Lecideaceae
Lecideaceae
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Classificação científica | |||||||||||||
| |||||||||||||
| Género-tipo | |||||||||||||
| Lecidea [en] Ach. (1803) | |||||||||||||
| Sinónimos[1] | |||||||||||||
_Ach_271638.jpg)
Lecideaceae[1] é uma família de fungos formadores de líquenes pertencente à ordem Lecideales [en]. A família abrange cerca de 30 gêneros e aproximadamente 250 espécies. Uma característica distintiva é a forma lecideína dos corpos frutíferos: geralmente circulares, escuros e sem uma margem talina. A maioria das espécies da família é liquenizada com algas verdes, embora algumas, distribuídas em diversos gêneros, sejam liquenícolas, vivendo sobre outros líquenes. Os líquenes da Lecideaceae tendem a crescer em rochas, madeira e solo. Algumas espécies aceleram a meteorização de superfícies rochosas, um processo conhecido como pedogênese, ao estender suas hifas em fissuras e expelir fragmentos de rocha. Isso resulta em taxas de meteorização significativamente mais rápidas em certos ambientes, afetando desde rochas naturais até telhas japonesas Sekishu, e envolve biomoléculas essenciais para sobrevivência e biodeterioração, incluindo compostos que resistem à intensa radiação ultravioleta.
O maior gênero da família, Lecidea [en], já foi um táxon da lixeira que reunia centenas de espécies morfologicamente semelhantes com talos crustosos, margens apoteciais sem fotobiontes e ascósporos unicelulares translúcidos. A taxonomia e classificação dentro da família foram refinadas com estudos recentes de filogenética molecular. Duas espécies da Lecideaceae foram avaliadas para a Lista Vermelha da IUCN global.
Sistemática
Taxonomia histórica
O primeiro membro da atual Lecideaceae a ser formalmente descrito foi Lichen fusco-ater, posteriormente conhecido como Lecidea fuscoatra.[2] O liquenólogo sueco Erik Acharius propôs o gênero Lecidea em 1803, com Lecidea fuscoatra como espécie-tipo.[3] Esse foi um dos vários líquenes descritos pelo taxonomista sueco Lineu em sua obra influente de 1753, Species Plantarum.[4] A família Lecideaceae foi proposta pelo botânico francês François Fulgis Chevallier em 1826, em sua obra Flore générale des environs de Paris; a grafia original do nome era Lecideae. A breve descrição de Chevallier destacava características da forma e textura das estruturas reprodutivas e da crosta. Ele observou que os apotécios (corpos frutíferos) eram inicialmente côncavos, evoluindo para formas planas ou convexas, semelhantes a pequenos pratos ou patelas, cada um delimitado por uma margem distinta. Essa margem pode parecer integrada à crosta, que desvanece com o envelhecimento. Chevallier descreveu a crosta como membranosa, variando de lisa a rachada, e, em alguns casos, espalhando-se de maneira pulverulenta.[5]
Em 1984, Josef Hafellner, em sua obra Studien in Richtung einer natürlicheren Gliederung der Sammelfamilien Lecanoraceae und Lecideaceae,[6] utilizou a estrutura do asco como um principal característica sistemática, dividindo essas duas grandes famílias em várias famílias menores. No entanto, as famílias propostas por ele (Koerberiellaceae, Lecidomataceae, Mycobilimbiaceae e Porpidiaceae) foram posteriormente incorporadas à Lecideaceae;[1] estudos posteriores mostraram que a estrutura do asco não é uma característica taxonômica consistente. Por exemplo, Buschbom e Mueller demonstraram em 2004 que a família Porpidiaceae não era monofilética a menos que a família Lecideaceae também fosse incluída, e que os tipos de asco usados para diferenciá-las eram modificações do mesmo tipo básico.[7] Essa descoberta foi corroborada em 2006 por Miadłikowska e colegas, que sugeriram a reclassificação da família da ordem Lecanorales para uma posição incerta (incertae sedis) na subclasse Lecanoromycetidae.[8] Trabalhos iniciais de filogenética molecular indicaram que a família era monofilética.[9][8]
Classificação
