maior parte do tempo colhendo a folhagem dos eucaliptos e pode satisfazer a maior parte de suas necessidades nutricionais com seiva e melada colhidas da folhagem. O papa-mel-barulhento não usa uma exibição estereotipada de cortejo sexual, mas a cópula é um evento comunitário frenético. Ele se reproduz durante todo o ano, construindo um ninho profundo em forma de xícara e colocando de dois a quatro ovos. A incubação é feita apenas pela fêmea, embora até vinte machos ajudantes cuidem dos recém-nascidos e dos filhotes. Eles têm uma série de estratégias para aumentar seu sucesso reprodutivo, incluindo várias ninhadas e o ataque de grupos de predadores. O aumento da população do mineiro barulhento foi correlacionado com a redução da diversidade aviária em paisagens afetadas pelo homem. Sua territorialidade significa que é improvável que a translocação seja uma solução para sua superabundância, e o abate foi proposto, embora o papa-mel-barulhento seja atualmente uma espécie protegida em toda a Austrália.
Taxonomia
O ornitólogo inglês John Latham descreveu o papa-mel-barulhento quatro vezes em sua obra de 1801, Supplementum Indicis Ornithologici, sive Systematis Ornithologiae, aparentemente sem saber que se tratava da mesma ave em cada caso: o Merops garrulus, o Gracula melanocephala, o Merops cucullatus e o Merops albifrons.[2][3] As primeiras anotações registraram sua tendência de assustar as presas quando os caçadores estavam prestes a atirar.[4] Foi como o Merops garrulus que ele foi pintado entre 1792 e 1797 por Thomas Watling.[5] John Gould tratou o nome Merops garrulus como a descrição original e o renomeou como Myzantha garrula em sua obra de 1865, Handbook to the Birds of Australia [en]. Ele observou que os colonos da Tasmânia o chamavam de "mineiro", e os aborígenes australianos o chamavam de cobaygin.[6] Que que gang era um nome aborígene local das Montanhas Azuis.[7]
No início do século XIX, os ornitólogos australianos começaram a usar o nome Manorina melanocephala, porque ela foi listada primeiro por Latham em 1801.[8] Esse uso não seguiu a letra do Código Internacional de Nomenclatura Zoológica e, em 2009, a Comissão Internacional de Nomenclatura Zoológica conservou o nome atual, suprimindo formalmente o nome M. garrula.[9] O nome da espécie melanocephala é derivado das palavras do grego antigo melas, “preto”, e kephale, “cabeça”,[10] referindo-se à seu píleo preto. Quatro subespécies são reconhecidas, incluindo a subespécie leachi encontrada no leste da Tasmânia. A população do continente foi dividida em três subespécies em 1999 por Richard Schodde: titaniota da Península do Cabo York em Queensland até o sul de Mareeba [en], lepidota da região central de Queensland e do interior de Nova Gales do Sul a oeste de Nyngan [en], e a subespécie nomeada melanocephala do sudeste de Nova Gales do Sul, Vitória e sul da Austrália Meridional. Há zonas amplas onde as aves são intermediárias entre as subespécies.[11] É necessário um estudo mais aprofundado para definir o status taxonômico dessas populações.[12]
O papa-mel-barulhento é uma das quatro espécies do gênero Manorina na grande família de melifagídeos conhecida como Meliphagidae. As outras três espécies do gênero Manorina são o papa-mel-de-orelha-preta (M. melanotis), o papa-mel-de-pescoço-amarelo (M. flavigula) e o papa-mel-sininho (M. melanophrys). Uma das características mais óbvias do gênero é uma mancha de pele amarela nua atrás dos olhos, o que lhes dá uma aparência estranha de “vesgos”. Dentro do gênero, o papa-mel-barulhento, o de orelha-preta e o de pescoço-amarelo formam o subgênero Myzantha.[11] O papa-mel-barulhento ocasionalmente hibridiza com o papa-mel-de-pescoço-amarelo.[12] A análise molecular mostrou que os melifagídeos estão relacionados com os Pardalotidae, Acanthizidae e Maluridae na grande superfamília Meliphagoidea.[13]
Descrição
Aparência

O papa-mel-barulhento é um melifagídeo grande, com 24 a 28 centímetros de comprimento, envergadura de 36 a 45 centímetros e peso de 70 a 80 gramas. Os pássaros machos, fêmeas e filhotes têm plumagem semelhante: cinza no dorso, na cauda e no peito, e branco por baixo, com listras brancas na nuca e no pescoço posterior, e no peito; testa e loro esbranquiçados; uma faixa preta sobre o píleo, bico amarelo-alaranjado brilhante e uma mancha distinta de pele amarela atrás do olho; uma ponta branca proeminente na cauda; um detalhe estreito amarelo-oliva na asa dobrada; e pernas e pés amarelo-alaranjados. Um filhote pode ser distinguido pela plumagem mais macia, uma coloração marrom no preto da cabeça e no cinza do dorso, e uma mancha de pele amarela-acinzentada mais opaca atrás do olho.[12]
O papa-mel-barulhento tem aparência semelhante ao papa-mel-de-pescoço-amarelo e ao papa-mel-de-orelha-preta; ele tem uma testa branca opaca e um píleo preto, enquanto os outros têm cabeças cinzas.[14]
Variações geográficas

A variação de tamanho do papa-mel-barulhento em sua área de distribuição segue a regra de Bergmann, ou seja, as aves tendem a ser maiores onde o clima é mais frio.[11] Os adultos do centro-leste e do norte de Queensland tendem a ter pouca ou nenhuma borda amarelo-oliva nas penas do dorso e das asas, e têm uma franja branca mais larga nas penas do pescoço posterior e do dorso, dando às aves de Queensland a aparência de ter listras mais distintas do que outras populações.[15] O comprimento das asas geralmente aumenta com a latitude, mas o M. m. leachi tem asas mensuravelmente mais curtas do que a raça indicada, embora não tenha sido encontrada nenhuma diferença significativa no comprimento das asas em um estudo que comparou populações ao norte de 30° S e ao sul do rio Murray.[15] A subespécie leachi também tem uma listra mais fina no pescoço posterior do que a raça indicada, uma coloração amarela mais intensa nas asas e uma ponta esbranquiçada ligeiramente mais larga na cauda.[12]
A subespécie titaniota, do extremo norte de Queensland, tem cauda mais curta, píleo mais pálido, mancha de pele amarela maior e partes superiores mais pálidas, sem o amarelo-oliva da raça indicada; e a lepidota, encontrada no oeste de Nova Gales do Sul, é menor do que a raça indicada, com píleo preto e partes superiores mais escuras e mosqueadas.[11]
Vocalizações
Como o nome comum sugere, o papa-mel-barulhento é uma espécie excepcionalmente vocal. Ele tem um repertório grande e variado de canções, chamados, repreensões e alarmes.[16] A maioria é alta e penetrante e consiste em notas únicas e ásperas.[17] Ele tem dois chamados de alarme de frequência ampla que são usados ao atacar intrusos em seu território ou quando predadores (inclusive humanos) são avistados; e um chamado de alarme de frequência estreita que é usado principalmente quando predadores aéreos são vistos, como o falcão-marrom [en] (Falco berigora) ou outros pássaros voadores grandes, inclusive a pega-australiana (Gymnorhina tibicen) e o carrauongue-malhado (Strepera graculina).[18] O chamado de alarme do predador aéreo é uma série de notas de assobio agudas e arrastadas. Os chamados de alarme de frequência ampla são uma série de notas de “churr”, graves e ásperas, que ocorrem em níveis baixos e altos de intensidade.[16] O chamado de frequência estreita é usado em situações em que a ave sinaliza a presença de um predador e restringe as informações sobre sua própria localização, enquanto o alarme de frequência ampla é usado para atrair a atenção,[19] e pode iniciar um comportamento de mobbing. Esses chamados de “churr” variam entre os indivíduos,[20] e testes de laboratório mostram que o papa-mel-barulhento pode distinguir os chamados de diferentes aves. Portanto, isso pode ser parte integrante da complexa estrutura social da espécie.[21]
