Normandina pulchella
Normandina pulchella
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
.jpg) | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Nome binomial | |||||||||||||||||
| Normandina pulchella (Borrer) Nyl. (1861) | |||||||||||||||||
| Sinónimos[2] | |||||||||||||||||
| |||||||||||||||||
Normandina pulchella é uma espécie de líquen esquamuloso da família Verrucariaceae. Esta espécie cosmopolita está amplamente distribuída em ambos os hemisférios, prosperando em micro-habitats úmidos. Prefere árvores decíduas e rochas cobertas por musgos, frequentemente colonizando musgos e briófitas. Ocasionalmente, cresce sobre cascas nuas ou outros líquens. Suas características distintivas incluem escamas verde azuladas com margens nitidamente elevadas, ausência de reatividade a testes químicos padrão e crescimento em habitats úmidos. Inicialmente, acreditava-se que a alga verde Nannochloris normandinae fosse seu fotobionte. Estudos mais recentes, no entanto, revisaram esse entendimento, identificando Diplosphaera chodatii como o parceiro algal.
Descrita cientificamente pela primeira vez pelo botânico inglês William Borrer em 1831, a posição de Normandina pulchella na família Verrucariaceae foi esclarecida em 2010 por meio de análises de filogenética molecular, resolvendo incertezas taxonômicas de longa data. Classificações anteriores variavam amplamente, colocando N. pulchella em grupos como Basidiomycota (como um basidiolíquen) e Fungi incertae sedis, devido a diferentes interpretações dos peritécios (estruturas reprodutivas) encontrados no líquen. Essas discrepâncias resultaram da confusão sobre se os peritécios pertenciam ao próprio líquen ou se estavam associados a um fungo liquenícola parasita.
Sistemática
Histórico taxonômico

O líquen foi formalmente descrito em 1831 pelo botânico inglês William Borrer, que o classificou no gênero Verrucaria. Ele criou o nome vernacular "pequena Verrucaria de folhas com película" devido à sua morfologia distinta.[3] O epíteto específico pulchella é o diminutivo do latim de pulchra, que significa "belo" ou "formoso".[4] Borrer observou que o líquen era caracterizado por escamas finas, membranosas, cinza-esverdeadas, com formas que variavam de arredondadas e lisas a aglomeradas, onduladas e lobadas, adornadas com grânulos pulverulentos. A face inferior dessas escamas apresenta uma coloração marrom pálida e fibras lanosas. Ele descreveu os tubérculos do líquen como quase globulares e pretos, revelando apenas o ápice através da superfície do talo, expondo um núcleo gelatinoso acastanhado com um poro central. Borrer notou a ocorrência frequente do líquen em árvores musgosas em Sussex, Inglaterra, e que se desenvolveu na espécie de hepática folhosa Jungermannia dilatata, formando manchas amplas, mas frequentemente interrompidas. Ele creditou a Ellen Hutchins a descoberta inicial da espécie em uma montanha perto de Bantry [en], na Irlanda, crescendo sobre Lichen plumbeus em caules de urze. Borrer destacou a singularidade do líquen:[5]
Esta curiosa pequena produção é tão diferente de qualquer outro líquen que seu próprio gênero teria permanecido incerto, não fosse a afortunada descoberta dos tubérculos por Miss Hutchins. Acharius, a quem espécimes de Sussex foram comunicados, pensou que se tratava de uma Thelephora, excluindo-a até mesmo da ordem natural à qual acreditamos que pertence.
