Myxozoa
Myxozoa
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Pré-Cambriano - Recente 600–0 Ma
(baseado em dados moleculares) | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Subclasses | |||||||||||||
| |||||||||||||
Myxozoa (etimologia: gr. μύξα myxa = "muco" + ζῷον zoon "animal") é uma linhagem de Cnidaria altamente modificada, constituída inteiramente por endoparasitas obrigatórios. Os mixozoários são organismos oligocelulares, – isto é, compostos de pouquíssimas células, mesmo em estágios maduros do ciclo de vida. De fato, eles estão entre os mais reduzidos e simplificados metazoários conhecidos.[1] Ademais desta simplificação, estes animais são super diversos, de ocorrência em habitats marinhos e de água doce, com morfologias bastante variáveis ao longo do ciclo de vida.[2] Dentro da grande linhagem de Cnidaria, apenas os mixozoários e os hidrozoários são conhecidos para ambientes de água doce.
Quanto à origem do grupo, análises moleculares apontam para uma cladogênese datada no Pré-Cambriano, há cerca de 600 milhões de anos atrás.[3] Este é um tempo de divergência 100 milhões de anos mais tarde daquele reconstruído para o último ancestral comum cnidário. Esta origem antiquíssima, somada a uma rápida evolução molecular, afeta as análises filogenéticas; gerando efeitos de ramos longos entre os mixozoários e seus parentes de vida livre mais próximos.[4][5]
Atualmente, há pouco mais de 3'000 espécies atualmente conhecidas, compreendendo cerca de 14% de toda a diversidade de cnidários (pouco menos de 21'000).[6] Algumas dessas são capazes de causar doenças economicamente importantes, prejudicando peixes selvagens e de criatórios.
Os mixozoários são agrupados em dois grandes clados: 1 - o primitivo Malacosporea Canning, Curry, Feist, Longshaw & Okamura, 2000, que inclui 6 espécies de malacosporídeos; e 2 - Myxosporea Bütschli, 1881, onde estão as outras 3'000 espécies, chamados mixosporídeos.[6][7] Mais recentemente, a única espécie de polipódio conhecida, Polypodium hydriforme, vem sendo entendida como grupo-irmão dos mixozoários (Polypodizoa). Juntos, o menor clado – aliás inteiramente parasita – que os une recebeu o nome de Endocnidozoa Zrzavý & Hypša, 2003, fazendo referência ao hábito endoparasitário das duas linhagens.[8]
História Taxonômica
A primeira espécie de Myxozoa parece ter sido reportada em 1825, pelo médico suíço Louis Jurine.[9] Contudo, uma descrição inequívoca desses parasitas só pode ser traçada em 1838, quando J. Müller os enquadra num grupo informal, conhecido a época como "Psorospermos".[10] Robins (1853) foi o primeiro a formalizar o grupo como uma tribo de diatomáceas do filo Protozoa.[11] Três décadas mais tarde, uma outra classificação taxonômica formal foi proposta pelo zoólogo alemão Otto Bütschli (1881).[12] Este reconhecera diferenças importantes entre os "mixosporidídios" (termo cunhado para a revisão) e o grupo das diatomáceas psorospermas. Os estágios trofozoítos plasmodiais, o desenvolvimento dos esporos e a descarga dos filamentos polares foram importantes para esta diferenciação.[13] Bütschli (1881) então propõe que os Myxosporea (ordem Myxosporida) pertenceriam ao grupo de esporozoários então reconhecido como natural, Sporozoa Leuckart, 1879. Este grupo, além dos mixozoários, hangariaria também alguns alveolados apicomplexos e a atual linhagem de fungos Microsporidia (a época Mixosporea Doflein, 1901).[10]
.jpg)
Mas isto quanto ao estágio de mixósporo. Quanto aos actinósporos, a infecção destes em oligoquetas aquáticos só foi primeiramente reportada pelo zoólogo tcheco Antonín Štolc (1899).[14] Ele criou o grupo Actinomyxidia, que agregava algumas "espécies" (hoje morfologias de esporos) infectantes de anelídios tubificídios, entre as quais: Hexactinomyxon sp., Synactinomyxon sp. e Tractinomyxon sp.