Os chamados de contato ou de facilitação social são sons de baixa frequência que são transmitidos a longas distâncias. Os chamados de “chip” são emitidos por pássaros individuais quando estão forrageando, e um chamado semelhante é emitido por filhotes que chamam em uma velocidade maior à medida que a mãe se aproxima do ninho.[18] Quando há um alto nível de atividade social, como durante disputas territoriais com conspecíficos, os chamados são uma série de notas rápidas, regulares e únicas. O papa-mel-barulhento tem uma canção de voo de exibição de acasalamento: um suave urro de notas de baixa frequência emitido durante voos curtos e ondulantes pelo macho e respondido pela fêmea com um assobio de baixa frequência.[18] O papa-mel-barulhento é encontrado em habitats de florestas abertas, onde é vantajoso fazer o chamado pelo ar, de modo a superar a atenuação do som.[22] Outro canto de exibição, descrito como “yammer”, é uma série rítmica e rápida de notas que é pronunciada durante as exibições de bico aberto e agitação das asas.[23] O papa-mel-barulhento tem uma canção descrita como “canção do amanhecer” - uma canção comunitária de notas claras e assobiadas emitidas em coro nas primeiras horas da manhã, de maio a janeiro.[18] A canção do amanhecer, que também é cantada ao anoitecer, é audível a longas distâncias e apresenta duetos que frequentemente envolvem antifonia.[16]
O filhote começa a emitir o chamado de “chip” logo após sair do ovo, e faz o chamado com frequência durante os primeiros dois terços do período de nidificação e constantemente no último terço.[23] O chamado não varia na presença de um adulto no ninho, portanto, parece provável que o chamado não seja direcionado ao pássaro adulto. O filhote fica em silêncio quando um adulto faz um chamado de alarme e emite um som de guincho quando o adulto entrega o alimento. O canto de pedido de um filhote é semelhante ao canto do filhote, mas significativamente mais alto e cobrindo uma faixa de frequência maior (o que pode torná-lo mais direcional). A taxa de vocalização, em média, é de 85 a 100 vocalizações em um minuto e, em áreas abertas, o chamado pode ser ouvido a até um quilômetro de distância. O subcanto, uma vocalização juvenil que inclui elementos de vários chamados, começa a ser emitido quando o filhote tem cerca de trinta dias de idade.[24]
O papa-mel-barulhento também produz sons não vocais clicando ou estalando o bico, geralmente durante encontros antagônicos com outras espécies de pássaros ou ao atacar um predador.[24]
Distribuição e habitat
O papa-mel-barulhento é endêmico do leste e do sudeste da Austrália, ocupando um amplo arco desde o extremo norte de Queensland, onde há populações dispersas, até Nova Gales do Sul, onde é comum e disseminado da costa até uma linha que vai de Angledool [en] a Balranald, passando por Vitória até o sudeste da Austrália Meridional e o leste da Tasmânia. Sua área de distribuição no sul da Austrália tem se expandido constantemente desde que foi registrado pela primeira vez perto de Adelaide, no início da década de 1890.[25] É sedentário em toda a sua área de distribuição.[26] O papa-mel-barulhento é territorial e o território de uma colônia é defendido agressivamente, o que levou a uma redução significativa da diversidade de aves nas áreas ocupadas pelo papa-mel-barulhento, com a exclusão de espécies menores.[27]

O papa-mel-barulhento habita principalmente florestas secas e abertas de eucalipto sem arbustos de sub-bosque. É comumente encontrado em florestas abertas de esclerófilas, incluindo aquelas em dunas costeiras ou afloramentos de granito; florestas dominadas por Corymbia maculata em cumes de montanhas e encostas expostas; florestas de eucaliptos no sopé da Cordilheira Australiana; florestas mistas de eucaliptos e ciprestes (Callitris); florestas dominadas por Eucalyptus ochrophloia [en], Acacia aneura, Acacia cambagei, Acacia harpophylla ou Eremophila; em povoamentos de Casuarina cristata [en] e aglomerados dispersos de Acacia pendula; nas bordas de bosques de Eucalyptus camaldulensis, incluindo bosques de pântano que margeiam planícies de inundação e áreas dominadas por espécies exóticas, como Fraxinus excelsior e salgueiro. Habita regularmente trechos degradados de floresta onde o sub-bosque foi desmatado, incluindo áreas queimadas recentemente, e habitats modificados, como áreas agrícolas e de pastagem com pouca madeira, reservas à beira de estradas, remanescentes de matas em cidades e vilas, e parques e jardins suburbanos com árvores e grama, mas sem arbustos densos.[28]
O papa-mel-barulhento tem se beneficiado do desbaste da mata em propriedades rurais, do pastoreio pesado que remove o sub-bosque, da fragmentação da mata que aumenta a porcentagem de efeito de borda e das práticas de paisagismo urbano que aumentam os ambientes abertos de eucalipto.[26] Ele foi descrito como uma “espécie-chave reversa”, pois está colonizando uma gama cada vez maior de habitats dominados pelo homem e excluindo agressivamente espécies menores de pássaros de ambientes urbanos.[27] Esse fenômeno também foi observado em áreas rurais. Um estudo de campo realizado nas Encostas do Sudoeste de Nova Gales do Sul mostrou que a presença do papa-mel-barulhento correspondia a um número reduzido de pássaros insetívoros, como aves dos gêneros Rhipidura e Pachycephala [en], o Myiagra inquieta e outras espécies de melifagídeos, e que essa diminuição era mais acentuada em locais com melhor acesso a água e nutrientes.[29] Embora tenha sido levantada a hipótese de que a proliferação de cultivares do gênero Grevillea com flores grandes tenha contribuído para a abundância do papa-mel-barulhento, pesquisas recentes identificaram a proliferação de áreas abertas com árvores leves e a presença de espécies de eucalipto como os fatores mais significativos para o aumento da população.[27] Híbridos de Grevillea com flores grandes podem beneficiar o papa-mel-barulhento, já que a abundância de recursos geralmente é dominada por melifagídeos maiores e agressivos,[30] e uma fonte contínua de néctar pode ser uma vantagem para as espécies não migratórias.[31] Um estudo de campo em uma região de floresta na região central de Vitória constatou que o número de papa-mel-barulhento estava correlacionado com a ocorrência de Eucalyptus leucoxylon, que produz flores (e néctar) de forma confiável todos os anos.[32] A abundância do papa-mel-barulhento é determinada principalmente pela estrutura do habitat.[33][34]