Apesar de sua prevalência em Sussex, Borrer mencionou que o líquen parecia ter sido ignorado em outros lugares.[5] Naquele mesmo ano, ele propôs transferi-lo para o gênero Endocarpon, mas essa mudança foi nomenclaturalmente inválida, pois o nome já havia sido usado para uma espécie diferente, violando as diretrizes do Código Internacional de Nomenclatura Botânica.[3] O taxonomista finlandês William Nylander transferiu o táxon para o gênero Normandina em 1861.[6]
Classificação
A posição taxonômica de N. pulchella foi alvo de debates entre liquenólogos por muitas décadas.[7] Embora o líquen produza ascósporos, a origem desses esporos e suas estruturas associadas era historicamente controversa. Antes dos estudos moleculares, os pesquisadores propuseram várias classificações, incluindo sua colocação nos Basidiomycota, devido a semelhanças morfológicas com certos basidiolíquens, particularmente Coriscium viride, associado ao basidiomiceto Lichenomphalia hudsoniana. A controvérsia central girava em torno dos peritécios encontrados no talo, que mais tarde foram atribuídos ao fungo parasita Sphaerulina chlorococca, e não a N. pulchella. Alguns pesquisadores consideravam essas estruturas como órgãos reprodutivos do próprio líquen, enquanto outros as interpretavam como pertencentes a um fungo parasita.[7] A descoberta de que Normandina não possui septos doliporos — uma característica chave dos basidiomicetos — levantou dúvidas sobre sua classificação como basidiolíquen, mas sua posição sistemática exata permaneceu incerta até 2010, quando análises de filogenética molecular estabeleceram definitivamente sua posição na família Verrucariaceae. Essa classificação foi reforçada pela presença de características típicas de ascomicetos, como corpos de Woronin e septos perfurados simples.[8]
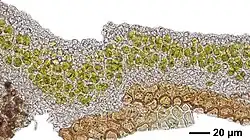
Um estudo citológico revelou que N. pulchella possui um talo heterômero, onde os componentes fúngico e algal são organizados em camadas distintas. Essa estrutura desempenha um papel fundamental em seus processos vitais, distinguindo-o dos basidiolíquens. Especificamente, a medula, ou camada interna, contém filamentos fúngicos hialinos (transparentes) entrelaçados ao redor de pequenos grupos de células algais. Essas células possuem paredes espessas e um cloroplasto lobado singular contendo um pirenoide metamérico distinto — uma estrutura que auxilia na formação de amido, junto com pequenos grãos de amido e plastoglóbulos contendo gordura.[8]
O parceiro fúngico, ou micobionte, apresenta hifas com septos perfurados simples, um tipo de divisão celular interna nas hifas, acompanhado por corpos de Woronin — organelas exclusivas dos ascomicetos, um grande grupo fúngico. Além disso, a relação próxima, mas não invasiva, entre as células fúngicas e algais sugeriu uma associação mutualística típica de ascolíquens.[8] Essas evidências citológicas, juntamente com a análise de filogenética molecular de 2010 de Lucia Muggia e colegas, estabeleceram de forma conclusiva a posição de N. pulchella na família Verrucariaceae.[9]
Descrição

Normandina pulchella possui um talo esquamuloso, composto por pequenas formações semelhantes a escamas chamadas escamulas. As escamulas apresentam cores que variam de glauco - verde azulado ou cinza - a cinza claro ou cinza esverdeado, intensificando-se para um verde mais rico quando umedecidas. Cada escamula pode atingir até 5 mm de diâmetro e pode incluir um ou mais lóbulos (projeções do talo) amplamente arredondados, com até 1,7 mm de diâmetro, reminiscentes de formas de concha ou orelha. A superfície superior desses lóbulos apresenta sulcos concêntricos, enquanto as bordas são nitidamente definidas e elevadas, geralmente com 50 a 100 μm de largura. As escamulas podem estar dispersas ou densamente agrupadas na superfície do líquen.[10] O talo fica intimamente aderido ao substrato.[11] Internamente, o talo exibe uma estrutura heterômera, com camadas distintas de componentes fúngicos e algais.[7] A medula contém filamentos fúngicos transparentes que formam uma rede ao redor de grupos de células algais. Essas hifas fúngicas apresentam septos perfurados simples com corpos de Woronin distintos, enquanto as células algais são caracterizadas por paredes celulares espessas e cloroplastos lobados únicos contendo pirenoides.[8]