A primeira história de vida completa de um mixozoário foi elucidada em 1984, para o famigerado parasita Myxobolus cerebralis Hofer, 1903.[15] Este é um parasita conhecido pela transmissão da chamada "doença do redopio", uma enfermidade que acomete jovens peixes salmonídeos. A época, Wolf & Markiw (1983) demonstraram que o desenvolvimento do então actinosporídeo Triactinomyxon sp. ocorria no epitélio intestinal do oligoqueta Tubifex tubifex.[16] Com esta descrição, entendeu-se que há uma forma dupla para os esporos dos mixozoários, cuja infecção é alternada entre actinósporos para os hospedeiros vertebrado e mixósporos para os invertebrado de um mesmo ciclo.[17]
Esta descoberta leva a uma grande revisão taxonômica do grupo, em que Kent et al. (1994) propõem o desuso da classe Actinosporea; com a persistência do nome em vernáculo usado apenas para se referir a um morfotipo de esporo.[18] A ocorrência desse estágio seria num mesmo ciclo de vida que um mixósporo.[19] Desde então, evidências ultraestruturais de características conhecidas dos metazoários, como diferenciação celular terminal, septo intercelular, junções desmossomais, comunicação citoplasmática intercelular e colágeno reticulado foram observadas; confirmando a filiação metazoária dos Myxozoa. Mais recentemente, dados moleculares participam de maneira essencial para desvendar a diversidade e as relações entre as espécies dentro do grupo.[20][21]
Ciclo de Vida
Todos os mixozoários conhecidos são endoparasitas obrigatórios. A maior parte deles exibe histórias de vida complexas, apresentando mais de uma morfologia, e sendo geralmente heteroxênicos – envolvendo a infecção e desenvolvimento em pelo mais de um hospedeiro.[22] Os casos documentados costumam tê-los, como hospedeiro intermediário, um vertebrado – geralmente peixes, – e, como hospedeiro definitivo, algum invertebrado, como anelídeos ou briozoários. Estes são considerados definitivos porque é neles em que ocorre a reprodução sexuada (meiose).[23] Alguns tetrápodes – como anfíbios, lepidossáurios, testudíneos, aves aquáticas e pequenos mamíferos – também já foram observados como hospedeiros intermediários.[24][25][26][27] Não há evidências de que alguma história de vida mixozoária envolva um hospedeiro humano. No entanto, formas espóricas latentes já foram detectadas em fezes humanas.[28]
Não há um padrão muito claro quanto a fidelidade dos parasitas aos hospedeiros.[9] Algumas espécies realizam uma infecção estritamente hospedeiro-específicas, enquanto outras apresentam pouca seletividade, podendo apresentar grande variação e capacidade de aclimatação.[29] De todas as mais de 2,600 espécies conhecidas, apenas cerca de 120 tiveram seus ciclos de vida completamente resolvidos. A ocorrência de mixozoários em tetrápodes já levantou suspeitas de que alguns ciclos de vida possam ser exclusivamente terrestres ou, no mínimo, semiaquáticos.[30] Este ciclo hipotético seria um caso único de conquista do ambiente terrestre por um cnidário.
Estágios de vida
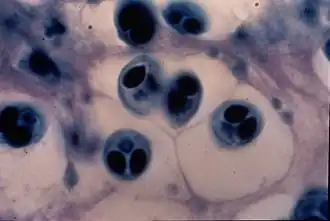
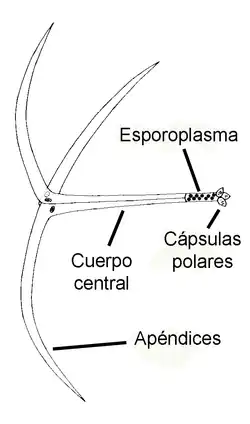
De maneira geral, há dois estágios principais e alternantes num ciclo de vida bifásico mixozoário: um estágio de mixósporo (produzido no vertebrado e infeccioso no invertebrados) e um estágio de actinósporo (produzido no invertebrado e infeccioso no vertebrado).[31] Ambos os estágios são elaborações multicelulares de formas espóricas, – unidades funcionais dispersivas, em geral latentes, e responsáveis pela transmissão do parasita entre os dois tipos de hospedeiros (intermediário e definitivo). Em Myxozoa, ambas as formas partilham das mesmas unidades anatômicas, sendo apenas o arranjo dessas unidades morfologicamente distinto para cada forma e espécie. Todas carregam, no mínimo: 1 - uma ou duas células ameboides infecciosas (denominadas esporoplasmas) e 2 - células portadoras de cápsulas polares (denominadas células capsulogênicas), em quantidades variáveis de um a quinze.
Em geral, além dos dois tipos celulares básicos, pode haver também uma camada protetora de células achatadas, conhecidas como células valvares. Estas células, assim chamadas por encerrarem as outras como as valvas de uma concha, podem secretar revestimentos protetores da superfície, como projeções ou envelopes mucosos, e formar elaborados apêndices de flutuação. Todas essas estruturas das valvas auxiliam na dispersão dos mixósporos pelo ambiente. A quantidade e a forma de cada um desses tipos celulares é um importante caráter taxonômico.[32]
As formas espóricas, latentes, se alternam com formas extrasporogônicas, ativas, também chamadas de trofozoítos. Nesses estágios, há o crescimento somático e germinativo do parasita; que podem ser organizados de diversas maneiras. Nessas formas tróficas, pode-se verificar um aumento no número de núcleos, tanto por meio de mitose clássica quanto por cariocinese endogênica. Esta última caracteriza-se pelo envolvimento de uma célula secundária, isto é, interiorizada, pelo retículo endoplasmático dentro de uma célula parental, neste caso chamada primária. Esta é uma modalidade de cariogênese particularmente comum na linhagem Malacosporea; em que este mesmo processo pode se repetir mesmo dentro das células secundárias. Sucessivamente, portanto, podem originar-se níveis mais altos de proliferação endógena (como terciária ou até mesmo quartenária); os quais, com o rompimento das respectivas células parentais, são liberadas e podem continuam a história de vida do parasita.
As formas extrasporogênicas mais comuns são: 1 - os pseudoplasmódios (sempre uninucleados); 2 - os plasmódios sinciciais (multinucleados, sem membranas encerrando os núcleos individuais); e 3 - os plasmódios estratificados. Todas estas formas são capazes de estender projeções citoplasmáticas temporárias (pseudópodes), que auxiliam a migração do parasita através dos tecidos hospedeiros.