A classificação convencional de fungos produtores de líquenes enfrentou desafios devido à dependência de características observáveis para definir grupos taxonômicos, o que frequentemente resultava em agrupamentos artificiais. Um exemplo disso é o gênero Lecidea, que, na década de 1930, tornou-se um dos maiores gêneros de líquenes, com cerca de 1200 espécies. Esse agrupamento polifilético de espécies semelhantes foi amplificado pelo trabalho de Alexander Zahlbruckner [en], Catalogus lichenum universalis, que incluía qualquer espécie com talo crustoso, apotécios biatorinos ou lecideínos e ascósporos simples nesse táxon da lixeira.[10] Contudo, estudos utilizando caracteres morfológicos e químicos demonstraram que Lecidea, no sentido de Zahlbruckner, era polifilético.[11] No final do século XX, os pesquisadores tinham uma melhor compreensão dos limites do gênero, e muitos táxons foram transferidos para gêneros novos ou já existentes. Até 2011, mais de 160 gêneros de várias famílias incluíam espécies anteriormente classificadas em Lecidea.[10] Na edição de 2008 do Dictionary of the Fungi, estimava-se que Lecidea continha 427 espécies, embora apenas cerca de 100 fossem qualificadas como Lecidea no sentido estrito (sensu stricto).[12] Nesse caso, sensu stricto no sentido de Hertel refere-se a líquenes saxícolas com certos caracteres anatômicos, como excípulo, paráfises e estruturas apicais do asco.[10]
A ordem Lecideales foi proposta por Edvard August Vainio [en] em 1934, no quarto volume de sua obra Lichenographica Fennica.[13] A ordem foi amplamente ignorada em classificações posteriores, pois a família era tradicionalmente colocada em Lecanorales. Em 2011, Schmull e colegas ressuscitaram a ordem, redefinindo o gênero-tipo para incluir apenas Lecidea sensu stricto. Eles usaram filogenética molecular para demonstrar que esse grupo de espécies, definido por morfologia e incluindo a espécie-tipo (Lecidea fuscoatra) e algumas espécies de Porpidia [en], formava um clado monofilético.[10] Análises filogenéticas moleculares mostram a ordem Lecideales como um grupo irmão da Peltigerales.[14]
Etimologia
Seguindo a prática padrão do Código Internacional de Nomenclatura Botânica,[15] o nome Lecideaceae é baseado no nome do gênero-tipo, Lecidea, com o sufixo -aceae indicando o nível taxonômico de família. O nome do gênero deriva da palavra grega antiga λέκος (lékos), que significa "prato" ou "pequeno escudo", e do sufixo "-ídes", indicando semelhança. Isso faz alusão aos apotécios lecideínos, geralmente circulares e sem margem talina.[16]
Descrição
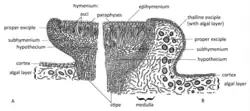
A família Lecideaceae inclui líquenes com formas de crescimento variando de crustoso a escamoso.[1] Em raros casos, o talo pode estar ausente.[17] Esses organismos estabelecem uma relação simbiótica principalmente com algas verdes, e, em alguns casos, como no gênero Amygdalaria [en], também com cianobactérias em estruturas especializadas chamadas cefalódios. As estruturas reprodutivas (ascocarpos) são geralmente apotécios, que podem ser proeminentes na superfície (sésseis) ou parcialmente embutidos (imersos) no talo. Esses apotécios podem assemelhar-se aos dos gêneros Lecidea e Aspicilia [en] (lecideínos ou aspicilioides).[1] A maioria dos gêneros de Lecideaceae possui apotécios lecideínos; exceções incluem Bellemerea, Koerberiella e Lecaimmeria, que têm apotécios lecanorinos.[18] A estrutura e posição do ascocarpo em Cyclohymenia epilithica são únicas em Lecideaceae: esse líquen apresenta uma coluna estéril central cercada por um himênio em forma de anel.[19]
.jpg)
A estrutura interna (hamatécio) dessas estruturas reprodutivas é composta por paráfises pouco ramificadas e interconectadas. Essas estruturas são amiloides, corando de azul com iodo.[1] As paráfises geralmente têm ápices engrossados e frequentemente pigmentados.[17] Os ascos, células portadoras de esporos, são parcialmente fendidos e apresentam uma estrutura amiloide na ponta e paredes amiloides, com uma área central pálida e uma região apical mais escura ou em forma de anel. Os ascos são clavados a cilíndricos.[1]
Os líquenes de Lecideaceae geralmente produzem oito esporos por asco. Esses ascósporos são não septados, de forma cilíndrica a elipsoide, hialinos e não amiloides. Os conidiomas de Lecideaceae são na forma de picnídios;[1] essas estruturas tendem a ter paredes escuras e estar imersas no talo.[17] Os conídios são não septados e podem ser cilíndricos, em forma de bastonete ou filiformes.[1]
Identificar espécies no maior gênero de Lecideaceae, Lecidea, é desafiador devido às semelhanças em morfologia, estruturas anatômicas e composições químicas com outros gêneros, especialmente Lecidella [en] na família Lecanoraceae [en] e Porpidia na família Lecideaceae. As principais distinções entre Lecidea e Lecidella incluem o talo geralmente cinza e granular de Lecidella, com parte inferior preta, azul-preta ou cinza-branca; estruturas reprodutivas como sorédios, isídios e blastídios; presença comum de conidiomas; ascos do tipo Lecanora; paráfises não fundidas e facilmente dispersas; e metabólitos secundários como xantonas, depsidonas de orcinol, depsídeos de β-orcinol e triterpenoides. Lecidea e Porpidia, ambos de Lecideaceae, diferem pelo fato de Porpidia possuir sorédios, isídios e blastídios; conidiomas; ápice de ascos do tipo Porpidia; esporos com halo; paráfises fundidas e ramificadas; e metabólitos secundários como ácido confluêntico, ácido norstíctico, ácido hipostíctico, ácido 2'-O-metilmicrofilínico e ácido 2'-O-metilperlático. Devido às diferenças morfológicas sutis entre esses gêneros, a identificação baseada apenas em morfologia e componentes químicos é difícil.[20]
