Hyalospheniidae
Hyalospheniidae
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Devoniano Médio[1] 370–0 Ma | |||||||||||||||||
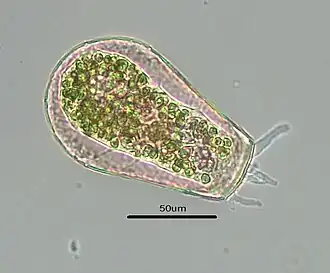 Hyalosphenia papilio | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Género-tipo | |||||||||||||||||
| Hyalosphenia Stein, 1857 | |||||||||||||||||
| Gêneros | |||||||||||||||||
| Sinónimos[6] | |||||||||||||||||
Nebelidae Taranek, 1882[5]
| |||||||||||||||||
Hyalospheniidae é uma família de amebas testáceas da ordem Arcellinida [en] e a única família da infraordem Hyalospheniformes. Essas amebas lobosas são caracterizadas pela capacidade de gerar uma concha composta por matéria orgânica ou partículas silicosas, que podem ser recicladas de amebas da ordem Euglyphida [en]. Elas habitam solo ou ambientes de água doce, sendo abundantes em musgos Sphagnum.
As amebas Hyalospheniidae surgiram após o Devoniano Médio, há cerca de 370 milhões de anos.[1] São consideradas importantes bioindicadores, frequentemente usadas para monitoramento ambiental. Seus fósseis são estudados para investigar a paleoecologia de habitats de zonas úmidas pré-históricos. A classificação das amebas Hyalospheniidae mudou várias vezes desde o século XIX com base em critérios morfológicos. Inicialmente classificadas como duas famílias distintas, Hyalospheniidae e Nebelidae, elas foram posteriormente consideradas sinônimas por meio de análises filogenéticas.
Morfologia
Os membros de Hyalospheniidae são amebas testáceas — seres unicelulares ameboides protistas que geram conchas aglutinadas com minerais. São caracterizadas por conchas ovoides, piriformes, em forma de vaso ou frasco, comprimidas lateralmente.[7][8] A construção e composição da concha variam significativamente dentro da família. Pode ser inteiramente secretada pelas próprias células e composta por uma matriz orgânica (por exemplo, Hyalosphenia), ou incluir escamas silicosas não orgânicas adicionais. Essas escamas minerais podem ser auto-secretadas (por exemplo, Quadrulella) ou recicladas de placas de conchas de pequenas amebas da ordem Euglyphida ou outros materiais similares, como frústulas de diatomáceas (por exemplo, Apodera [en], Padaungiella [en], Nebela [en]).[7] A característica de reciclar placas de conchas de amebas da ordem Euglyphida é conhecida como "cleptosquamia" e parece ser um traço ancestral dentro da família.[1]


Ecologia
As amebas Hyalospheniidae são consideradas importantes bioindicadores em estudos de monitoramento ambiental. Sua sensibilidade a mudanças ambientais, como poluição atmosférica,[9] torna-as indicadores confiáveis de mudanças hidrológicas. Juntamente com a preservação de suas conchas por milhares de anos, sua sensibilidade ambiental confere-lhes um papel proeminente na reconstrução do paleoclima em turfeiras, pântanos e charcos.[7]
Essa família inclui várias das amebas testáceas lobosas mais comuns e bem estudadas. Seus membros são especialmente diversos e abundantes em ecossistemas de zonas úmidas oligotróficos, como turfeiras dominadas por musgos Sphagnum. Algumas também podem ser encontradas em diferentes musgos, habitats de água doce e solo. Embora várias espécies de Hyalospheniidae tenham uma distribuição cosmopolita, muitas são restritas ao Hemisfério Sul e à faixa tropical dentro do Hemisfério Norte.[8]
Uma espécie, Hyalosphenia papilio, foi observada com endossimbiontes fotossintéticos (zooclorelas).[10] Essa espécie é um mixótrofo obrigatório, vivendo em associação constante com simbiontes intracelulares pertencentes à classe de algas verdes Trebouxiophyceae, especificamente algas da família Chlorellaceae [en].[11]