Embora a área de distribuição do papa-mel-barulhento não tenha se expandido significativamente, a densidade da população dentro dessa área aumentou substancialmente.[35] Altas densidades de papa-mel-barulhento são registradas regularmente em florestas com sub-bosque espesso no sul de Queensland, a 20 quilômetros ou mais da borda da floresta/terra agrícola. Muitos desses locais têm extensas redes de estradas usadas para o manejo florestal, além de áreas de piquenique e trilhas de caminhada para uso recreativo, e descobriu-se que esses espaços desmatados desempenham um papel na abundância dessas aves na floresta.[36] Há evidências que sugerem que densidades mais altas de estradas correspondem a níveis mais altos de população de papa-mel-barulhento.[37] O trabalho de campo em Vitória mostrou que os pássaros se infiltraram em qualquer lugar de 150 a 300 m em florestas remanescentes a partir das bordas, com maior penetração ocorrendo em áreas menos densamente florestadas. Isso tem implicações para o tamanho do habitat de floresta necessário para conter áreas livres dessas aves - cerca de 36 hectares.[38] Os projetos de revegetação que restauram a floresta de Allocasuarina luehmannii [en], uma espécie de planta da família Casuarinaceae, essencial para a sobrevivência da cacatua-negra-de-cauda-vermelha (Calyptorynchus banksii), foram interplantados com uma espécie-enfermeira, geralmente eucaliptos de crescimento rápido. As populações de papa-mel-barulhento eram mais prováveis nas florestas de Allocasuarina luehmannii onde os eucaliptos haviam sido plantados em densidades de até 16 por hectare. A presença do papa-mel-barulhento foi acompanhada por uma diferença substancial no número e nos tipos de outros pássaros encontrados na floresta.[39]
Comportamento
Organização social
O papa-mel-barulhento é uma espécie gregária, e as aves raramente são vistas sozinhas ou em duplas; elas forrageiam, se deslocam e se empoleiram em colônias que podem consistir em várias centenas de aves.[40]

Em uma colônia, um pássaro macho ocupará um “espaço de atividade”, que se sobreporá aos espaços de atividade de outros machos. Os machos com espaços de atividade sobrepostos formam associações, geralmente compostas de 10 a 25 aves. As associações são a unidade mais estável dentro da colônia. As aves também formam bandos temporários quando estão envolvidas na mesma atividade. Esses bandos, chamados de “coalizões”, geralmente são formados por cinco a oito aves, embora possam ocorrer coalizões de até 40 aves ao atacar um predador em potencial. Os membros da coalizão mudam com frequência, pois os indivíduos deixam o grupo quando ele ultrapassa o limite de seu espaço de atividade, ou quando a atividade termina ou muda, como no início da estação de reprodução.[41] As fêmeas usam espaços de atividade que se sobrepõem aos dos pássaros machos, mas não aos de outras fêmeas, de modo que as fêmeas se juntam a coalizões com machos em sua área, mas raramente haverá mais de uma fêmea na coalizão. A exclusividade dos espaços de atividade das fêmeas faz com que as fêmeas jovens sejam expulsas da colônia em que nasceram e também dificulta a conquista de um lugar em uma nova colônia. Um estudo de filhotes que sobreviveram em uma colônia até a próxima estação de reprodução constatou que todos eram pássaros machos, sugerindo que todas as fêmeas haviam morrido ou deixado a colônia.[41] A emigração de machos parece não ocorrer até que a densidade populacional da colônia atinja um nível crítico.[40]
O cuidado com os filhotes é comunitário, com os machos da colônia levando comida para os filhotes e removendo os sacos fecais. A alimentação comunitária aumenta depois que o filhote aprende a voar, quando os machos dos grupos próximos podem até levar comida para os pássaros jovens, se não forem expulsos.[42] O empoleiramento geralmente é comunitário, com dois a seis adultos e filhotes empoleirados em contato uns com os outros, geralmente perto da extremidade de um galho suspenso a até 20 metros acima do solo, dentro de seu espaço de atividade. Eles escolhem um novo local todas as noites, muitas vezes selecionando e rejeitando vários locais, e se envolvem em chamadas e perseguições agressivas quando outras aves tentam se juntar ao grupo. Eles costumam ser os últimos pássaros a se empoleirar à noite, mas parecem dormir profundamente, sem serem perturbados.[43] Eles bebem água juntos na beira de lagos e represas, geralmente empoleirando-se em um galho submerso. Eles tomam banho mergulhando de cabeça na água e, quando estão quase totalmente submersos, batem as asas vigorosamente e abaixam a cabeça sob a água. Eles sacodem o excesso de água e, em seguida, voam para um galho próximo para se arrumar. Eles foram observados usando folhagens encharcadas pela chuva ou pelo orvalho para se banharem e, em tempo seco, tomam banho de poeira em solo seco ou em lixo fino, como aparas de grama. O banho é comunitário, e as aves são estimuladas a participar observando outras.[44]
Comportamento do bando

O papa-mel-barulhento se envolve na maioria das atividades em um grupo. A postura, o forrageamento, a preparação, o banho e o banho de poeira são atividades comunitárias. O canto do amanhecer é um coro comunitário, principalmente durante a época de reprodução. A interação comunitária é facilitada por exibições ritualizadas que foram categorizadas como exibições de voo, exibições posturais e exibições faciais. Em exibições de “voo longo”, iniciadas por pássaros machos ou fêmeas, grupos de até vinte pássaros de mais de um grupo voam cerca de 40 metros acima do dossel por distâncias de até 1,5 km da colônia, chamando constantemente e não retornando à colônia por cerca de vinte minutos. Quando retornam, as aves restantes mostram sinais de agitação e, às vezes, voam para se juntar a elas. A exibição de “voo curto” é realizada pelo macho e pode ser análoga às exibições de propaganda territorial de outras aves. Em um movimento ritualizado, o papa-mel-barulhento voa de um poleiro por uma área aberta, em um padrão ondulatório rítmico, geralmente chamando durante o voo. No final da clareira, ele dá uma guinada para cima e voa silenciosamente de volta para um poleiro próximo ao ponto de partida. O “voo de cabeça para cima” é realizado pela fêmea durante o período de nidificação e pode funcionar para atrair machos ajudantes. Em sua forma mais intensa, o corpo e a cauda são mantidos quase na vertical, com as pernas balançando e a cabeça erguida e para trás. É realizada pela fêmea quando ela está selecionando o local do ninho e quando carrega o material de construção do ninho, e provavelmente tem a função de indicar a localização do ninho para outros membros do grupo.[44]

As exibições posturais incluem poses altas e baixas, apontando, com o bico aberto e agitando as asas. A “postura alta” é usada quando se está em contato próximo com outra ave e é uma ameaça leve. A ave se mantém ereta, com o pescoço e as pernas esticados, e fica de frente para a outra ave. A “postura baixa” é um gesto de submissão; a ave senta-se em um poleiro baixo, com as pernas obscurecidas por penas felpudas, e geralmente fica de costas para a outra ave. Os filhotes ameaçados por aves adultas adotam uma postura baixa e abrem muito o bico. Apontar é uma demonstração de ameaça em que a ave se estende horizontalmente, com as penas eriçadas e o bico apontado para o alvo da agressão. Às vezes, um estalo do bico acompanha o ato de apontar. A “exibição de bico aberto” é usada por subordinados em encontros entre duas aves e por uma fêmea no ninho quando outras aves se aproximam. A “exibição do bico em tridente” envolve um bico totalmente aberto com a língua levantada e saliente, e é uma exibição submissa de maior intensidade. O “agitar de asas” geralmente é realizado ao mesmo tempo que uma exibição de bico aberto. As asas são flexionadas e mantidas ligeiramente afastadas do corpo, e são batidas para fora e para cima cerca de três a seis vezes. O bater de asas pode ser acompanhado por um canto de grito. As exibições de “águia” envolvem manter as asas e a cauda abertas, em uma posição vertical ou horizontal.[44]