Fotobiontes
Em um estudo de 1971, Margalith Galun [en] e colegas examinaram a relação micobionte-fotobionte em três espécies de Lecidea (Lecidea olivacea, Lecidea opaca e Lecidea decipiens) usando microscopia eletrônica. Os fotobiontes das duas primeiras são Trebouxia [en], enquanto o da terceira é Myrmecia [en]. Eles observaram que o contato entre os simbiontes variava de invasão fúngica intracelular em talos primitivamente organizados a uma associação mais frouxa de contato em formas de crescimento mais diferenciadas.[21] Trebouxia arboricola [en] foi identificado como um fotobionte comum em Lecideaceae.[22]
Química
Metabólitos secundários (produtos liquênicos) encontrados em líquenes de Lecideaceae incluem depsídeos e depsidonas.[1] O gênero principal, Lecidea, produz diversos metabólitos secundários: ácido confluêntico [en], ácido norstíctico [en], ácido hipostíctico, ácido girofórico [en], 2'-O-metilanziáico, 2'-O-metilmicrofilínico, 2'-O-metilperlático e 4'-O-desmetilplanáico.[20]
No líquen Lecidea lactea, a depsidona ácido norstíctico está envolvida na imobilização de cobre (Cu), formando um composto de coordenação Cu2+-ácido norstíctico. Similarmente, o ácido psorômico, quimicamente semelhante ao ácido norstíctico, é sugerido como quelante de Cu2+ em Lecidea bullata. Essas espécies pertencem à aliança Lecideion inops, uma comunidade de espécies de líquenes adaptadas a sobreviver em substratos ricos em cobre, podendo exibir uma coloração verde distinta em partes do talo ou apotécios devido a depósitos de cobre.[23] Uma investigação subsequente identificou cristais de moolooíta, um composto de oxalato de cobre, em Lecidea inops crescendo sobre minério de calcopirita. Essa descoberta confirma a capacidade do líquen de biomineralizar esse composto em condições ambientais adequadas.[24]
Gêneros



_Hertel_%2526_A.J._Schwab_259698.jpg)

.jpg)
.jpg)

Na sétima edição (2001) do Dictionary of the Fungi, estimava-se que Lecideaceae continha 9 gêneros e 441 espécies;[25] na edição seguinte (2008), esses números foram ajustados para 7 gêneros e 436 espécies. Nesse último trabalho, o maior gênero, Lecidea, foi estimado em cerca de 427 espécies, embora apenas cerca de 100 fossem qualificadas como Lecidea sensu stricto.[12] Segundo uma estimativa de 2022, Lecideaceae compreende 29 gêneros e cerca de 260 espécies.[26] Até fevereiro de 2024, o Index Fungorum (no Catalogue of Life) aceitava 29 gêneros e 244 espécies na família Lecideaceae.[27] Muitos desses gêneros são monoespecíficos ou pequenos, com menos de cinco espécies.[18] A lista a seguir indica o nome do gênero, a autoridade taxonômica, o ano de publicação e o número de espécies:[26]
- Amygdalaria [en] Norman (1852)[28] – 11 espécies
- Bahianora [en] Kalb (1984)[29] – 1 espécie
- Bellemerea [en] Hafellner & Cl.Roux (1984)[30] – 10 espécies
- Bryobilimbia [en] Fryday, Printzen & S.Ekman (2014)[31] – 6 espécies
- Catarrhospora [en] Brusse (1994)[32] – 2 espécies
- Cecidonia [en] Triebel & Rambold (1988)[33] – 2 espécies
- Clauzadea [en] Hafellner & Bellem. (1984)[6] – 7 espécies
- Cyclohymenia [en] McCune & M.J.Curtis (2017)[19] – 1 espécie
- Farnoldia [en] Hertel (1983)[34] – 6 espécies
- Immersaria [en] Rambold & Pietschm. (1989)[35] – 8 espécies
- Imsharria [en] Fryday & U.Rupr. (2024)[36] – 1 espécie
- Koerberiella [en] Stein (1879) – 2 espécies
- Labyrintha [en] Malcolm, Elix & Owe-Larss. (1995)[37] – 1 espécie
- Lecaimmeria [en] C.M.Xie, Lu L.Zhang & Li S.Wang (2022)[18] – 11 espécies
- Lecidea [en] Ach. (1803)[3] – 100 espécies
- Lecidoma [en] Gotth.Schneid. & Hertel (1981)[38] – 1 espécie
- Lopacidia [en] Kalb (1984)[29] – 1 espécie
- Melanolecia [en] Hertel (1981)[39] – 7 espécies
- Pachyphysis [en] R.C.Harris & Ladd (2007)[40] – 1 espécie
- Paraporpidia [en] Rambold & Pietschm. (1989)[35] – 3 espécies
- Poeltiaria [en] Hertel (1984)[41] – 8 espécies
- Poeltidea [en] Hertel & Hafellner (1984)[41] – 3 espécies
- Porpidia [en] Körb. (1855)[42] – 51 espécies
- Porpidinia [en] Timdal (2010)[43] – 2 espécies
- Pseudopannaria [en] (B.de Lesd.) Zahlbr. (1924)[44] – 1 espécie
- Rhizolecia [en] Hertel (1984)[41] – 1 espécie