Quanto ao hábito, as formas tróficas podem ser tanto histozoicas – isto é, residirem entre as células dos tecidos do hospedeiro (intercelulares); quanto celozoicas – residindo nas cavidades corpóreas do hospedeiro (primordialmente no trato urinário ou na bexiga natatória). Algumas várias espécies também tiveram hábito citozoico documentado, caracterizando-os como verdadeiros parasitas intracelulares; raríssimos entre os metazoários.
Mixósporo
A forma de mixósporos representa a maior parte da diversidade conhecida em Myxozoa.[9] Este é o estágio produzido dentro do hospedeiro vertebrado. Seu tamanho típico varia entre 10 e 20 μm, com algumas espécies do gênero Ceratomyxa chegando a mais de 740 μm (C. maxima).[33] Quanto a morfologia, nos mixósporos da linhagem Bivalvulida (Myxosporea), há duas valvas; enquanto em Multivalvulida (Myxosporea), há quatro ou mais, que se aderem em conjunto formando uma linha de sutura.[34] O estágio mixósporo da grande linhagem Malacosporea é chamado "malacósporo de peixes" (do inglês: fishmalacospores). Este é caracterizado por duas células valvares envolvendo um esporoplasma, que carrega duas cápsulas polares.[35]

A depender da espécie e de seu hábito parasita, os mixospóros (e malacospóros de peixes) podem se desenvolver em "plasmódios" ou "pseudoplasmódios" ainda no hospedeiro vertebrado, – transmitindo-se assim na forma extraesporogênica aos hospedeiros invertebrados. Alternativamente, mixósporos são ingeridos diretamente pelo hospedeiro invertebrado, – caso em que há o chamado desenvolvimento "intraoligoqueta".[36] Uma vez no intestino deste último, os filamentos das cápsulas polares são extrudidos e ancoram o parasita no epitélio intestinal. A abertura das valvas permitem o esporoplasma penetrar no epitélio histozoica, cenozoica ou intracelularmente, onde o parasita realiza sua reprodução sexuada, formando uma estrutura chamada pansporocisto.[37] O produto da reprodução são dezesseis células gaméticas (células α e β), que formam oito zigotos. Estes oito zigotos são actinósporos diploides, ainda envolvidos por células chamadas pericitos, componentes do pansporocisto.[9]
A manutenção da infecciosidade dos esporos parece ser diferente entre as linhagens de Myxosporea e Malacosporea. Enquanto os últimos demonstraram uma vida útil curta; os primeiros podem continuar infecciosos por períodos variáveis entre meses e anos.[38]
Actinósporo
As formas actinospóricas são caracterizados por uma simetria trirradiada. A maior parte desses esporos contêm três cápsulas polares e três valvas (apêndices), que participam na abertura dos tecidos hospedeiros para a ação dos filamentos das cápsulas. Tipicamente, o esporoplasma já é semelhante a um estágio de plasmódio (com muitos núcleos e células secundárias), e fica localizado posteriormente à estrutura das valvas. Ademais há, atualmente, aproximadamente 20 grandes grupos morfológicos reconhecidos e válidos de actinósporos.[39][40] Desses, o maior dos grupos é o chamado aurantiactinomyxon, com 61 tipos, baseados em morfometria, reportados.[17] Este tipo é caracterizado por três processos valvulares em forma de folha, que se ligam a um esporoplasma com muitas células secundárias (internas).[32]
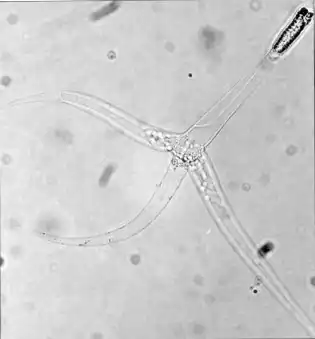
Após a maturação, actinósporos são lançados de seus hospedeiros invertebrados à coluna d'água.[41] A partir do contato com o muco da pele ou das brânquias do vertebrado, seus filamentos das cápsulas são disparados e os esporoplasmas penetram o epitélio hospedeiro.[42] Em contato com a superfície, o esporoplasma inicial um movimento ameboide que, facilitado por atividades proteolíticas, penetra os tecidos afins ao seu desenvolvimento.[43] Em seguida, o esporoplasma se aloja e inicia novamente o desenvolvimento da forma mixospórica, passando pelo estágio de trofozoíto (também chamada de forma pré-esporogênica). Muitas vezes, este trofozoíto, com células interiorizadas e inúmeros núcleos, migra via sistema nervoso e/ou circulatório pelo corpo do hospedeiro.[44] Dessa maneira, a forma pré-esporogênica atinge microambientes propícios à geração da forma esporogênica. O desenvolvimento dos mixósporos é então realizado no corpo do hospedeiro vertebrado, e não envolve a maquinaria meiótica.[37]