As exibições dos olhos são feitas expondo ou cobrindo a mancha amarela nua atrás do olho - quando as penas estão completamente arrepiadas, a mancha inteira fica visível e, quando estão afofadas, a mancha fica oculta. As exibições dos olhos são usadas em conjunto com exibições posturais, com a mancha amarela totalmente exibida por aves dominantes usando posturas de ameaça e aves imaturas que tendem a reduzir o tamanho da mancha dos olhos quando estão sendo atacadas por outros membros do grupo.[44]
Ocasionalmente, no início da estação reprodutiva, ocorrem exibições em massa, em que vinte ou trinta pássaros realizam as várias exibições de espalhamento das asas, voos curtos e chamadas constantes. As aves em exibição são atacadas por outras, e grupos de aves silenciosas, mas agitadas, observam as interações. As exibições em massa são mais comuns no início da manhã, podem durar até 40 minutos e parecem ser uma combinação de comportamento sexual e agonístico.[43]
Um “corroboree” (da palavra para uma reunião cerimonial de aborígenes australianos) é uma exibição em grupo, em que os pássaros convergem em galhos adjacentes e simultaneamente se colocam de costas, agitando as asas e exibindo o bico aberto, além do chamado yammer. O corroboree ocorre quando as aves se reúnem após uma mudança no ambiente social, como o retorno de uma ave após uma ausência, a repulsão de um intruso ou a reunião de diferentes grupos. O corroboree parece ter uma função de vínculo de par e pode envolver todos os membros de uma colônia.[44]
Comportamento agonístico
Descrito como “sempre em guerra com outros da espécie emplumada” nas primeiras anotações,[4] o papa-mel-barulhento é um dos mais agressivos dos melifagídeos. Grande parte da atividade dentro de uma colônia de papa-mel-barulhento é agonística, com perseguições, bicadas, brigas, repreensões e ataques que ocorrem com frequência durante o dia. Os pássaros se unem para atacar predadores e defender a área da colônia contra todas as outras espécies de pássaros; a espécie também é altamente agressiva intraespecificamente.[45]
As fêmeas são agressivas umas com as outras, e uma das causas da proporção de machos nas colônias pode ser a maior intolerância das fêmeas umas com as outras, expulsando os imaturos da colônia e impedindo a imigração de novas fêmeas. A agressão no ninho é comum entre os machos. Os machos adultos começam a atacar os filhotes quando eles têm cerca de 11 semanas de idade, e os agressores podem incluir machos que anteriormente cuidavam do filhote. As fêmeas adultas são menos agressivas com as aves jovens, embora as mães ocasionalmente ataquem seus próprios filhotes, e já foi registrado infanticídio. Há pouca agressividade entre machos e fêmeas além dos “voos de condução” que fazem parte do ritual de acasalamento. Em ataques diretos a aves jovens, as bicadas são direcionadas ao olho. Foi observado um comportamento agonístico entre os filhotes, com a agressão se intensificando após o aprendizado de voo e, às vezes, resultando na morte de um irmão.[46]
A colônia de papa-mel-barulhento se une para atacar intrusos e predadores interespecíficos. O papa-mel-barulhento se aproxima da ameaça e aponta, expõe as manchas dos olhos e, com frequência, bate com o bico. De cinco a quinze pássaros voam ao redor do intruso, alguns pássaros mergulham nele e se afastam ou atacam o intruso. O mobbing continua até que o invasor permaneça imóvel, como no caso de um boca-de-sapo-australiano (Podargus strigoides), ou saia da área. O mobbing de cobras e goannas é particularmente intenso, e a maioria das espécies de aves, mesmo as não predadoras, que entram no território são imediatamente perseguidas. O papa-mel-barulhento foi registrado atacando um egotelo-australiano [en] (Aegotheles cristatus) durante o dia, mergulhões, garças, patos e corvos-marinhos em lagos na borda dos territórios, rolas-de-crista (Ocyphaps lophotes), pardalotes e roselas. Mamíferos não predadores, como morcegos, gado e ovelhas também são atacados, embora com menos vigor do que as aves.[46]
Os ataques não se limitam a perseguir o intruso, e os incidentes agressivos geralmente resultam na morte do invasor. Os relatos incluem os de dois indivíduos que bicaram repetidamente um pardal-doméstico (Passer domesticus) na base do crânio e o mataram em seis minutos; um papa-mel-barulhento que agarrou um pardalote-estriado [en] (Pardalotus striatus) pela asa, enquanto outro o bicou na cabeça até ele morrer; e um martim-caçador-sagrado [en] (Todiramphus sanctus) que foi perseguido e assediado por mais de cinco horas e depois encontrado morto com o crânio fraturado.[45]
Resposta a ameaças
O papa-mel-barulhento faz chamadas de alarme mais altas em seções mais barulhentas de ambientes urbanos, como estradas principais.[47] A resposta inicial mais comum às chamadas de alarme é permanecer na área e procurar ameaças, em vez de se retirar. Um estudo realizado em Melbourne e em uma área rural próxima constatou que p papa-mel-barulhento em áreas urbanas tinha menos probabilidade de voar e, quando o fazia, voava distâncias mais curtas. Não está claro se isso é uma adaptação ou se foram os pássaros mais corajosos que se estabeleceram na cidade.[48] Um estudo de campo em Camberra descobriu que os Malurus cyaneus que viviam em áreas frequentadas pelo papa-mel-barulhento reconheciam os chamados de alarme dele e voavam, e tinham aprendido a ignorar os chamados que não eram de alarme, enquanto os que viviam em áreas não frequentadas pelo papa-mel-barulhento não respondiam aos chamados de alarme. Isso sugere que a espécie se adaptou e aprendeu a discriminar e responder às vocalizações de outra espécie.[49]
Reprodução

O papa-mel-barulhento não usa uma exibição estereotipada de cortejo sexual; as exibições podem envolver “condução”, em que o macho salta ou voa para a fêmea a uma distância de 1 a 2 metros e, se ela se afastar, ele a persegue agressivamente. A fêmea pode realizar uma “exibição de asas curvadas”, em que as asas e a cauda ficam abertas e trêmulas, com as asas arqueadas e a cabeça apontando para baixo. O macho pode adotar uma “exibição de águia” vertical ou horizontal, com as asas e a cauda bem abertas e paradas por vários segundos. A cópula é frequente e conspícua, com machos e fêmeas copulando com várias aves, enquanto outros membros da colônia se exibem ou interferem no par acasalado.[44] A cópula geralmente ocorre em galhos maiores e expostos perto do local do ninho e pode ocorrer a qualquer hora do dia, embora seja um pouco mais frequente entre 11:00 e 13:00, quando as atividades comunitárias são menos frequentes.[50] A atividade frenética de cortejo levou à especulação de que a fêmea se acasala de forma promíscua para recrutar machos para ajudar a cuidar dos filhotes, mas testes genéticos recentes mostram que 96,5% das ninhadas de papa-mel-barulhento resultam de acasalamento monogâmico e que a paternidade múltipla é rara. Uma observação de pássaros mostrou que, embora as fêmeas copulassem repetidamente, era sempre com o mesmo macho. A troca de parceiros entre ninhadas é incomum, e os pares permanecem juntos por vários anos.[51]