- Romjularia [en] Timdal (2007)[45] – 1 espécie
- Schizodiscus [en] Brusse (1988)[46] – 1 espécie
- Stenhammarella [en] Hertel (1967)[47] – 1 espécie
- Stephanocyclos [en] Hertel (1983)[48] – 1 espécie
- Xenolecia [en] Hertel (1984)[41] – 2 espécies
Em 2014, Alan Fryday e Hannes Hertel [en] propuseram reduzir os gêneros Labyrintha e Notolecidea a sinônimos de Poeltidea e Poeltiaria, respectivamente.[49]
Diversos gêneros historicamente classificados na família Lecideaceae foram realocados para outras famílias com base em estudos de filogenética molecular. Exemplos incluem Bacidia [en], que foi demonstrado pertencer à Ramalinaceae [en], uma família em Lecanorales.[50] Heppsora [en] foi inicialmente proposto para inclusão em Lecideaceae com base em sua semelhança com Heppia [en] e Psora [en],[51] mas agora também está em Ramalinaceae.[26] Mycobilimbia [en] é outro exemplo de gênero que, até recentemente, era colocado em Lecideaceae[12] mas agora está em Ramalinaceae.[26] Em 2013, o gênero Hypocenomyce [en], residente de Lecideaceae por décadas, foi mostrado como extremamente polifilético e dividido em quatro gêneros distribuídos entre três famílias, nenhuma das quais era Lecideaceae.[52] Roccellinastrum [en] foi incluído na família após uma emenda em 1983, mas agora está na Pilocarpaceae [en].[26]
Habitat, distribuição e ecologia
Os líquenes de Lecideaceae geralmente crescem em cascas, solo e rochas. Coletivamente, a família tem uma distribuição cosmopolita.[1] É uma das famílias mais comuns em comunidades criptoendolíticas da Antártida.[53] Diferentemente dos habitats típicos preferidos por outras espécies norte-americanas de Lecideaceae, Cyclohymenia epilithica prospera exclusivamente em ambientes sombreados em climas temperados frescos e úmidos.[19] A família foi menos estudada no Hemisfério Sul em comparação com o Hemisfério Norte.[49]
Existem 17 espécies de fungos liquenícolas (que vivem sobre líquenes) e líquenes em Lecideaceae. Essas espécies estão distribuídas entre os gêneros Bellemerea, Cecidonia, Immersaria, Lecidea e Poeltiaria. Em alguns casos de líquenes liquenícolas, como Bellemerea cupreoatra e Poeltiaria coromandelica, o líquen juvenil é facultativamente liquenícola, mas torna-se independente na fase adulta.[54]
Pedogênese
.jpg)
Várias espécies de Lecidea contribuem para os processos de meteorização em superfícies rochosas, conhecidos como pedogênese. Essas espécies saxícolas estendem suas hifas em fissuras rochosas, gradualmente desprendendo, incorporando e expelindo fragmentos de rocha. Lecidea auriculata, um exemplo de líquen euendolítico, penetra ativamente e habita a matriz mineral dentro das rochas. Em ambientes alpinos árticos específicos, superfícies colonizadas por esses líquenes meteorizam em taxas estimadas de 25 a 50 vezes mais rápidas do que as causadas por outros processos naturais.[55] O impacto da meteorização de Lecidea auriculata na Pequena Idade do Gelo da geleira Storbreen [en] em Jotunheimen, no sul da Noruega, foi documentado. A atividade degradativa da espécie é limitada por sua faixa ecológica, predominantemente restrita a áreas com cobertura mínima de neve.[56] Espécies de Lecidea também foram observadas degradando uma variedade de materiais, incluindo granito,[57] quartzito de Magaliesberg [en],[58] rochas ultramáficas de serpentinito,[59] e andesito vulcânico.[60] Pesquisas sobre Lecidea tessellata, encontrada em rochas desérticas no oeste da América do Norte, identificaram biomoléculas como oxalato de cálcio monoidratado, o protetor UV citonemina [en] e hematita para resistir a altos níveis de radiação UV, utilizando espectroscopia Raman.[61] Além de materiais naturais, espécies de Lecidea afetam objetos artificiais. Telhas Sekishu são componentes tradicionais de habitações japonesas, cobertas com um esmalte opaco vermelho-acastanhado composto por um vidro amorfo de feldspato alcalino, uma superfície improvável de ser afetada pelo intemperismo químico. Essas telhas são colonizadas por Lecidea, que, após cerca de 7 a 10 anos, resultam no aparecimento de pontos de corrosão de até 50 μm de profundidade.[62][63] Embora a pesquisa sobre pedogênese frequentemente se concentre em Lecidea na família Lecideaceae, efeitos semelhantes de meteorização foram documentados no gênero Porpidia.[64][65]
Conservação