Evolução
| |||||||||||||||||||||||||||||||||||||||||||||
| Posição das amebas Hyalospheniidae na filogenia do grupo Arcellinida.[12] |
Hyalospheniidae é uma família de Arcellinida, uma ordem de amebas testáceas lobosas dentro do supergrupo eucariótico Amoebozoa. Em contraste com as amebas testáceas filosas, encontradas no supergrupo Rhizaria (por exemplo, amebas da ordem Euglyphida), elas apresentam pseudópodes mais espessos com extremidades rombudas. É a única família da infraordem Hyalospheniformes, que pertence à subordem Glutinoconcha. Glutinoconcha, que contém a maioria das espécies de Arcellinida, evoluiu de um ancestral comum com conchas aglutinadas com minerais, em contraste com as conchas orgânicas de Organoconcha. Em particular, Hyalospheniformes e Volnustoma, uma infraordem diferente de Glutinoconcha, evoluíram de ancestrais com conchas aglutinadas xenossômicas (isto é, compostas por partículas incorporadas de uma fonte externa).[2][12]
Por meio de aproximações de relógio molecular, a idade da Hyalospheniidae foi estimada em 2015 como cerca de 370 milhões de anos, entre o Devoniano e o início do Carbonífero. Essa reconstrução molecular sugere que as amebas Hyalospheniidae se diversificaram após o Devoniano Médio, quando a diversificação de plantas terrestres formou florestas extensas com uma produção abundante de matéria orgânica e solos. A cleptosquamia, a capacidade das amebas Hyalospheniidae de "roubar" escamas de concha de suas presas, amebas da ordem Euglyphida, é hipotetizada como um traço ancestral dentro da família. Essa hipótese de trabalho é baseada na presença de cleptosquamia na maioria das amebas Hyalospheniidae.[1] Além das estimativas do relógio molecular, foi sugerido que microfósseis em forma de vaso de 750 milhões de anos poderiam pertencer a essa família.[13][7][14]
Sistemática
História da taxonomia
Houve várias tentativas de classificar Hyalospheniidae entre as amebas testáceas, bem como sua classificação interna. O paleontólogo americano Joseph Leidy, em 1874, foi possivelmente o primeiro a notar características comuns entre as células. Ele descreveu as testas em forma de vaso como compostos por pequenas partículas silicosas ("placas discoides e hastes minúsculas") capturadas dentro de uma matriz orgânica, interpretada como originada pela ameba ("intrínseca"). Ele agrupou essas espécies no gênero Nebela, distinguindo-as do gênero previamente conhecido de amebas testáceas Difflugia. Em vez disso, ele descreveu as espécies de Difflugia como tendo uma "testa composta de corpos estranhos, como partículas de areia de quartzo e proteções de diatomáceas".[15][7]
Em 1877, o zoólogo alemão Franz Eilhard Schulze [en] descreveu as famílias Hyalosphenidae, Arcellidae, Quadrulidae e Difflugidae. Amebas com um testa orgânica homogênea, como Hyalosphenia, foram colocadas em Hyalospheniidae, enquanto Nebela foi colocada em Difflugidae, e Quadrulella em Quadrulidae.[3][7]
Em 1882, Taranek descreveu a família Nebelidae para incluir amebas com placas silicosas: Nebela, Lesquereusia, Quadrulella, Corythion (que foi posteriormente excluído), Amphizonella, Cochliopodium [en], Hyalosphenia, Leptochlamys e Zonomyxa.[5] Essa família foi redefinida em 1942 por Jung e organizada em treze gêneros recém-descritos por ele: Alocodera, Apodera [en], Argynnia, Deflandria, Nebela, Leidyella, Penardiella, Physochila, Porosia [en], Pterygia, Quadrulella, Schaudinnia e Umbonaria.[16] No entanto, Jung não designou espécie-tipo em sua classificação, o que invalidou todos os gêneros com mais de uma espécie.[a] Consequentemente, apenas gêneros monotípicos como Alocodera, Physochila e Porosia foram reconhecidos, e os demais gêneros foram absorvidos por Nebela. Posteriormente, ao designar tipos, os micropaleontólogos Alfred R. Loeblich Jr [en]. e Helen Niña Tappan Loeblich [en] validaram Apodera e Certesella [en] em 1961,[18] e Vucetich validou Argynnia em 1974.[19]