O papa-mel-barulhento se reproduz o ano todo, com maior atividade de julho a novembro, embora o período de pico esteja sujeito a variações sazonais, com picos acentuados na atividade de postura quando as condições são particularmente favoráveis para a criação de filhotes. O ninho é construído em árvores espinhosas ou frondosas, e o papa-mel-barulhento é frequentemente registrado nidificando em eucaliptos e também em Acacia, Araucaria, Banksia, Bursaria [en], Hibiscus, visco, Melaleuca, Pittosporum, Schinus e jacarandá-mimoso. Parece preferir uma folhagem moderadamente densa para fazer o ninho, geralmente perto da extremidade de galhos horizontais caídos. O suporte para o ninho pode ser o principal critério de um local de nidificação adequado, em vez das características da vegetação ou do local.[52] A fêmea constrói sozinha o ninho, que é profundo e em forma de xícara, tecido com galhos e gramíneas com outros materiais vegetais, pelos de animais e teias de aranha. Ocasionalmente, o ninho inclui materiais feitos pelo homem, como barbante, restos de material e papel de seda. Ele é forrado com lã, pelos, penas, flores ou penugem de plantas e acolchoado com um tapete circular tecido com fibras retiradas dos casulos de lagartas da família Lasiocampidae.[24] A fêmea do papa-mel-barulhento caminha pelo solo próximo ao local do ninho, coletando material.[52] Ela coleta material de ninhos abandonados de outras aves ou desmonta o ninho mais recente para construir um novo. A fêmea completa o ninho em cinco ou seis dias.[52] Em média, os ninhos têm um diâmetro externo de 15 a 17,8 centímetros e uma profundidade externa de 9 a 11,4 centímetros. A profundidade interna do ninho é de cerca de 5,5 centímetros.[24]

Os ovos variam muito em tamanho, forma e marcações, mas geralmente são ovais e alongados; de cor branca a creme ou rosada ou lustrosa; manchados de marrom-avermelhado a castanho ou vermelho-arroxeado, às vezes com marcações subjacentes de violeta ou cinza-arroxeado.[53] A ninhada consiste de dois a quatro ovos. A incubação é feita somente pela fêmea e o período de incubação é de cerca de dezesseis dias. A eclosão é assíncrona, com registro de até seis dias entre a eclosão do primeiro e do último filhote de uma ninhada.[54] Os filhotes estão nus na eclosão e desenvolvem uma cobertura de penugem dentro de dois a três dias. O período de aprendizagem de voo é de cerca de dezesseis dias, e os filhotes começam a encontrar comida para si mesmos entre vinte e seis e trinta dias após a fuga, mas ainda são alimentados regularmente pelos adultos até trinta e cinco dias.[54] Os filhotes deixam o ninho antes de estarem totalmente crescidos e só conseguem voar para baixo e subir. Eles não se afastam muito do ninho, retornam a ele à noite e levam algumas semanas para deixá-lo completamente. Muitos filhotes são encontrados no chão e em arbustos baixos durante esse período, onde continuam a ser cuidados até que possam subir nas árvores. Muitas vezes, essas aves são erroneamente “resgatadas”. Os filhotes procuram os irmãos, caso estejam separados, e ficam juntos por até três semanas após a saída.[55]
O papa-mel-barulhento tem alguns dos maiores tamanhos de grupo de qualquer ave de reprodução comunitária, com até vinte machos e uma fêmea cuidando de uma única ninhada.[56] Somente os machos ajudam com um ninho e, embora muitas aves possam estar associadas a uma ninhada específica, alguns machos dedicam todo o seu tempo a um único ninho, enquanto outros distribuem seus esforços de ajuda em cinco ou seis ninhos. [51] Evidências comportamentais e testes genéticos indicam que os ajudantes são machos descendentes do par reprodutor ou irmãos completos do pai.[51] Os machos quase sempre levam comida para o filhote individualmente e, se vários chegarem ao mesmo tempo, um deles passará a comida para um filhote enquanto os outros esperam. A fêmea deixa o ninho rapidamente quando um pássaro macho chega e nunca pega o alimento de um dos ajudantes. A alimentação comunitária dos filhotes aumenta após a aprendizagem do voo, e os filhotes imploram por comida com constantes chamados de “chip chip” e bocas abertas. A fêmea raramente alimenta os pássaros jovens depois que eles aprendem a voar.[55]
Predação do ninho
A reprodução cooperativa foi descrita como uma estratégia para diminuir a predação de ninhos,[57] embora um estudo não tenha encontrado nenhuma relação entre o número de ajudantes que visitam um local de ninho e seu sucesso ou fracasso. Observou-se que o papa-mel-barulhento tem uma série de estratégias para aumentar seu sucesso reprodutivo, incluindo várias ninhadas, postura de ovos no início da estação, ninhos baixos na copa das árvores e ataque contra predadores em grupo; essas medidas não garantiram o sucesso do ninho, devido à diversidade de predadores em potencial no habitat de floresta aberta do papa-mel-barulhento.[58]
Alimentação

O papa-mel-barulhento se alimenta principalmente de néctar, frutas e insetos e, ocasionalmente, de pequenos répteis ou anfíbios. Ele é arborícola e terrestre, alimentando-se na copa das árvores, em troncos e galhos e no solo. Forrageia dentro do território da colônia durante todo o ano, geralmente em grupos de cinco a oito aves, embora centenas possam se reunir em uma plantação de árvores floridas, como a Banksia. O papa-mel-barulhento coleta o néctar diretamente das flores, pendurando-se de cabeça para baixo ou atravessando galhos finos de forma acrobática para ter acesso ao néctar; pega frutas das árvores ou caídas no chão; apanha invertebrados ou caça-os; e escolhe insetos entre as folhas. Foi registrado revirando as fezes secas do emu (Dromaius novaehollandiae) e do canguru-cinza-oriental (Macropus giganteus) em busca de insetos.[59]

Em um estudo de aves que buscam alimentos em jardins suburbanos, observou-se que o papa-mel-barulhento passava mais tempo em espécies de Banksia, Grevillea e eucalipto e, quando em flor, em Callistemon, do que em outras plantas, inclusive exóticas. A maior parte do tempo foi gasta coletando a folhagem dos eucaliptos, e eles foram significativamente mais abundantes em locais onde havia eucaliptos. O papa-mel-barulhento pode satisfazer a maior parte de suas necessidades nutricionais a partir de seiva e melada coletadas da folhagem dos eucaliptos.[27] Um número menor de foi registrado em Banksia e Grevillea do que em outros grandes melifagídeos, como o papa-mel-de-asa-ruiva (Anthochaera chrysoptera) e o papa-mel-de-barbela-vermelha (Anthochaera carunculata).[60]