Duas espécies da Lecideaceae foram avaliadas para a Lista Vermelha da IUCN global: Lecidea mayeri (deficiente de dados, 2023)[66] e Immersaria fuliginosa (vulnerável, 2020).[67] Lecidea mayeri é classificada como espécie deficiente de dados devido à falta de informações sobre sua população, habitat, ecologia e ameaças potenciais. Ela é conhecida apenas no município de Angelópolis no departamento de Antioquia, na Colômbia. Mais informações são conhecidas sobre Immersaria fuliginosa, encontrada em duas localidades nas Ilhas Malvinas. Ela enfrenta várias ameaças que podem levar rapidamente ao seu declínio e possível extinção, principalmente devido à sua distribuição muito limitada em apenas duas áreas com uma área total de ocupação de 8 km². As principais ameaças incluem pisoteamento por gado, incêndios (naturais e antrópicos) e os efeitos adversos das mudanças climáticas, como redução das chuvas no verão e aumento da luz solar, que podem impactar negativamente sua sobrevivência. Esforços de conservação são necessários, incluindo proteção de terras e educação local, para mitigar essas ameaças e proteger a espécie.[67]
Referências
- ↑ a b c d e f g h i j k Jaklitsch, Walter; Baral, Hans-Otto; Lücking, Robert; Lumbsch, H. Thorsten (2016). Frey, Wolfgang, ed. Syllabus of Plant Families: Adolf Engler's Syllabus der Pflanzenfamilien. 1/2 13 ed. Berlin Stuttgart: Gebr. Borntraeger Verlagsbuchhandlung, Borntraeger Science Publishers. pp. 129–130. ISBN 978-3-443-01089-8. OCLC 429208213
- ↑ Linnaeus, Carl (1753). Species plantarum (em latim). 2. Stockholm: Impensis Laurentii Salvii. p. 1140
- ↑ a b Acharius, E. (1803). Methodus qua Omnes Detectos Lichenes Secundum Organa Carpomorpha ad Genera, Species et Varietates Redigere atque Observationibus Illustrare Tentavit Erik Acharius (em latim). Stockholm: impensis F.D.D. Ulrich, typis C.F. Marquard. p. 32
- ↑ Jørgensen, Per M. (1994). «Linnaean lichen names and their typification». Botanical Journal of the Linnean Society. 115 (4): 261–405. doi:10.1111/j.1095-8339.1994.tb01784.x
- ↑ Chevallier, François-Fulgis (1826). Flore Générale des Environs de Paris (em francês). 1. Paris: imprimerie de Decourchant rue d'Erfurth nº. 1. p. 549
- ↑ a b Hafellner, J. (1984). «Studien in Richtung einer natürlichen Gliederung der Sammelfamilien Lecanoracae und Lecideaceae» [Studies towards a natural classification of the collective families Lecanoraceae and Lecideaceae]. Beihefte zur Nova Hedwigia (em alemão). 79: 241–371
- ↑ Buschbom, Jutta; Mueller, Gregory (2004). «Resolving evolutionary relationships in the lichen-forming genus Porpidia and related allies (Porpidiaceae, Ascomycota)». Molecular Phylogenetics and Evolution. 32 (1): 66–82. PMID 15186798. doi:10.1016/j.ympev.2003.11.012
- ↑ a b Miadlikowska, J.; Kauff, F.; Hofstetter, V.; Fraker, E.; Grube, M.; Hafellner, J.; Reeb, V.; Hodkinson, B.P.; Kukwa, M.; Lucking, R.; Hestmark, G.; Otalora, M.G.; Rauhut, A.; Budel, B.; Scheidegger, C.; Timdal, E.; Stenroos, S.; Brodo, I.; Perlmutter, G.B.; Ertz, D.; Diederich, P.; Lendemer, J.C.; May, P.; Schoch, C. L.; Arnold, A.E.; Gueidan, C.; Tripp, E.; Yahr, R.; Robertson, C.; Lutzoni, F. (2006). «New insights into classification and evolution of the Lecanoromycetes (Pezizomycotina, Ascomycota) from phylogenetic analyses of three ribosomal RNA- and two protein-coding genes». Mycologia. 98 (6): 1088–1103. PMID 17486983. doi:10.3852/mycologia.98.6.1088
- ↑ Peršoh, Derek; Beck, Andreas; Rambold, Gerhard (2004). «The distribution of ascus types and photobiontal selection in Lecanoromycetes (Ascomycota) against the background of a revised SSU nrDNA phylogeny». Mycological Progress. 3 (2): 103–121. Bibcode:2004MycPr...3..103P. doi:10.1007/s11557-006-0081-0
- ↑ a b c d Schmull, Michaela; Miadlikowska, Jolanta; Pelzer, Monika; Stocker-Wörgötter, Elfie; Hofstetter, Valerie; Fraker, Emily; Hodkinson, Brendan P.; Reeb, Valerie; Kukwa, Martin; Lumbsch, H. Thorsten; Kauff, Frank; Lutzoni, François (2011). «Phylogenetic affiliations of members of the heterogeneous lichen-forming fungi of the genus Lecidea sensu Zahlbruckner (Lecanoromycetes, Ascomycota)». Mycologia. 103 (5): 983–1003. PMID 21642348. doi:10.3852/10-234
- ↑ Vários estudos publicados nas décadas de 1970 e 1980 demonstraram a natureza polifilética da versão de Zahlbruckner de Lecidea, incluindo:
- Poelt, Josef (1973). «Systematic evaluation of morphological characters». In: Ahmadjian, Vernon; Hale, Mason E. The Lichens. New York: Academic Press. pp. 91–115. ISBN 978-0-12-044950-7
- Hertel, H. (1983). «Über einige aus Lecidea und Melanolecia (Ascomycetes lichenisati) auszuschliessende Arten» [Sobre algumas espécies a serem excluídas de Lecidea e Melanolecia (Ascomycetes lichenisati)]. Mitteilungen aus der Botanischen Staatssammlung München. 19: 441–447