Para reprodução vegetativa, N. pulchella desenvolve sorálios — estruturas nas superfícies e bordas dos lóbulos — que liberam partículas granulares conhecidas como sorédios. Esses sorálios são verdes ou combinam com a cor dos lóbulos e contêm sorédios granulares medindo 40 a 80 μm de diâmetro. A face inferior do líquen apresenta uma aparência esbranquiçada, ligeiramente feltrada (tomentosa), e adere ao substrato por meio de numerosos filamentos fúngicos, ou hifas.[10] Rizinas não estão presentes nesta espécie.[11]
Suas estruturas produtoras de esporos, ou ascomas, aparecem como peritécios globulares a ligeiramente cônicos, semi-imersos, com pigmentação laranja-acastanhada moderada na parede de textura angular. Eles são caracterizados por perífises abundantes, ausência de paráfises e algas himeniais, e um gel himenial I+ vermelho/KI+ azul.[7] Assemelham-se aqueles em espécies relacionadas, mas distinguem-se por sua natureza uniformemente pigmentada e coesa sob análise microscópica. Os esporos medem tipicamente 29 a 37 por 6 a 7 μm, geralmente com sete septos internos,[10] embora alguns espécimes apresentem esporos predominantemente com 4 a 5 septos, medindo 17 a 29 por 5 a 7 μm.[7] Análises químicas, particularmente por cromatografia em camada delgada, identificam o zeorin como um metabólito secundário, mas N. pulchella não reage a testes químicos padrão.[10]
Espécies semelhantes
As escamulas de Lichenomphalina hudsoniana compartilham as bordas nítidas e bem definidas características de N. pulchella. No entanto, podem ser distinguidas pela ausência de sorálios, pela presença de córtex superior e inferior, e pela adaptação a ambientes ártico-alpinos, geralmente crescendo em solos turfosos ou madeira em decomposição. Em contraste, N. pulchella tende a desenvolver sorálios mais pronunciados em ambientes sombreados e úmidos, divergindo dos habitats preferidos de Lichenomphalina hudsoniana. Além disso, os ascomas de N. pulchella são mais prevalentes e maiores em regiões tropicais do que em temperadas, e frequentemente não contêm sorédios. Isso sugere que fatores ambientais ou diferenças taxonômicas subjacentes podem influenciar as variações observadas entre essas espécies em diferentes locais.[10][11]
Fotobionte
Em 1981, Elisabeth Tschermak-Woess identificou Nannochloris normandinae como o parceiro fotobionte associado a Normandina pulchella.[12] Estudos subsequentes, no entanto, foram menos conclusivos sobre o papel de Nannochloris.[13] Em 2011, pesquisas de Thüs e colegas revelaram que Diplosphaera, e não Nannochloris, estava presente em dez espécimes de Normandina examinados.[14] Em 2020, Pröschold e Darienko demonstraram que Nannochloris normandinae é, na verdade, sinônimo de Diplosphaera chodatii da ordem Prasiolales, resolvendo o aparente conflito em estudos anteriores e confirmando D. chodatii como o fotobionte de N. pulchella.[15][16]
Habitat e distribuição

Normandina pulchella tem uma distribuição cosmopolita, crescendo em diversos climas e regiões.[10] O liquenólogo sueco Gunnar Degelius, em seu estudo fitogeográfico de 1934, destaca a distribuição oceânica da espécie na Europa. Ela ocupa predominantemente áreas costeiras no norte da Europa, incluindo Escandinávia e as Ilhas Britânicas, e se estende a regiões costeiras e montanhosas na Áustria, Baviera, França, República Tcheca e Eslováquia e alguns locais mediterrâneos.[17] Em contraste, a análise de William Louis Culberson e Mason Hale em 1966 sobre sua presença na América do Norte destacou sua prevalência nas regiões montanhosas ocidentais e nas montanhas dos Apalaches, sem indicar um padrão de distribuição oceânica.[18] Sua distribuição na América do Norte se estende ao norte até o Alasca,[19] embora, em 2022, Alan Orange tenha mostrado que algumas coleções de lá representam uma espécie diferente encontrada nas Américas, provisoriamente chamada N. americana.[20]