Quanto a longevidade, actinósporos na coluna d'água têm duração variável entre 4 a 25 dias, dependendo das condições ambientais (especialmente temperatura) e da espécie.[45][46] Imagina-se que, durante a fase planctônica, a alta velocidade dos fluxos d'água possam causar danos mecânicos ao esporo; além de limitar o tempo de contato entre estes e o hospedeiro vertebrado.[47]
Cápsulas polares
As cápsulas polares são estruturas características dos mixozoários, essenciais para o sucesso do parasitismo desses organismos. Elas possuem uma notável semelhança estrutural e funcional com as cnidas presentes nos cnidários de vida livre, como as águas-vivas, anêmonas e corais. Essa semelhança tem sido fundamental para a compreensão da filogenia dos mixozoários e é um dos principais indícios de que esses parasitas microscópicos seriam, portanto, cnidários altamente modificados.[48]
_Figure_4.png)
As cápsulas polares dos mixozoários compartilham com as cnidas a mesma arquitetura geral: uma cápsula rígida contendo um tubo enrolado que é evertido rapidamente durante o disparo. No entanto, existem diferenças importantes tanto na fisiologia do mecanismo quanto na estrutura do tubo e na finalidade do disparo. A pressão interna das cápsulas é consideravelmente menor, variando entre 50 e 100 atmosferas, e a velocidade de eversão do tubo também é inferior, em torno de 0,5 a 1 m/s. Embora também haja um componente osmótico no processo, há evidências de que proteínas semelhantes ao colágeno desempenham um papel crucial na contração da cápsula, acumulando e liberando repentinamente energia potencial elástica para o disparo. Além disso, o tubo das cápsulas polares não é espiralado como nas cnidas típicas, mas apresenta microespículas pontiagudas em sua extremidade, adaptados para perfurar as membranas das células do hospedeiro e garantir a fixação do parasita no momento da infecção.[48]
Essas diferenças refletem adaptações distintas aos modos de vida de cada grupo. Enquanto as cnidas são utilizadas para predação e defesa nos cnidários de vida livre, as cápsulas polares dos mixozoários são fundamentais para iniciar o ciclo de vida parasitário, funcionando como dispositivos de penetração e fixação. Apesar dessas especializações, a semelhança estrutural entre as cápsulas polares e as cnidas é tão significativa que sugere uma origem evolutiva comum. Essa homologia entre as duas organelas é um dos principais argumentos para sustentar a inclusão dos mixozoários dentro do filo Cnidaria.[48]
Mecanismo de disparo
Nas cnidas, o mecanismo de disparo depende de uma pressão osmótica extremamente elevada, gerada pelo acúmulo de poliglutamato e íons, principalmente potássio, no interior da cápsula. Isso cria uma tensão interna que pode ultrapassar 150 atmosferas. Quando a cápsula é ativada, o tubo que se encontra enrolado em seu interior é evertido de maneira abrupta, perfurando o alvo e liberando toxinas. Esse processo ocorre em frações de milissegundo e pode atingir velocidades de 2 a 3 m/s, sendo uma das reações biológicas mais rápidas conhecidas. O tubo evertido é geralmente espiralado e liso, adequado para penetração e envenenamento de presas ou para defesa.[48]
Taxonomia
A taxonomia dos mixozoários, desde sua descrição, enfrenta mudanças importantes nos níveis genérico, familiar e de subordens. Na última década, o grupo recebia classificações provisórias dentro do clado Cnidaria. Em 2015, houve uma grande revisão do sistema taxonômico baseado em esporos até o nível de gênero:[49]
| Filo: Cnidaria | |
| Subfilo: Myxozoa (não classificado) | |
| Classe: Malacosporea | Classe: Myxosporea (continuação) |
| Ordem: Malacovalvulida | Ordem: Bivalvulida |
| Família: Saccosporidae | Família: Myxobilatidae |
| Gênero: Buddenbrockia, Tetracapsuloides | Gênero: Myxobilatus, Acauda, Hoferellus |
| Classe: Myxosporea | Família: Chloromyxidae |
| Ordem: Bivalvulida | Gênero: Chloromyxum, Caudomyxum, Agarella |
| SubOrdem: Variisporina | Família: Coccomyxidae |
| Família: Sphaeromyxidae | Gênero: Coccomyxa, Auerbachia, Globospora |
| Gênero: Sphaeromyxa | Família: Alatosporidae |
| Família: Myxidiidae | Gênero: Alatospora, Pseudalatospora, Renispora |
| Gênero: Myxidium, Zschokkella, Enteromyxum, Sigmomyxa, Soricimyxum, Cystodiscus | Família: Parvicapsulidae |
| Família: Ortholineidae | Gênero: Parvicapsula, Neoparvicapsula, Gadimyxa |