- Hertel, H. (1984). Über saxicole, lecideoide Flechten der Subantarktis [Sobre líquenes saxícolas leideoide da Subantártida]. Col: Beihefte zur Nova Hedwigia (em alemão). 79. [S.l.]: J. Cramer. pp. 399–499. ISBN 978-3-7682-5479-3
- Rambold, G. (1985). «Lecidea sect. Armeniacae: lecideoide Arten der Flechtengattungen Lecanora und Tephromela (Lecanorales)» [Lecidea seção Armeniacae: espécies leideoide dos gêneros de líquenes Lecanora e Tephromela (Lecanorales)]. Botanische Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie (em alemão). 107: 469–501
- Timdal, E. (1987). «Problems of generic delimitation among squamiform members of the Lecideaceae». In: Peveling, Elisabeth. Progress and Problems in Lichenology in the Eighties: proceedings of an international symposium held at the University of Münster, 16.-21.3. 1986. Col: Bibliotheca Lichenologica. 25. [S.l.]: Lubrecht & Cramer. pp. 243–247. ISBN 978-3-443-58004-9
- ↑ a b c Kirk, Paul M.; Cannon, Paul F.; Minter, David W.; Stalpers, Joost A., eds. (2008). Dictionary of the Fungi 10th ed. Wallingford, UK: CAB Europe. p. 366. ISBN 978-0-85199-826-8
- ↑ Vainio, Edvard August (1934). «Lichenographia Fennica IV. Lecideales 2» (PDF). Acta Societatis Pro Fauna et Flora Fennica (em latim). 57 (2): 8
- ↑ Kraichak, Ekaphan; Huang, Jen-Pan; Nelsen, Matthew; Leavitt, Steven D.; Lumbsch, H. Thorsten (2018). «A revised classification of orders and families in the two major subclasses of Lecanoromycetes (Ascomycota) based on a temporal approach». Botanical Journal of the Linnean Society. 188 (3): 233–249. doi:10.1093/botlinnean/boy060
- ↑ Hawksworth, David L. (1974). Mycologist's Handbook. Kew: Commonwealth Mycological Institute. p. 39. ISBN 978-0-85198-300-4
- ↑ Ulloa, Miguel; Aguirre-Acosta, Elvira (2020). Illustrated Generic Names of Fungi. St. Paul, Minnesota: APS press. p. 210. ISBN 978-0-89054-618-5
- ↑ a b c Cannon, Paul F.; Kirk, Paul M. (2007). Fungal Families of the World. Wallingford: CAB International. 184 páginas. ISBN 978-0-85199-827-5. OCLC 60741230
- ↑ a b c Xie, Cong-Miao; Wang, Li-Song; Zhao, Zun-Tian; Zhang, Yan-Yun; Wang, Xin-Yu; Zhang, Lu-Lu (2022). «Revision of Immersaria and a new lecanorine genus in Lecideaceae (lichenised Ascomycota, Lecanoromycetes)». MycoKeys. 87: 99–132. PMC 8863769
 . PMID 35210924. doi:10.3897/mycokeys.87.72614
. PMID 35210924. doi:10.3897/mycokeys.87.72614
- ↑ a b c McCune, Bruce; Curtis, Marc J.; Di Meglio, Joseph (2017). «New taxa and a case of ephemeral spore production in Lecideaceae from western North America». The Bryologist. 120 (2): 115–124. doi:10.1639/0007-2745-120.2.115
- ↑ a b Jahamate, Aerguli; Abbas, Abudulla (2018). «The research progress of the genus Lecidea and identification of some Lecidea, Lecidella and Porpidia species in many areas». Botanical Research (em chinês). 7 (2): 93–100
- ↑ Gallun, Margalith; Ben-Shaul, Y.; Paran, Navah (1971). «The fungus-alga association in the Lecideaceae: an ultrastructural study». New Phytologist. 70 (3): 483–485. doi:10.1111/j.1469-8137.1971.tb02549.x
- ↑ Beck, A.; Friedl, T.; Rambold, G. (1998). «Selectivity of photobiont choice in a defined lichen community: inferences from cultural and molecular studies». New Phytologist. 139 (4): 709–720. doi:10.1046/j.1469-8137.1998.00231.x
- ↑ Hauck, Markus; Huneck, Siegfried; Elix, John A.; Paul, Paul (2007). «Does secondary chemistry enable lichens to grow on iron-rich substrates?». Flora. 202: 471–478. doi:10.1016/j.flora.2006.08.007. hdl:1885/35655
- ↑ Frank‐Kamenetskaya1, Olga V.; Zelenskaya, Marina S.; Izatulina1, Alina R.; Vereshchagin, Oleg S.; Vlasov, Dmitry Yu.; Himelbrant, Dmitry E.; Pankin, Dmitrii V. (2021). «Copper oxalate formation by lichens and fungi». Scientific Reports. 11: e24329. doi:10.1038/s41598-021-03600-5. hdl:11701/38652
- ↑ Kirk, Paul M.; Cannon, Paul F.; David, J.C.; Stalpers, Joost A. (2001). «Teloschistaceae». Ainsworth & Bisby's Dictionary of the Fungi 9th ed. Oxon, UK: CABI Bioscience. p. 274. ISBN 978-0-85199-377-5
- ↑ a b c d e Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; et al. (2022). «Outline of Fungi and fungus-like taxa – 2021». Mycosphere. 13 (1): 53–453. doi:10.5943/mycosphere/13/1/2. hdl:10481/76378
- ↑ «Lecideaceae». Catalogue of Life. Species 2000: Leiden, the Netherlands. Consultado em 11 de fevereiro de 2024
- ↑ Norman, J.M. (1852). «Conatus praemissus redactionis novae generum nonnullorum Lichenum in organis fructificationes vel sporis fundatae» [The effort put forward for the new edition of some genera of lichens based on fruiting bodies or spores]. Nytt Magazin for Naturvidenskapene (em latim). 7: 213–252 [230]
- ↑ a b Kalb, K. 1984. Lichenes Neotropici. 8: 301–350.