Em 2002, o protozoólogo alemão Ralf Meisterfeld publicou a última revisão da família baseada exclusivamente em caracteres morfológicos. Ele reclassificou Nebela e gêneros similares em duas famílias: Hyalospheniidae, composta por gêneros com testas rígidas, quitinoides e orgânicas (Hyalosphenia e Leptochlamys); e Nebelidae, composta por gêneros com testas construídas a partir de placas de pequenas amebas da ordem Euglyphida ou fragmentos de diatomáceas (Apodera, Argynnia, Certesella, Nebela, Physochila, Porosia, Schoenbornia). Seguindo uma classificação de 1979,[20] ele excluiu o gênero Quadrulella para a família Lesquereusiidae junto com outros gêneros da ordem Arcellinida com hastes silicosas auto-secretadas em suas testas.[21][7]
As primeiras análises filogenéticas de Arcellinida baseadas em dados moleculares demonstraram que Nebela e Nebelidae não eram monofiléticos, ou seja, não formavam um clado ou grupo de táxons evoluídos de um ancestral comum sem incluir outros gêneros de Hyalospheniidae.[22] Espécies de Apodera, Porosia, Nebela e Hyalosphenia estavam entrelaçadas em um clado conhecido como "Nebelas centrais".[8] Devido a esse resultado e à presença de características distintivas de Hyalospheniidae em alguns Nebelidae, as duas famílias foram sinonimizadas sob o primeiro nome.[4] Além disso, muitas espécies do gênero polifilético Nebela foram separadas em novos gêneros monofiléticos, nomeadamente Padaungiella em 2012,[4] Cornutheca, Gibbocarina, Longinebela, Mrabella e Planocarina em 2016,[23] e finalmente Alabasta [en] em 2018.[24]
Classificação
| Filogenia das amebas Hyalospheniidae | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Baseado em uma análise filogenética de 2018 da maioria dos gêneros,[24] e uma análise de 2021 que recuperou Apodera [en] como o grupo irmão de Alocodera e Padaungiella [en].[25] Porosia [en], excluída das análises, é considerada um parente próximo de Certesella [en].[26] |
A taxonomia atual da família reconhece 14 gêneros,[2][12] com um total de 87 espécies:
- Alabasta [en] Duckert, Blandenier, Kosakyan & Singer 2018 — 3 espécies.[24]
- Alocodera Jung, 1942[27] — 1 espécie.[28]
- Apodera [en] Loeblich & Tappan, 1961 — 4 espécies.[7][25]
- Certesella [en] Loeblich & Tappan, 1961 — 5 espécies.[29][30]
- Cornutheca Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 3 espécies.[23]
- Gibbocarina Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 (=Umbonaria Jung, 1942)[31] — 3 espécies.[23][31]
- Hyalosphenia (Stein, 1857) Schulze [en], 1877 — 18 espécies.[7]
- Longinebela Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 5 espécies.[23][32]
- Mrabella Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 2 espécies.[23]
- Nebela [en] Leidy, 1874 em. Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 13 espécies.[23]
- Padaungiella [en] Lara et Todorov, 2012 (=Schaudinnia Jung, 1942) — 5 espécies.[4]
- Planocarina [en] Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 4 espécies.[23]
- Porosia [en] Jung, 1942[27] — 2 espécies.[26]
- Quadrulella Cockerell, 1909 em. Kosakyan, Lahr, Mulot, Meisterfeld, Mitchell & Lara, 2016 — 19 espécies.[7][23][33]
Notas