Estudos detalhados sobre a dieta do papa-mel-barulhento registram que ele come uma variedade de alimentos, incluindo: aranhas; insetos (crisomelídeos, joaninhas, membros da ordem Pentatomidae, formigas, larvas da ordem Lepidoptera); néctar (de jacarandá-mimoso, Erythrina variegata, Lagunaria patersonia, Melaleuca salicina, Melaleuca viminalis, eucaliptos Eucalyptus cinerea, Eucalyptus cladocalyx, Eucalyptus leucoxylon, Eucalyptus paniculata e Eucalyptus punctata, Banksia ericifolia, Banksia integrifolia, Banksia serrata, Grevillea aspleniifolia, Grevillea banksii, Grevillea hookeriana, Grevillea juniperina, Grevillea rosmarinifolia e Chaenomeles speciosa [en]); sementes de aveia, trigo e aroeira-salso; frutas de Chenopodium, Amyema e Malus sylvestris; sapos e escíncidos; e outros materiais, como pão, pedaços de carne e queijo e restos de comida.[59]
No primeiro estudo que demonstrou diferentes técnicas de aprendizagem em uma única espécie, descobriu-se que o papa-mel-barulhento empregava diferentes estratégias cognitivas, dependendo do recurso que estava forrageando. Ao procurar néctar, que não se move, mas é rapidamente esgotado, o papa-mel-barulhento usa uma estratégia baseada na memória espacial, identificando características do ambiente - uma estratégia que é eficiente em novos ambientes e não é afetada por mudanças nas atividades da ave. Ao procurar invertebrados, ele parece empregar uma estratégia diferente, baseada em regras aprendidas de movimentação de insetos (eles melhoram a localização de invertebrados com a prática). As duas estratégias diferentes implicam a existência de mecanismos cognitivos adaptados, capazes de responder adequadamente a diferentes contextos de forrageamento.[61]
Status de conservação
Abundante em toda a sua significativa área de distribuição, o papa-mel-barulhento é considerado espécie pouco preocupante para a conservação,[1] e suas densidades populacionais extremas em algumas áreas constituem, na verdade, uma ameaça a outras espécies. A forte correlação entre a presença dessas aves e a ausência de diversidade aviária foi bem documentada.[62] O papel desempenhado pelo papa-mel-barulhento no declínio acentuado de muitas aves florestais, seu impacto sobre espécies ameaçadas de extinção com requisitos de forrageamento semelhantes e o nível de danos às folhas que levam à morte que acompanha a exclusão de aves insetívoras de florestas remanescentes significa que qualquer estratégia para restaurar a diversidade aviária precisará levar em conta o gerenciamento das populações de papa-mel-barulhento.[63] Alguns projetos de restauração de habitat e revegetação aumentaram inadvertidamente o problema do papa-mel-barulhento ao estabelecer o habitat de eucalipto aberto que ele prefere. Um dos focos de muitos projetos de regeneração tem sido o estabelecimento de corredores de habitat que conectam trechos de florestas remanescentes e o uso de eucaliptos como espécies de crescimento rápido. Ambas as práticas têm um valor ecológico sólido, mas permitem que o papa-mel-barulhento se prolifere, de modo que os esforços de conservação estão sendo modificados com o plantio de um sub-bosque arbustivo com os eucaliptos e evitando a criação de saliências estreitas, cantos ou aglomerados de árvores nos corredores de vegetação.[35][64] Um estudo de campo realizado nas Terras Altas do Sul descobriu que o papa-mel-barulhento tendia a evitar áreas dominadas por acácias, cujas espécies na área de estudo tinham folhas bipinadas. Por isso, os autores propuseram que os projetos de revegetação incluíssem pelo menos 15% de espécies de Acacia com folhas bipinadas, se possível, bem como plantas arbustivas de sub-bosque.[65]
É improvável que a translocação do papa-mel-barulhento seja uma solução para sua superabundância em habitats remanescentes. Em um estudo realizado em Vitória, na Austrália, no qual as aves foram colocadas em faixas e realocadas, as colônias se mudaram para a área agora despovoada, mas logo retornaram aos seus territórios originais. As aves translocadas não se estabeleceram em um novo território. Elas não foram assimiladas pelas populações residentes de papa-mel-barulhento, mas, em vez disso, vagaram por até 4,2 quilômetros do ponto de soltura, passando por um habitat aparentemente adequado ocupado por outros grupos de papa-mel-barulhento - pelo menos nos primeiros 50 dias após a translocação. Duas aves com dispositivos de rastreamento por rádio percorreram 18 quilômetros de volta ao local de captura.[66] Embora eles sejam protegidos em toda a Austrália e seja necessária uma autorização para abatê-los, o abate foi proposto como o método mais humano e prático de reduzir seu impacto, especialmente quando combinado com a reabilitação do habitat para atender a uma variedade maior de pássaros.[63] Um abate não sancionado foi realizado em uma propriedade rural privada em 1991 e 1992, o que, combinado com plantações extensas e densas de árvores nativas, supostamente resultou em um aumento na diversidade de espécies.[67]
Referências
- ↑ a b c BirdLife International (2018). «Manorina melanocephala». Lista Vermelha de Espécies Ameaçadas. 2018: e.T22704433A132071923. doi:10.2305/IUCN.UK.2018-2.RLTS.T22704433A132071923.en
 . Consultado em 19 de novembro de 2021
. Consultado em 19 de novembro de 2021
- ↑ Latham, John (1801). Supplementum Indicis Ornithologici, sive Systematis Ornithologiae (em latim). London: G. Leigh, J. & S. Sotheby. pp. xxviii, xxxiv, xxxiii, xxxv
- ↑ Salomonsen, F. (1967). «Family Maliphagidae, Honeyeaters». In: Paynter, R.A. Jnr. Check-list of birds of the world (Volume 12). 12. Cambridge, Mass.: Museum of Comparative Zoology. p. 400
- ↑ a b Sharpe, Richard Bowdler (1906). The History of the Collections Contained in the Natural History Departments of the British Museum. Volume 2. 2. London: British Museum. p. 126
- ↑ The Natural History Museum, London (2007). «'Blue-cheeked Bee Eater', native name 'Der-ro-gang'». First Fleet Artwork Collection. The Natural History Museum, London. Consultado em 2 de janeiro de 2012
- ↑ Gould, John (1865). Handbook to the Birds of Australia. Volume 1. 1 [Darwin's copy]. London: J. Gould. p. 574
- ↑ Koch, Harold; Hercus, Luise (2009). Aboriginal Placenames: Naming and Re-Naming the Australian Landscape. Canberra, Australian Capital Territory: ANU E Press. p. 104. ISBN 978-1-921666-08-7
- ↑ Schodde, R.; Bock, W.J. (2008). «Case 3418. Specific Names of Nine Australian Birds (Aves): Proposed Conservation». Bulletin of Zoological Nomenclature. 65 (1): 35–41. ISSN 0007-5167. doi:10.21805/bzn.v65i1.a9
- ↑ International Commission on Zoological Nomenclature (2009). «Opinion 2240 (Case 3418). Specific names of nine Australian birds (Aves) conserved». Bulletin of Zoological Nomenclature. 66 (4): 375–78. ISSN 0007-5167. doi:10.21805/bzn.v66i4.a21
- ↑ Liddell, Henry George; Scott, Robert (1980) [1871]. A Greek-English Lexicon abridged ed. Oxford, United Kingdom: Oxford University Press. pp. 374, 431. ISBN 978-0-19-910207-5
- ↑ a b c d Schodde, Richard; Mason, Ian J. (1999). The Directory of Australian Birds: Passerines. A Taxonomic and Zoogeographic Atlas of the Biodiversity of Birds in Australia and its Territories. Collingwood, Victoria: CSIRO Publishing. p. 266. ISBN 978-0-643-10293-4
- ↑ a b c d Higgins, Peter & Steele 2001, p. 648.