| Gênero: Ortholinea, Neomyxobolus, Cardimyxobolus, Triangula, Kentmoseria | SubOrdem: Platysporina |
| Família: Sinuolineidae | Família: Myxobolidae |
| Gênero: Sinuolinea, Myxodavisia, Myxoproteus, Bipteria, Paramyxoproteus, Neobipteria, Schulmania, Noblea, Latyspora | Gênero: Myxobolus, Spirosuturia, Unicauda, Dicauda, Phlogospora, Laterocaudata, Henneguya, Hennegoides, Tetrauronema, Thelohanellus, Neothelohanellus, Neohenneguya, Trigonosporus |
| Família: Fabesporidae | Ordem: Multivalvulida |
| Gênero: Fabespora | Família: Trilosporidae |
| Família: Ceratomyxidae | Gênero: Trilospora, Unicapsula |
| Gênero: Ceratomyxa, Meglitschia, Ellipsomyxa, Ceratonova | Família: Kudoidae |
| Família: Sphaerosporidae | Gênero: Kudoa |
| Gênero: Sphaerospora, Wardia, Palliatus | Família: Spinavaculidae |
| Gênero: Octospina | |
| *Incertae sedis em Multivalvulida: Trilosporoides | |
Em 2025, por outro lado, uma nova classificação foi publicada mundialmente pelo World List of Myxozoa; um banco de dados de espécies global sob os domínios do renomado World Register of Marine Species (WoRMS).[50]
Subclasse Malacosporea
- Ordem Malacovalvullida
- Família Saccosporidae Canning, Okamura & Curry, 1996
Subclasse Myxosporea
- Ordem Bivalvulida
- Subordem Platysporina
- Família Myxobilatidae Shulman, 1953
- Família Myxobolidae Thélohan, 1892
- Família Spirosuturidae Chen & Hsieh, 1984
- Família Tetraunematidae Wu, Wang & Jiang, 1988
- Subordem Spaeromyxina
- Família Sphaeromyxidae Lom & Noble, 1984
- Subordem Variisporina
- Família Alatasporidae Schulman, Kovaleva & Dubina, 1979
- Família Ceratomyxidae Doflein, 1899
- Família Chloromyxidae Thélohan, 1892
- Família Coccomyxidae Léger & Hesse, 1907
- Família Enteromyxidae Freeman, Yanagida & Kristmundsson, 2020
- Família Fabesporidae Naidenova & Zaika, 1969
- Família Gadimyxidae Yurakhno, 2011
- Família Myxidiidae Thélohan, 1892
- Família Neothelohanellidae Sarkar & Misra, 1986
- Família Ortholineidae Lom & Noble, 1984
- Família Parvicapsulidae Shulman, 1953
- Família Sinuolineidae Shulman, 1959
- Família Sphaerosporidae Davis, 1917
- Subordem Platysporina
- Ordem Multivalvullida
- Família Kudoidae Meglitsch, 1960
- Família Spinavaculidae Hsieh & Xiao, 1993
- Família Trilosporidae Shulman, 1959
- Multivalvulida incertae sedis
Referências
- ↑ Atkinson, Stephen D.; Bartholomew, Jerri L.; Lotan, Tamar (1 de agosto de 2018). «Myxozoans: Ancient metazoan parasites find a home in phylum Cnidaria». Zoology: 66–68. ISSN 0944-2006. doi:10.1016/j.zool.2018.06.005. Consultado em 24 de julho de 2025
- ↑ Fiala, Ivan; Bartošová-Sojková, Pavla; Okamura, Beth; Hartikainen, Hanna (2015). Okamura, Beth; Gruhl, Alexander; Bartholomew, Jerri L., eds. «Adaptive Radiation and Evolution Within the Myxozoa». Cham: Springer International Publishing (em inglês): 69–84. ISBN 978-3-319-14753-6. Consultado em 24 de julho de 2025
- ↑ Holzer, Astrid S.; Bartošová‐Sojková, Pavla; Born‐Torrijos, Ana; Lövy, Alena; Hartigan, Ashlie; Fiala, Ivan (abril de 2018). «The joint evolution of the Myxozoa and their alternate hosts: A cnidarian recipe for success and vast biodiversity». Molecular Ecology (7): 1651–1666. ISSN 0962-1083. doi:10.1111/mec.14558. Consultado em 24 de julho de 2025
- ↑ Nesnidal, Maximilian P.; Helmkampf, Martin; Bruchhaus, Iris; El-Matbouli, Mansour; Hausdorf, Bernhard (30 de jan. de 2013). «Agent of Whirling Disease Meets Orphan Worm: Phylogenomic Analyses Firmly Place Myxozoa in Cnidaria». PLOS ONE (em inglês) (1): e54576. ISSN 1932-6203. doi:10.1371/journal.pone.0054576. Consultado em 24 de julho de 2025
- ↑ Chang, E. Sally; Neuhof, Moran; Rubinstein, Nimrod D.; Diamant, Arik; Philippe, Hervé; Huchon, Dorothée; Cartwright, Paulyn (dezembro de 2015). «Genomic insights into the evolutionary origin of Myxozoa within Cnidaria». Proceedings of the National Academy of Sciences (48): 14912–14917. doi:10.1073/pnas.1511468112. Consultado em 24 de julho de 2025
- ↑ a b «WoRMS - World Register of Marine Species - Myxozoa». www.marinespecies.org (em inglês). Consultado em 24 de julho de 2025
- ↑ CANNING, ELIZABETH U.; CURRY, ALAN; FEIST, STEPHEN W.; LONGSHAW, MATTHEW; OKAMURA, BETH (setembro de 2000). «A New Class and Order of Myxozoans to Accommodate Parasites of Bryozoans with Ultrastructural Observations on Tetracapsula bryosalmonae (PKX Organism)». Journal of Eukaryotic Microbiology (5): 456–468. ISSN 1066-5234. doi:10.1111/j.1550-7408.2000.tb00075.x. Consultado em 24 de julho de 2025