Ecologicamente, Normandina pulchella prefere árvores decíduas e rochas cobertas por musgos em florestas e parques, frequentemente colonizando musgos, briófitas e, ocasionalmente, cascas nuas.[10] Também cresce frequentemente sobre outros líquens, particularmente aqueles que contêm cianobactérias,[21] como Fuscopannaria, Pannaria, Parmeliella, Pectenia e Peltigera. Sua presença está aumentando no sul e oeste da Grã-Bretanha e em toda a Irlanda, refletindo uma distribuição em expansão.[10]
Está listada como uma espécie vulnerável na Lista Vermelha Regional da Finlândia devido à sua população conhecida reduzida.[22]
Interações com espécies
Entre os fungos liquenícolas associados especificamente a N. pulchella, várias espécies apresentam interações únicas. Capronia normandinae é caracterizada por suas estruturas pretas, superficiais, semelhantes a pelos, conhecidas como peritécios setosos. Cladophialophora normandinae distingue-se por seus esporocarpos pretos, chamados esporodóquios, que desempenham um papel em seu ciclo reprodutivo. Além disso, Tremella normandinae é notada por produzir crescimentos pálidos e inchados, chamados galhas, indicativos de sua relação parasitária com o líquen. Outro parasita, Globosphaeria jamesii, também interage com Normandina pulchella.[10][23]
Lawreymyces pulchellae é um fungo que vive oculto nos tecidos de Normandina pulchella, um dos vários basidiomicetos crípticos descobertos recentemente em associações com líquens. Descrito em 2017 como parte da família Corticiaceae [en], ele difere dos fungos liquenícolas típicos por não deixar sinais visíveis de sua presença, sendo detectável apenas por métodos moleculares. Pertence a um grupo de espécies de Lawreymyces que se associam especificamente com líquens da família Verrucariaceae, particularmente Normandina e Agonimia. Essa relação é paralela à de Cyphobasidium em líquens Parmeliaceae [en]. A espécie é identificável apenas por sua sequência de DNA única no espaçador interno transcrito.[24]
Ver também
Referências
- ↑ Nylander, William (1855). «Additamentum in floram cryptogamicam Chilensem». Annales des Sciences Naturelles. Botanique. 4 (em latim). 3: 145–187 [151]
- ↑ «Synonymy. Current Name: Normandina pulchella (Borrer) Nyl., Annls Sci. Nat., Bot., sér. 4 15: 382 (1861)». Species Fungorum. Consultado em 28 de setembro de 2025
- ↑ a b «Record Details: Endocarpon pulchellum Borrer, in Hooker & Sowerby, Suppl. Engl. Bot. 1: tab. 2602 (1831)». Index Fungorum. Consultado em 28 de setembro de 2025
- ↑ Bomfleur, Benjamin; Grimm, Guido W.; McLoughlin, Stephen (2015). «Osmunda pulchella sp. nov. from the Jurassic of Sweden—reconciling molecular and fossil evidence in the phylogeny of modern royal ferns (Osmundaceae)». BMC Evolutionary Biology. 15 (1): e15. Bibcode:2015BMCEE..15..126B. PMC 4487210
 . PMID 26123220. doi:10.1186/s12862-015-0400-7
. PMID 26123220. doi:10.1186/s12862-015-0400-7
- ↑ a b Borrer, William (1831). Supplement to the English Botany of the late Sir J. E. Smith and Mr. Sowerby. 1. London: J. D. C. and C. E. Sowerby. tab. 2602, fig. 1
- ↑ Nylander, William (1861). «Additamentum ad lichenographiam Andium Boliviensium» [Supplement to the study of lichens of the Bolivian Andes]. Annales des Sciences Naturelles. Botanique. 4 (em latim). 15: 365–382 [382]
- ↑ a b c d e Frisch, Andreas; Ohmura, Yoshihito (2015). «The phylogenetic position of Normandina simodensis (Verrucariaceae, lichenized Ascomycota)». Bulletin of the National Museum of Nature and Science, Series B (Botany). 41 (1): 1–7
- ↑ a b c d Mares, Donatello; Fasulo, Maria P.; Bruni, Alessandro (1993). «Contribution to the study of Normandina pulchella: a cytological approach». Orsis. 8: 33–40