- ↑ Clauzade, G.; Roux, C. (1984). «Les genres Aspicilia Massal. et Bellemerea Hafellner & Roux.» [The genera Aspicilia Massal. and Bellemerea Hafellner & Roux] (PDF). Bulletin de la Société Botanique du Centre-Ouest (em esperanto). 15: 127–141
- ↑ Fryday, Alan M.; Printzen, Christian; Ekman, Stefan (2014). «Bryobilimbia, a new generic name for Lecidea hypnorum and closely related species». The Lichenologist. 46 (1): 25–37. doi:10.1017/s0024282913000625
- ↑ Brusse, Franklin A. (1994). «A remarkable new lichen genus Catarrhospora (Ascomycotina, Porpidiaceae), from Cape Floral Kingdom, South Africa». Mycotaxon. 52 (2): 501–512
- ↑ Triebel, D.; Rambold, G. (1988). «Cecidonia und Phacopsis (Lecanorales): zwei lichenicole Pilzgattungen mit cecidogenen Arten» [Cecidonia and Phacopsis (Lecanorales): two lichenicolous fungal genera with gall-inducing species]. Nova Hedwigia (em alemão). 47 (3–4): 279–309
- ↑ Hertel, H. (1983). «Über einige aus Lecidea und Melanolecia (Ascomycetes lichenisati) auszuschliessende Arten» [About some species to be excluded from Lecidea and Melanolecia (Ascomycetes lichenisati)]. Mitteilungen aus der Botanischen Staatssammlung München. 19: 441–447
- ↑ a b Rambold, G. (1989). «A Monograph of the Saxicolous Lecideoid Lichens of Australia (excl. Tasmania)». Berlin/Stuttgart: J. Cramer. Bibliotheca Lichenologica. 34: 239. ISBN 978-3-443-58013-1
- ↑ Fryday, Alan M.; Götz, Anna M.; Ruprecht, Ulrike (2024). «Imsharria orangei (Ascomycota, Lecideaceae), a new genus and species, and a new species of Porpidia, from the Falkland Islands». The Lichenologist. 56 (4): 137–147. doi:10.1017/S0024282924000148
- ↑ Malcolm, W.M.; Elix, J.A.; Owe-Larsson, B. (1995). «Labyrintha implexa (Porpidiaceae), a new genus and species from New Zealand». The Lichenologist. 27 (4): 241–248. doi:10.1006/lich.1995.0022
- ↑ Hertel, H. (1981). «Beiträge zur Kenntnis der Flechtenfamilie Lecideaceae VIII» [Contributions to the knowledge of the lichen family Lecideaceae VIII]. Herzogia (em alemão). 5 (3–4): 449–463. doi:10.1127/herzogia/5/1981/449
- ↑ Poelt, J.; Vězda, A. (1981). Bestimmungsschlüssel europäischer Flechten. Ergänzungsheft II [Identification key to European lichens. Supplement II]. Col: Bibliotheca Lichenologica (em alemão). 16. Vaduz: J. Cramer. p. 364. ISBN 978-3-7682-1162-8
- ↑ Harris, Richard C.; Ladd, Douglas (2007). «New taxa of lichens and lichenicolous fungi from the Ozark Ecoregion» (PDF). Opuscula Philolichenum. 4: 57–68
- ↑ a b c d Hertel, H. (1984). Über saxicole, lecideoide Flechten der Subantarktis [On saxicolous, lecideoid lichens of the Subantarctic]. Col: Beihefte zur Nova Hedwigia (em alemão). 79. [S.l.]: J. Cramer. pp. 399–499. ISBN 978-3-7682-5479-3
- ↑ Körber, G.W. (1855). Systema lichenum Germaniae (em latim). Breslau: Verlag von Trewendt & Granier. p. 221
- ↑ Timdal, E. (2010). «Porpidinia (Porpidiaceae), a new genus for Toninia tumidula». In: Hafellner, Josef; Kärnefelt, Ingvar; Wirth, Volkmar. Diversity and Ecology of Lichens in Polar and Mountain Ecosystems. Col: Bibliotheca Lichenologica. 104. [S.l.]: J. Cramer. pp. 333–337. ISBN 978-3-443-58083-4
- ↑ Zahlbruckner, A. (1923). Catalogus Lichenum Universalis. 2. [S.l.: s.n.] p. 686
- ↑ Timdal, E. (2007). «Romjularia». In: Nash III, Thomas H.; Gries, Corinna; Bungartz, Frank. Lichen Flora of the Greater Sonoran Desert Region. 3. Tempe: Lichens Unlimited, Arizona State University. pp. 287–289
- ↑ Brusse, Franklin A. (1988). «Schizodiscus, a new porpidioid lichen genus from the Drakensberg». Bothalia. 18: 94–96. doi:10.4102/abc.v18i1.988
- ↑ Hertel, H. (1967). Revision einiger calciphiler Formenkreise der Flechtengattung Lecidea [Revision of some calciphilous species groups of the lichen genus Lecidea]. Col: Beihefte zur Nova Hedwigia (em alemão). 24. [S.l.]: J. Cramer. p. 124. ISBN 978-3-768-25424-3
- ↑ Hertel, Hannes. (1983). Lecideaceae exsiccatae, Fascicle V (München): no. 96.