- ↑ Devido ao artigo 13.3 do Código Internacional de Nomenclatura Zoológica,[7] gêneros descritos após 1930 só podem ser validados se houver uma espécie-tipo designada para eles.[17]
Referências
- ↑ a b c d Lahr, Daniel JG; Bosak, Tanja; Lara, Enrique; Mitchell, Edward AD (setembro de 2015). «The Phanerozoic diversification of silica-cycling testate amoebae and its possible links to changes in terrestrial ecosystems». PeerJ. 3: e1234. PMC 4699787
 . PMID 26734499. doi:10.7717/peerj.1234
. PMID 26734499. doi:10.7717/peerj.1234
- ↑ a b c Lahr D, Kosakyan A, Lara E, Mitchell E, Morais L, Porfirio-Sousa AL, Ribeiro GM, Tice AK, Pánek T, Kang S, Brown MW (2019). «Phylogenomics and Morphological Reconstruction of Arcellinida Testate Amoebae Highlight Diversity of Microbial Eukaryotes in the Neoproterozoic». Current Biology. 29 (6): 991–1001. PMID 30827918. doi:10.1016/j.cub.2019.01.078. hdl:11380/1301486
- ↑ a b Schulze FE (1877). «Rhizopodenstudien VI». Archiv für Mikroskopische Anatomie. 13: 9–30. doi:10.1007/BF02933929. Consultado em 24 de setembro de 2023. Cópia arquivada em 25 de outubro de 2022
- ↑ a b c d Kosakyan A, Heger TJ, Leander BS, Todorov M, Mitchell EAD, Lara E (2012). «COI Barcoding of Nebelid Testate Amoebae (Amoebozoa: Arcellinida): Extensive Cryptic Diversity and Redefinition of the Hyalospheniidae Schulze». Protist. 163 (3): 415–434. PMID 22130576. doi:10.1016/j.protis.2011.10.003
- ↑ a b Taranek KJ (1882). «Monographie der Nebeliden Böhmens. Ein Beitrag zur Kenntnis der Süsswasser Monothalamien» [Monograph of the Nebelids of Bohemia. A contribution to the knowledge of freshwater monothalamids]. Abhandlungen der Königlichen Böhmischen Gesellschaft der Wissenschaften (em alemão). VI: 1–56
- ↑ Kosakyan, Anush; Gomaa, Fatma; Lara, Enrique; Lahr, Daniel J.G. (2016). «Current and future perspectives on the systematics, taxonomy and nomenclature of testate amoebae». European Journal of Protistology. 55B (Pt B): 105–117. PMID 27004416. doi:10.1016/j.ejop.2016.02.001
- ↑ a b c d e f g h i j k Kosakyan, Anush (16 de abril de 2014). Phylogeny, systematics and ecology of free living protists. Case study: family Hyalospheniidae (Tese). University of Neuchâtel. Consultado em 24 de setembro de 2023. Cópia arquivada em 29 de setembro de 2023
- ↑ a b c Lara E, Heger TJ, Ekelund F, Lamentowicz M, Mitchell EAD (2008). «Ribosomal RNA genes challenge the monophyly of the Hyalospheniidae (Amoebozoa: Arcellinida)». Protist. 159 (2): 165–176. PMID 18023614. doi:10.1016/j.protis.2007.09.003. Consultado em 22 de outubro de 2023. Cópia arquivada em 23 de junho de 2021
- ↑ Payne R, Mitchell E, Nguyen-Viet H, Gilbert D (2012). «Can pollution bias peatland paleoclimate reconstruction?». Quaternary Research. 78 (2): 170–173. Bibcode:2012QuRes..78..170P. doi:10.1016/j.yqres.2012.05.004
- ↑ Zagumyonnaya ON, Philippov DA, Zagumyonnyi DG, Komarov AA, Tsyganov AN, Tikhonenkov DV (2023). «Changes in Testate Amoeba Assemblages in a Series of Different Types of Aquatic and Terrestrial Habitats of Wetland and Forest Ecosystems». Biology Bulletin. 50: 1719–1737. doi:10.1134/S1062359023080332
- ↑ Gomaa, Fatma; Kosakyan, Anush; Heger, Thierry J.; Corsaro, Daniele; Mitchell, Edward A.D.; Lara, Enrique (2014). «One Alga to Rule them All: Unrelated Mixotrophic Testate Amoebae (Amoebozoa, Rhizaria and Stramenopiles) Share the Same Symbiont (Trebouxiophyceae)». Protist. 165 (2): 161–176. doi:10.1016/j.protis.2014.01.002