- ↑ Muggia, Lucia; Gueidan, Cécile; Grube, Martin (2010). «Phylogenetic placement of some morphologically unusual members of Verrucariales». Mycologia. 102 (4): 835–846. PMID 20648751. doi:10.3852/09-153
- ↑ a b c d e f g h i Orange, A.; Cannon, P.; Prieto, M.; Coppins, B.; Sanderson, N.; Simkin, J. (2023). Verrucariales: Verrucariaceae (PDF). Col: Revisions of British and Irish Lichens. 31. [S.l.]: British Lichen Society. p. 43
- ↑ a b c McMullin, R. Troy (2023). Lichens. The Macrolichens of Ontario and the Great Lakes Region of the United States. [S.l.]: Firefly Books. p. 311. ISBN 978-0-228-10369-1
- ↑ Tschermak-Woess, E. (1981). «Zur Kenntnis der Phycobionten von Lobaria linita und Normandina pulchella» [Sobre o conhecimento dos fitobiontes de "Lobaria linita" e "Normandina pulchella]. Nova Hedwigia (em alemão). 35: 63–73
- ↑ Lohtander, Katileena; Oksanen, Ilona; Rikkinen, Jouko (2003). «Genetic diversity of green algal and cyanobacterial photobionts in Nephroma (Peltigerales)». The Lichenologist. 35 (4): 325–339. doi:10.1016/S0024-2829(03)00051-3
- ↑ Thüs, Holger; Muggia, Lucia; Pérez-Ortega, Sergio; Favero-Longo, Sergio E.; Joneson, Suzanne; O’Brien, Heath; Nelsen, Matthew P.; Duque-Thüs, Rhinaixa; Grube, Martin; Friedl, Thomas; Brodie, Juliet; Andrew, Carrie J.; Lücking, Robert; Lutzoni, François; Gueidan, Cécile (2011). «Revisiting photobiont diversity in the lichen family Verrucariaceae (Ascomycota)». European Journal of Phycology. 46 (4): 399–415. Bibcode:2011EJPhy..46..399T. doi:10.1080/09670262.2011.629788
- ↑ Pröschold, Thomas; Darienko, Tatyana (2020). «The green puzzle Stichococcus (Trebouxiophyceae, Chlorophyta): new generic and species concept among this widely distributed genus». Phytotaxa. 441 (1): 113–142. doi:10.11646/phytotaxa.441.2.2
- ↑ Veselá, Veronika; Malavasi, Veronica; Škaloud, Pavel (2024). «A synopsis of green-algal lichen symbionts with an emphasis on their free-living lifestyle». Phycologia. 63 (3): 317–338. doi:10.1080/00318884.2024.2325329. hdl:11577/3511125
- ↑ Degelius, Gunnar (1935). Das oceanische Element der Strauch- und Laubflechtenflora von Skandinavien [O elemento oceânico da flora de arbustos e líquens de folhas da Escandinávia]. Col: Acta Phytogeographica Suecica (em alemão). 7. [S.l.: s.n.]
- ↑ Culberson, William Louis; Mason E., Hale (1966). «The range of Normandina pulchella in North America». The Bryologist. 69 (3): 365–367. JSTOR 3240836. doi:10.2307/3240836
- ↑ Spribille, Toby; Fryday, Alan M.; Hampton-Miller, Celia J.; Ahti, Teuvo; Dillman, Karen; Thor, Göran; Tonsberg, Tor; Schirokauer, Dave, eds. (2023). Compendium of the Lichens and Associated Fungi of Alaska. Col: Bibliothecia Lichenologica. [S.l.]: J. Cramer. p. 278. ISBN 978-3-443-58093-3. doi:10.1127/bibl_lich/2023/112
- ↑ Orange, Alan (2022). «The crustose species of Normandina Verrucariaceae)». The Lichenologist. 54 (6): 371–378. doi:10.1017/s0024282922000317
- ↑ Sharnoff, Stephen (2014). A Field Guide to California Lichens. New Haven/London: Yale University Press. p. 357. ISBN 978-0-300-19500-2
- ↑ Hyvärinen, Esko; Juslén, Aino; Kemppainen, Eija; Uddström, Annika; Liukko, Ulla-Maija, eds. (2019). Suomen lajien uhanalaisuus – Punainen kirja 2019. The 2019 Red List of Finnish Species (Relatório). Ministry of the Environment. Finnish Environment Institute. p. 292. ISBN 978-952-11-4974-0
- ↑ Diederich, Paul; Lawrey, James D.; Ertz, Damien (2018). «The 2018 classification and checklist of lichenicolous fungi, with 2000 non-lichenized, obligately lichenicolous taxa». The Bryologist. 121 (3): 385; 392. doi:10.1639/0007-2745-121.3.340
- ↑ Lücking, Robert; Moncada, Bibiana (2017). «Dismantling Marchandiomphalina into Agonimia (Verrucariaceae) and Lawreymyces gen. nov. (Corticiaceae): setting a precedent to the formal recognition of thousands of voucherless fungi based on type sequences». Fungal Diversity. 84 (1): 119–138. doi:10.1007/s13225-017-0382-4