- ↑ «Myxozoa, Polypodium, and the origin of the Bilateria: The phylogenetic position of "Endocnidozoa" in light of the rediscovery of Buddenbrockia». Cladistics (em inglês) (2): 164–169. 2003. ISSN 1096-0031. doi:10.1111/j.1096-0031.2003.tb00305.x. Consultado em 24 de julho de 2025
- ↑ a b c d Grabner, Daniel; Fiala, Ivan (2025). Smit, Nico J.; Sures, Bernd, eds. «Biology and Life Cycles of Microsporidia and Myxozoa». Cham: Springer Nature Switzerland (em inglês): 41–69. ISBN 978-3-031-83903-0. doi:10.1007/978-3-031-83903-0_3. Consultado em 20 de julho de 2025
- ↑ a b Foox, Jonathan; Siddall, Mark E. (junho de 2015). «The Road To Cnidaria: History of Phylogeny of the Myxozoa». Journal of Parasitology (3): 269–274. ISSN 0022-3395. doi:10.1645/14-671.1. Consultado em 24 de julho de 2025
- ↑ Robin, Ch. (1853). Histoire naturelle des végétaux parasites qui croissent sur l'homme et sur les animaux vivants /. New York :: J.-B. Ballière,. Consultado em 24 de julho de 2025
- ↑ Bütschli, Otto (1881). «Myxosporidien». Zoologischer Jahresbericht für 1880. 1: 162–164
- ↑ Poblete Vidal, Letícia; Iannacone, José; M. Whipps, Christopher; Luque, José Luis (1 de julho de 2017). «SINOPSIS DE LAS ESPECIES DE MYXOZOA GRASSÉ, 1970 (CNIDARIA: MYXOSPOREA) EN LAS AMÉRICAS». Neotropical Helminthology (2): 413–51. ISSN 1995-1043. doi:10.24039/rnh2017112716. Consultado em 22 de julho de 2025
- ↑ Štolc, Antonín (1899). «Actinomyxidies, nouveau groupe de Mesozoaires parent des Myxosporidies.». Bulletin International de l’Académie des Sciences de Bohème (12): 1–12
- ↑ Wolf, Ken; Markiw, Maria E. (28 de setembro de 1984). «Biology Contravenes Taxonomy in the Myxozoa: New Discoveries Show Alternation of Invertebrate and Vertebrate Hosts». Science (4669): 1449–1452. doi:10.1126/science.225.4669.1449. Consultado em 20 de julho de 2025
- ↑ Markiw, Maria E.; Wolf, Ken (1983). «Myxosoma cerebralis (Myxozoa: Myxosporea) Etiologic Agent of Salmonid Whirling Disease Requires Tubificid Worm (Annelida: Oligochaeta) in its Life Cycle». The Journal of Protozoology (em inglês) (3): 561–564. ISSN 1550-7408. doi:10.1111/j.1550-7408.1983.tb01422.x. Consultado em 24 de julho de 2025
- ↑ a b Rocha, Sónia (1 de junho de 2023). «Synopsis of the aurantiactinomyxon collective group (Cnidaria, Myxozoa), with a discussion on the validity of morphotype definition and demise of guyenotia». Systematic Parasitology (em inglês) (3): 307–323. ISSN 1573-5192. PMC 10129985
 . doi:10.1007/s11230-023-10089-1. Consultado em 20 de julho de 2025
. doi:10.1007/s11230-023-10089-1. Consultado em 20 de julho de 2025
- ↑ Kent, M. L.; Margolis, L.; Corliss, J. O. (1 de maio de 1994). «The demise of a class of protists: taxonomic and nomenclatural revisions proposed for the protest phylum Myxozoa Grassé, 1970». Canadian Journal of Zoology (5): 932–937. ISSN 0008-4301. doi:10.1139/z94-126. Consultado em 20 de julho de 2025
- ↑ Yokoyama, H; Ogawa, K; Wakabayashi, H (1993). «Some biological characteristics ol actmosporeans from the oiigochaete Branchiura sowerbyi». Diseases of Aquatic Organisms: 223–228. ISSN 0177-5103. doi:10.3354/dao017223. Consultado em 22 de julho de 2025
- ↑ Kodádková, Alena; Bartošová-Sojková, Pavla; Holzer, Astrid S.; Fiala, Ivan (1 de março de 2015). «Bipteria vetusta n. sp. – an old parasite in an old host: tracing the origin of myxosporean parasitism in vertebrates». International Journal for Parasitology (4): 269–276. ISSN 0020-7519. doi:10.1016/j.ijpara.2014.12.004. Consultado em 20 de julho de 2025
- ↑ Lisnerová, Martina; Blabolil, Petr; Holzer, Astrid; Jurajda, Pavel; Fiala, Ivan (28 de julho de 2020). «Myxozoan hidden diversity: the case of Myxobolus pseudodispar Gorbunova, 1936». Folia Parasitologica (em inglês). doi:10.14411/fp.2020.019. Consultado em 20 de julho de 2025
- ↑ Gruhl, Alexander (2015). Wanninger, Andreas, ed. «Myxozoa». Vienna: Springer (em inglês): 165–177. ISBN 978-3-7091-1862-7. doi:10.1007/978-3-7091-1862-7_7. Consultado em 24 de julho de 2025