- ↑ a b Fryday, Alan M.; Hertel, Hannes (2014). «A contribution to the family Lecideaceae s. lat. (Lecanoromycetidae inc. sed. , lichenized Ascomycota) in the southern subpolar region; including eight new species and some revised generic circumscriptions». The Lichenologist. 46 (3): 389–412. doi:10.1017/S0024282913000704
- ↑ Ekman, Stefan (2001). «Molecular phylogeny of the Bacidiaceae (Lecanorales, lichenized Ascomycota)». Mycological Research. 105 (7): 783–797. doi:10.1017/S0953756201004269
- ↑ Awasthi, Dharani Dhar; Singh, Krishna Pal (1977). «Heppsora, a new lichen genus from India». The Bryologist. 80 (3): 536–538. JSTOR 3242031. doi:10.2307/3242031
- ↑ Bendiksby, Mika; Timdal, Einar (2013). «Molecular phylogenetics and taxonomy of Hypocenomyce sensu lato (Ascomycota: Lecanoromycetes): Extreme polyphyly and morphological/ecological convergence». Taxon. 62 (5): 940–956. doi:10.12705/625.18
- ↑ Coleine, Claudia; Stajich, Jason E.; Zucconi, Laura; Onofri, Silvano; Pombubpa, Nuttapon; Egidi, Eleonora; Franks, Ashley; Buzzini, Pietro; Selbmann, Laura (2018). «Antarctic cryptoendolithic fungal communities are highly adapted and dominated by Lecanoromycetes and Dothideomycetes». Frontiers in Microbiology. 9: 1–14. PMC 6033990. PMID 30008702. doi:10.3389/fmicb.2018.01392
- ↑ Diederich, Paul; Lawrey, James D.; Ertz, Damien (2018). «The 2018 classification and checklist of lichenicolous fungi, with 2000 non-lichenized, obligately lichenicolous taxa». The Bryologist. 121 (3): 373–374. doi:10.1639/0007-2745-121.3.340
- ↑ McCarroll, Danny; Viles, Heather (1995). «Rock-weathering by the lichen Lecidea auriculata in an arctic alpine environment». Earth Surface Processes and Landforms. 20 (3): 199–206. doi:10.1002/esp.3290200302
- ↑ Matthews, John A.; Owen, Geraint (2008). «Endolithic lichens, rapid biological weathering and Schmidt Hammer r-values on recently exposed rock surfaces: Storbreen glacier foreland, Jotunheimen, Norway». Geografiska Annaler. 90 (4): 287–297. JSTOR 40204904. doi:10.1111/j.1468-0459.2008.00346.x
- ↑ De los Ríos, A.; Sancho, L.G.; Grube, M.; Wierzchos, J.; Ascaso, C. (2005). «Endolithic growth of two Lecidea lichens in granite from continental Antarctica detected by molecular and microscopy techniques». New Phytologist. 165: 181–190. doi:10.1111/j.1469-8137.2004.01199.x
- ↑ Cooks, J.; Otto, E. (1990). «The weathering effects of the lichen Lecidea aff. sarcogynoides (Koerb.) on magaliesberg quartzite». Earth Surface Processes and Landforms. 15 (6): 491–500. doi:10.1002/esp.3290150602
- ↑ Favero-Longo, Sergio E.; Castelli, Daniele; Salvadori, Ornella; Belluso, Elena; Piervittori, Rosanna (2005). «Pedogenetic action of the lichens Lecidea atrobrunnea, Rhizocarpon geographicum gr. and Sporastatia testudinea on serpentinized ultramafic rocks in an alpine environment». International Biodeterioration & Biodegradation. 56 (1): 17–27. ISBN 978-90-481-6724-1. doi:10.1016/j.ibiod.2004.11.006
- ↑ Acaso, C.; Sancho, L.G.; Rodriguez-Pascual, C. (1990). «The weathering action of saxicolous lichens in maritime Antarctica» (PDF). Polar Biology. 11: 33–39
- ↑ Edwards, Howell G.M.; Jorge Villar, Susana E.; Seaward, Mark R.D.; St. Clair, Larry L. (2004). «Raman spectroscopy of rock biodeterioration by the lichen Lecidea tessellata flörke in a desert environment, Utah, USA». In: St.Clair, L.L.; Seaward, M.R.D. Biodeterioration of Stone Surfaces. Dordrecht: Springer. doi:10.1007/978-1-4020-2845-8_13
- ↑ Watanabe, Katsuaki; Kitagawa, Ryuji (2006). «Dissolution process of sekishu roof liles affected by crustose lichen Lecidea sp. grown on their surfaces». Clay Science. 12 (S2): 240–244
- ↑ Watanabe, K.; Ohfuji, H.; Ando, J.; Kitagawa, R. (2006). «Elemental behaviour during the process of corrosion of sekishu glazed roof-tiles affected by Lecidea s.lat. sp. (crustose lichen)». Clay Minerals. 41 (4): 819–826. doi:10.1180/0009855064140221
- ↑ Watanabe, Katsuaki; Kitagawa, Ryuji (2004). «Alteration mechanism of biotite in granitic rock caused by crustose lichen Porpidia». Clay Science. 12 (4): 249–257. doi:10.11362/jcssjclayscience1960.12.249
- ↑ Prieto, B.; Edwards, H.G.M.; Seaward, M.R.D. «A Fourier transform-Raman spectroscopic study of lichen strategies on granite monuments». Geomicrobiology Journal. 17 (1): 55–60. doi:10.1080/014904500270495
- ↑ Moncada, B.; Simijaca, D.; Soto-Medina, E.; Coco, L.F.; Jaramillo, M. (2020). «Lecidea mayeri». Lista Vermelha de Espécies Ameaçadas. 2020. Consultado em 11 de fevereiro de 2024
- ↑ a b Fryday, A. (2020). «Immersaria fuliginosa». Lista Vermelha de Espécies Ameaçadas. 2020. Consultado em 11 de fevereiro de 2024