- ↑ Barker, F. Keith; Cibois, Alice; Schikler, Peter; Feinstein, Julie; Cracraft, Joel (2004). «Phylogeny and Diversification of the Largest Avian Radiation». Proceedings of the National Academy of Sciences. 101 (30): 11040–45. Bibcode:2004PNAS..10111040B. PMC 503738. PMID 15263073. doi:10.1073/pnas.0401892101
- ↑ Officer 1965, p. 72.
- ↑ a b Higgins, Peter & Steele 2001, p. 647.
- ↑ a b c Higgins, Peter & Steele 2001, p. 640.
- ↑ Officer 1965, p. 70.
- ↑ a b c d Jurisevic, Mark A.; Sanderson, Ken J. (1994). «The Vocal Repertoires of Six Honeyeater (Meliphagidae) Species from Adelaide, South Australia». Emu. 94 (3): 141–48. Bibcode:1994EmuAO..94..141J. ISSN 0158-4197. doi:10.1071/MU9940141
- ↑ Klump, G.M; Kretzschmar, E.; Curio, E. (1986). «The Hearing of an Avian Predator and its Avian Prey». Behavioral Ecology and Sociobiology. 18 (5): 317–23. ISSN 0340-5443. doi:10.1007/BF00299662
- ↑ Kennedy, R. A. W.; Evans, C. S.; McDonald, P. G. (2009). «Individual Distinctiveness in the Mobbing Call of a Cooperative Bird, the Noisy Miner Manorina melanocephala». Journal of Avian Biology. 40 (5): 481–490. doi:10.1111/j.1600-048X.2008.04682.x
- ↑ McDonald, P. G. (2012). «Cooperative Bird Differentiates Between the Calls of Different Individuals, Even When Vocalizations Were From Completely Unfamiliar Individuals». Biology Letters. 8 (3): 365–368. PMC 3367754. PMID 22258445. doi:10.1098/rsbl.2011.1118
- ↑ Cosens, Susan E.; Falls, J. Bruce (1984). «A Comparison of Sound Propagation and Song Frequency in Temperate Marsh and Grassland Habitats». Behavioral Ecology and Sociobiology. 15 (3): 161–70. JSTOR 4599714. doi:10.1007/BF00292970
- ↑ a b Higgins, Peter & Steele 2001, p. 641.
- ↑ a b c d Higgins, Peter & Steele 2001, p. 642.
- ↑ Higgins, Peter & Steele 2001, p. 628.
- ↑ a b Higgins, Peter & Steele 2001, p. 629.
- ↑ a b c d Ashley, Lisa C.; Major, Richard E.; Taylor, Charlotte E. (2009). «Does the Presence of Grevilleas and Eucalypts in Urban Gardens Influence the Distribution and Foraging Ecology of Noisy Miners?». Emu. 109 (2): 135–42. Bibcode:2009EmuAO.109..135A. ISSN 0158-4197. doi:10.1071/MU07043
- ↑ Higgins, Peter & Steele 2001, p. 627–28.
- ↑ Montague-Drake, R. M.; Lindenmayer, D. B.; Cunningham, R. B.; Stein, J. A. (2011). «A Reverse Keystone Species Affects the Landscape Distribution of Woodland Avifauna: A Case Study Using the Noisy Miner (Manorina melanocephala) and other Australian Birds». Landscape Ecology. 26 (10): 1383–1394. Bibcode:2011LaEco..26.1383M. doi:10.1007/s10980-011-9665-4
- ↑ Ford, Hugh A.; Paton, David C. (1976). «Resource Partitioning and Competition in Honeyeaters of the Genus Meliphaga». Australian Journal of Ecology. 1 (4): 281–87. Bibcode:1976AusEc...1..281F. doi:10.1111/j.1442-9993.1976.tb01118.x
- ↑ Griffioen, Peter A.; Clarke, Michael F. (2002). «Large-scale Bird-movement Patterns Evident in Eastern Australian atlas data». Emu. 102 (1): 99–125. Bibcode:2002EmuAO.102...99G. doi:10.1071/MU01024
- ↑ Oldland, J. M.; Taylor, R. S.; Clarke, M. F. (2009). «Habitat Preferences of the Noisy Miner (Manorina melanocephala) – A Propensity for Prime Real Estate?». Austral Ecology. 34 (3): 306–316. Bibcode:2009AusEc..34..306O. doi:10.1111/j.1442-9993.2009.01931.x
- ↑ Catterall, Carla P. (2004). «Birds, Garden Plants and Suburban Bushlots: Where Good Intentions Meet Unexpected Outcomes». In: D. Lunney; S. Burgin. Urban Wildlife: More Than Meets the Eye. Sydney: Royal Zoological Society of New South Wales. pp. 21–31. ISBN 978-0-9586085-7-2
- ↑ Daniels, Grant; Kirkpatrick, Jamie (2006). «Does Variation in Garden Characteristics Influence the Conservation of Birds in Suburbia?». Biological Conservation. 133 (3): 326–35. Bibcode:2006BCons.133..326D. doi:10.1016/j.biocon.2006.06.011
- ↑ a b Clarke, Michael F.; Taylor, Richard; Oldland, Joanne; Grey, Merilyn J.; Dare, Amanda (2007). «Challenges in Managing Miners». The Victorian Naturalist. 124 (2): 102–05. ISSN 0042-5184
- ↑ Piper, Scott D.; Catterall, Carla P. (2006). «Impacts of Picnic Areas on Bird Assemblages and Nest Predation Activity Within Australian Eucalypt Forests». Landscape and Urban Planning. 78 (3): 251–62. doi:10.1016/j.landurbplan.2005.09.001
- ↑ Maron, Martine (2009). «Nesting, Foraging and Aggression of Noisy Miners Relative to Road Edges in an Extensive Queensland Forest». Emu. 109 (1): 75–81. Bibcode:2009EmuAO.109...75M. doi:10.1071/MU08064
- ↑ Clarke, M. F.; Oldland, J. M. (2007). «Penetration of Remnant Edges by Noisy Miners (Manorina melanocephala) and Implications for Habitat Restoration». Wildlife Research. 34 (4): 253–261. doi:10.1071/WR06134
- ↑ Maron, Martine (2007). «Threshold Effect of Eucalypt Density on an Aggressive Avian Competitor». Biological Conservation. 136 (1): 100–07. Bibcode:2007BCons.136..100M. doi:10.1016/j.biocon.2006.11.007
- ↑ a b Higgins, Peter & Steele 2001, p. 632.