- ↑ Holzer, Astrid S.; Bartošová-Sojková, Pavla; Born-Torrijos, Ana; Lövy, Alena; Hartigan, Ashlie; Fiala, Ivan (2018). «The joint evolution of the Myxozoa and their alternate hosts: A cnidarian recipe for success and vast biodiversity». Molecular Ecology (em inglês) (7): 1651–1666. ISSN 1365-294X. doi:10.1111/mec.14558. Consultado em 20 de julho de 2025
- ↑ Eiras, Jorge C. (2005). «An overview on the myxosporean parasites in amphibians and reptiles» (PDF). Acta Parasitologica. 50 (4): 267–275. ISSN 1230-2821
- ↑ Prunescu, Carol-Constantin; Prunescu, Paula; Pucek, Zdzislaw; Lom, Jiří (1 de setembro de 2007). «The first finding of myxosporean development from plasmodia to spores in terrestrial mammals: Soricimyxum fegati gen. et sp. n. (Myxozoa) from Sorex araneus (Soricomorpha)». Folia Parasitologica (3): 159–164. doi:10.14411/fp.2007.022. Consultado em 19 de julho de 2025
- ↑ Bartholomew, Jerri L.; Atkinson, Stephen D.; Hallett, Sascha L.; Lowenstine, Linda J.; Garner, Michael M.; Gardiner, Chris H.; Rideout, Bruce A.; Keel, M. Kevin; Brown, Justin D. (agosto de 2008). «Myxozoan parasitism in waterfowl». International Journal for Parasitology (em inglês) (10): 1199–1207. doi:10.1016/j.ijpara.2008.01.008. Consultado em 19 de julho de 2025
- ↑ Szekely, Csaba; Cech, Gabor; Atkinson, Stephen D.; Molnar, Kalman; Egyed, Laszlo; Gubanyi, Andras (19 de agosto de 2015). «A novel myxozoan parasite of terrestrial mammals: description of Soricimyxum minuti sp. n. (Myxosporea) in pygmy shrew Sorex minutus from Hungary». Folia Parasitologica. doi:10.14411/fp.2015.045. Consultado em 19 de julho de 2025
- ↑ REIS, Lisiane Lappe dos; JESUS, Lucas Carvalho de; FERNANDES, Ormezinda Celeste Christo; BARROSO, David Eduardo (abril de 2019). «First report of Myxobolus (Cnidaria: Myxozoa) spores in human feces in Brazil». Acta Amazonica (2): 162–165. ISSN 1809-4392. doi:10.1590/1809-4392201802671. Consultado em 23 de julho de 2025
- ↑ Feist, S. W.; Longshaw, M. (janeiro de 2006). «Phylum Myxozoa.». UK: CABI: 230–296. ISBN 978-0-85199-015-6. Consultado em 19 de julho de 2025
- ↑ Hallett, Sascha L.; Atkinson, Stephen D.; Bartholomew, Jerri L.; Székely, Csaba (2015). Okamura, Beth; Gruhl, Alexander; Bartholomew, Jerri L., eds. «Myxozoans Exploiting Homeotherms». Cham: Springer International Publishing (em inglês): 125–135. ISBN 978-3-319-14752-9. doi:10.1007/978-3-319-14753-6_7. Consultado em 19 de julho de 2025
- ↑ Rocha, S.; Alves, Â; Antunes, C.; Azevedo, C.; Casal, G. (outubro de 2019). «Molecular data infers the involvement of a marine aurantiactinomyxon in the life cycle of the myxosporean parasite Paramyxidium giardi (Cnidaria, Myxozoa)». Parasitology (em inglês) (12): 1555–1563. ISSN 0031-1820. doi:10.1017/S0031182019000866. Consultado em 20 de julho de 2025
- ↑ a b Lom, Jiří; Dyková, Iva (1 de março de 2006). «Myxozoan genera: definition and notes on taxonomy, life-cycle terminology and pathogenic species». Folia Parasitologica (1): 1–36. ISSN 0015-5683. doi:10.14411/fp.2006.001. Consultado em 20 de julho de 2025
- ↑ Eiras, Jorge C.; Cruz, Cristina; Saraiva, Aurélia (1 de junho de 2018). «Synopsis of the species of Ceratomyxa Thélohan, 1892 (Cnidaria, Myxosporea, Ceratomyxidae) described between 2007 and 2017». Systematic Parasitology (em inglês) (5): 427–446. ISSN 1573-5192. doi:10.1007/s11230-018-9791-3. Consultado em 20 de julho de 2025
- ↑ Whipps, C. M.; Grossel, G.; Adlard, R. D.; Yokoyama, H.; Bryant, M. S.; Munday, B. L.; Kent, M. L. (junho de 2004). «PHYLOGENY OF THE MULTIVALVULIDAE (MYXOZOA: MYXOSPOREA) BASED ON COMPARATIVE RIBOSOMAL DNA SEQUENCE ANALYSIS». Journal of Parasitology (em inglês) (3): 618–622. ISSN 0022-3395. doi:10.1645/GE-153R. Consultado em 23 de julho de 2025
- ↑ Morris, D. J.; Adams, A. (agosto de 2008). «Sporogony of Tetracapsuloides bryosalmonae in the brown trout Salmo trutta and the role of the tertiary cell during the vertebrate phase of myxozoan life cycles». Parasitology (em inglês) (9): 1075–1092. ISSN 1469-8161. doi:10.1017/S0031182008004605. Consultado em 20 de julho de 2025
- ↑ Eszterbauer, Edit; Atkinson, Stephen; Diamant, Arik; Morris, David; El-Matbouli, Mansour; Hartikainen, Hanna (2015). Okamura, Beth; Gruhl, Alexander; Bartholomew, Jerri L., eds. «Myxozoan Life Cycles: Practical Approaches and Insights». Cham: Springer International Publishing (em inglês): 175–198. ISBN 978-3-319-14752-9. doi:10.1007/978-3-319-14753-6_10. Verifique
|doi=(ajuda). Consultado em 23 de julho de 2025 - ↑ a b Yokoyama, Hiroshi; Grabner, Daniel; Shirakashi, Sho (11 de abril de 2012). «Transmission Biology of the Myxozoa». InTech. ISBN 978-953-51-0497-1. Consultado em 23 de julho de 2025
- ↑ De Kinkelin, P; Gay, M; Forman, S (2002). «The persistence of infectivity of Tetracapsulabryosalmonae-infected water for rainbow trout, Oncorhynchus mykiss (Walbaum)». Journal of Fish Diseases (em inglês) (8): 477–482. ISSN 1365-2761. doi:10.1046/j.1365-2761.2002.00382.x. Consultado em 20 de julho de 2025
- ↑ Rocha, S; Alves, â; Fernandes, P; Antunes, C; Azevedo, C; Casal, G (5 de setembro de 2019). «New actinosporean description prompts union of the raabeia and echinactinomyxon collective groups (Cnidaria, Myxozoa)». Diseases of Aquatic Organisms (em inglês) (3): 175–191. ISSN 0177-5103. doi:10.3354/dao03389. Consultado em 20 de julho de 2025
- ↑ Rocha, Sónia; Alves, Ângela; Antunes, Carlos; Fernandes, Pedro; Azevedo, Carlos; Casal, Graça (20 de fevereiro de 2020). «Characterisation of sphaeractinomyxon types (Cnidaria: Myxozoa) from marine and freshwater oligochaetes in a Portuguese estuary, with the demise of the endocapsa collective group». Folia Parasitologica (em inglês). doi:10.14411/fp.2020.002. Consultado em 20 de julho de 2025
- ↑ El-Matbouli, M.; Hoffmann, R.W. (janeiro de 1998). «Light and electron microscopic studies on the chronological development of Myxobolus cerebralis to the actinosporean stage in Tubifex tubifex». International Journal for Parasitology (1): 195–217. ISSN 0020-7519. doi:10.1016/s0020-7519(97)00176-8. Consultado em 23 de julho de 2025
- ↑ Xiao, C.; Desser, S. S. (fevereiro de 2000). «The Longevity of Actinosporean Spores from Oligochaetes of Lake Sasajewun, Algonquin Park, Ontario, and Their Reaction to Fish Mucus». The Journal of Parasitology (1). 193 páginas. ISSN 0022-3395. doi:10.2307/3284940. Consultado em 23 de julho de 2025
- ↑ Kelley, GO; Zagmutt-Vergara, FJ; Leutenegger, CM; Adkison, MA; Baxa, DV; Hedrick, RP (2004). «Identification of a serine protease gene expressed by Myxobolus cerebralis during development in rainbow trout Oncorhynchus mykiss». Diseases of Aquatic Organisms: 235–248. ISSN 0177-5103. doi:10.3354/dao059235. Consultado em 23 de julho de 2025
- ↑ Grossel, G W; Dykova, I; Handlinger, J; Munday, B L (junho de 2003). «Pentacapsula neurophila sp.n. (Multivalvulida) from the central nervous system of striped trumpeter, Latris lineata (Forster)». Journal of Fish Diseases (6): 315–320. ISSN 0140-7775. doi:10.1046/j.1365-2761.2003.00459.x. Consultado em 23 de julho de 2025
- ↑ Markiw, Maria E. (janeiro de 1992). <0044:eiwdid>2.3.co;2 «Experimentally Induced Whirling Disease II. Determination of Longevity of the Infective Triactinomyxon Stage ofMyxobolus cerebralisby Vital Staining». Journal of Aquatic Animal Health (1): 44–47. ISSN 0899-7659. doi:10.1577/1548-8667(1992)004<0044:eiwdid>2.3.co;2. Consultado em 23 de julho de 2025
- ↑ Yokoyama, Hiroshi; Ogawa, Kazuo; Wakabayashi, Hisatsugu (1991). «A New Collection Method of Actinosporeans-A Probable Infective Stage of Myxosporeans to Fishes-from Tubificids and Experimental Infection of Goldfish with the Actinosporean, Raabeia sp.». Fish Pathology (3): 133–138. ISSN 0388-788X. doi:10.3147/jsfp.26.133. Consultado em 23 de julho de 2025
- ↑ HALLETT, S. L.; BARTHOLOMEW, J. L. (6 de dezembro de 2007). «Effects of water flow on the infection dynamics ofMyxobolus cerebralis». Parasitology (3): 371–384. ISSN 0031-1820. doi:10.1017/s0031182007003976. Consultado em 23 de julho de 2025
- ↑ a b c d Cannon, Quinn; Wagner, Eric (julho de 2003). «Comparison of Discharge Mechanisms of Cnidarian Cnidae and Myxozoan Polar Capsules». Reviews in Fisheries Science (em inglês) (3): 185–219. ISSN 1064-1262. doi:10.1080/10641260390244305. Consultado em 23 de julho de 2025
- ↑ Fiala, Ivan; Bartošová-Sojková, Pavla; Whipps, Christopher M. (2015). «Classification and Phylogenetics of Myxozoa». In: Beth Okamura, Alexander Gruhl and Jerri L. Bartholomew. Myxozoan Evolution, Ecology and Development (em inglês). [S.l.]: Springer International Publishing. p. 85-110. 455 páginas. ISBN 978-3-319-14752-9. doi:10.1007/978-3-319-14753-6_5
- ↑ «World List of Myxozoa - browser». www.marinespecies.org. Consultado em 31 de outubro de 2025