- ↑ a b Higgins, Peter & Steele 2001, p. 631.
- ↑ Higgins, Peter & Steele 2001, p. 692.
- ↑ a b Higgins, Peter & Steele 2001, p. 634.
- ↑ a b c d e f Dow, Douglas D. (1975). «Displays of the Honeyeater Manorina melanocephala». Ethology. 38 (1): 70–96. PMID 1237204. doi:10.1111/j.1439-0310.1975.tb01993.x
- ↑ a b Dow, Douglas D. (1979). «Agonistic and Spacing Behaviour of the Noisy Miner Manorina melanocephala, a Communally Breeding Honeyeater». Ibis. 121 (4): 423–36. doi:10.1111/j.1474-919X.1979.tb06683.x
- ↑ a b Higgins, Peter & Steele 2001, p. 635.
- ↑ Lowry, H. L. N. L. N.; Lill, A.; Wong, B. B. M. (2012). Iwaniuk, Andrew, ed. «How Noisy Does a Noisy Miner Have to Be? Amplitude Adjustments of Alarm Calls in an Avian Urban 'Adapter'». PLOS ONE. 7 (1): e29960. Bibcode:2012PLoSO...729960L. PMC 3251594. PMID 22238684. doi:10.1371/journal.pone.0029960
- ↑ Lowry, Hélène L. N.; Lill, Alan; Wong, Bob B. M. (2011). «Tolerance of Auditory Disturbance by an Avian Urban Adapter, the Noisy Miner». Ethology. 117 (6): 490–97. Bibcode:2011Ethol.117..490L. doi:10.1111/j.1439-0310.2011.01902.x
- ↑ Magrath, Robert D.; Bennett, Thomas H. (2011). «A Micro-geography of Fear: Learning to Eavesdrop on Alarm Calls of Neighbouring Heterospecifics». Proceedings of the Royal Society B: Biological Sciences. 279 (1730): 902–09. PMC 3259928. PMID 21849313. doi:10.1098/rspb.2011.1362
- ↑ Higgins, Peter & Steele 2001, p. 637–38.
- ↑ a b c Pōldmaa, Tarmo; Holder, Karen (1997). «Behavioural Correlates of Monogamy in the Noisy Miner, Manorina melanocephala». Animal Behaviour. 54 (3): 571–78. PMID 9299042. doi:10.1006/anbe.1996.0451
- ↑ a b c Dow, Douglas D. (1978). «Breeding Biology and Development of the Young of Manorina melanocephala, a Communally Breeding Honeyeater». Emu. 78 (4): 207–22. Bibcode:1978EmuAO..78..207D. doi:10.1071/MU9780207
- ↑ Beruldsen 1980, p. 368.
- ↑ a b Higgins, Peter & Steele 2001, p. 643.
- ↑ a b Higgins, Peter & Steele 2001, p. 639.
- ↑ Conrad, Kelvin F.; Clarke, Michael F.; Robertson, Raleigh J.; Boag, Peter T. (1998). «Paternity and the Relatedness of Helpers in the Cooperatively Breeding Bell Miner». The Condor. 100 (2): 342–49. JSTOR 1370275. doi:10.2307/1370275
- ↑ Roper, James J.; Goldstein, Rachel R (1997). «A Test of the Skutch Hypothesis: Does Activity at Nests Increase Nest Predation Risk?». Journal of Avian Biology. 28 (2): 111–16. JSTOR 3677304. doi:10.2307/3677304
- ↑ Arnold, Kathryn E (2000). «Strategies of the Cooperatively Breeding Noisy Miner to Reduce Nest Predation». Emu. 100 (4): 280–85. Bibcode:2000EmuAO.100..280A. doi:10.1071/MU9909
- ↑ a b Higgins, Peter & Steele 2001, p. 630.
- ↑ French, Kris; Major, Richard; Hely, K. (2005). «Use of Native and Exotic Garden Plants by Suburban Nectarivorous Birds». Biological Conservation. 121 (4): 545–59. Bibcode:2005BCons.121..545F. doi:10.1016/j.biocon.2004.06.004
- ↑ Sulikowski, Danielle; Burke, Darren (2011). «Movement and Memory: Different Cognitive Strategies are Used to Search for Resources with Different Natural Distributions». Behavioral Ecology and Sociobiology. 65 (4): 621–31. doi:10.1007/s00265-010-1063-4
- ↑ Grey, Merilyn J.; Clarke, Michael F.; Loyn, Richard H. (1998). «Influence of the Noisy Miner Manorina melanocephala on Avian Diversity and Abundance in Remnant Grey Box Woodland». Pacific Conservation Biology. 4: 55–69. doi:10.1071/PC980055
- ↑ a b Grey, Merilyn; Clarke, Michael (2011). The Noisy Miner: Challenges in Managing an Overabundant Species (PDF). Melbourne, Victoria: La Trobe University. Consultado em 18 de dezembro de 2011
- ↑ Taylor, R. S.; Oldland, J. M.; Clarke, M. F. (2008). «Edge Geometry Influences Patch-level Habitat Use by an Edge Specialist in South-eastern Australia». Landscape Ecology. 23 (4): 377–389. Bibcode:2008LaEco..23..377T. doi:10.1007/s10980-008-9196-9
- ↑ Hastings, R. A.; Beattie, A. J. (2006). «Stop the Bullying in the Corridors: Can Including Shrubs Make Your Revegetation More Noisy Miner Free?». Ecological Management and Restoration. 7 (2): 105–112. Bibcode:2006EcoMR...7..105H. doi:10.1111/j.1442-8903.2006.00264.x
- ↑ Clarke, Michael F.; Schedvin, Natasha (1997). «An Experimental Study of the Translocation of Noisy Miners Manorina Melanocephala and Difficulties Associated with Dispersal». Biological Conservation. 80 (2): 161–67. Bibcode:1997BCons..80..161C. doi:10.1016/S0006-3207(96)00075-4
- ↑ Debus, S.J.S. (2008). «The Effect of Noisy Miners on Small Bush Birds: an Unofficial Cull and Its Outcome» (PDF). Pacific Conservation Biology. 14 (3): 185–90. doi:10.1071/PC080185
Bibliografia
- Beruldsen, Gordon (1980). A Field Guide to Nests and Eggs of Australian Birds. Adelaide, South Australia: Rigby. ISBN 978-0-7270-1202-9
- Higgins, Peter; Peter, J.M.; Steele, W.K. (2001). «Manorina melanocephala Noisy Miner». Handbook of Australian, New Zealand and Antarctic Birds Volume 5, Tyrant-flycatchers to Chats. Melbourne, Victoria: Oxford University Press. pp. 626–50. ISBN 978-0-19-553258-6
- Officer, Hugh R. (1965). Australian Honeyeaters. Melbourne, Victoria: The Bird Observers Club, Melbourne. ISBN 978-0-909711-03-0
Ligações externas
- «Noisy miner». Birds in Backyards. Consultado em 18 de dezembro de 2011