- ↑ a b c González-Miguéns, Rubén; Todorov, Milcho; Blandenier, Quentin; Duckert, Clément; Porfirio-Sousa, Alfredo L.; Ribeiro, Giulia M.; Ramos, Diana; Lahr, Daniel J.G.; Buckley, David; Lara, Enrique (2022). «Deconstructing Difflugia: The tangled evolution of lobose testate amoebae shells (Amoebozoa: Arcellinida) illustrates the importance of convergent evolution in protist phylogeny». Molecular Phylogenetics and Evolution. 175. 107557 páginas. PMID 35777650. doi:10.1016/j.ympev.2022.107557. hdl:10261/281619
- ↑ Porter SM, Meisterfeld R, Knoll AH (maio de 2003). «Vase-shaped microfossils from the Neoproterozoic Chuar Group, Grand Canyon: a classification guided by modern testate amoebae» (PDF). Journal of Paleontology. 77 (3): 409–429. doi:10.1666/0022-3360(2003)077<0409:VMFTNC>2.0.CO;2. Consultado em 24 de setembro de 2023. Cópia arquivada (PDF) em 8 de março de 2018
- ↑ Tingle KE, Porter SM, Raven MR, Czaja AD, Webb SM, Bloeser B (2023). «Organic preservation of vase-shaped microfossils from the late Tonian Chuar Group, Grand Canyon, Arizona, USA». Geobiology. 21 (3): 290–309. PMID 36651474. doi:10.1111/gbi.12544
- ↑ Leidy J (1874). «Notice of some new freshwater rhizopods» (PDF). Annals and Magazine of Natural History. 14 (83): 383–385. doi:10.1080/00222937408680997
- ↑ Jung W (1942). «Illustrierte Thekamöben-Bestimmungstabellen I. Die Systematik der Nebelinen» [Illustrated identification tables of thecate amoebae I. The systematics of the nebelines]. Archiv für Protistenkunde (em alemão). 95: 357–390
- ↑ «ICZN Code Art. 13». Consultado em 25 de setembro de 2023. Cópia arquivada em 5 de outubro de 2021
- ↑ Loeblich, Alfred Richard; Tappan, Helen Niña (1961). «Remarks on the systematics of the Sarkodina (Protozoa), renamed homonyms and new and validated genera». Proceedings of the Biological Society of Washington. 74: 213–234. Consultado em 24 de setembro de 2023. Cópia arquivada em 30 de setembro de 2023
- ↑ Vucetich MC (1974). «Comentarios criticos sobre Argynnia Jung, 1942 (Rhizopoda, Testacea)» [Critical comments on Argynnia Jung, 1942 (Rhizopoda, Testaeca)]. Neotropica (em espanhol). 20 (63): 126–128
- ↑ Ogden CG (1979). «Siliceous structures secreted by members of the subclass Lobosia (Rhizopodea, Protozoa)». Bulletin of the British Museum (Natural History) Zoology. 36: 203–207. Consultado em 22 de outubro de 2023. Cópia arquivada em 3 de outubro de 2023
- ↑ Meisterfeld R (2002). «Order Arcellinida Kent, 1880» (PDF). In: Lee JJ, Leedale GF, Bradbury P. An illustrated guide to the protozoa: organisms traditionally referred to as protozoa, or newly discovered groups 2nd ed. Lawrence, Kansas, USA: Society of protozoologists. pp. 827–860. Consultado em 25 de setembro de 2023. Cópia arquivada em 29 de setembro de 2023
- ↑ Nikolaev SI, Mitchell EA, Petrov NB, Berney C, Fahrni J, Pawlowski J (2005). «The testate lobose amoebae (order Arcellinida Kent, 1880) finally find their home within Amoebozoa». Protist. 156 (2): 191–202. PMID 16171186. doi:10.1016/j.protis.2005.03.002
- ↑ a b c d e f g h Kosakyan, Anush; Lahr, Daniel J. G.; Mulot, Matthieu; Meisterfeld, Ralf; Mitchell, Edward A. D.; Lara, Enrique (2016). «Phylogenetic reconstruction based on COI reshuffles the taxonomy of hyalosphenid shelled (testate) amoebae and reveals the convoluted evolution of shell plate shapes». Cladistics. 32 (6): 606–623. PMID 34727671. doi:10.1111/cla.12167
- ↑ a b c Duckert C, Blandenier Q, Kupferschmid FAL, Kosakyan A, Mitchell EAD, Lara E, Singer D (2018). «En garde! Redefinition of Nebela militaris (Arcellinida, Hyalospheniidae) and erection of Alabasta gen. nov.» (PDF). European Journal of Protistology. 66: 156–165. PMID 30366198. doi:10.1016/j.ejop.2018.08.005. Consultado em 3 de janeiro de 2023. Cópia arquivada (PDF) em 10 de junho de 2020
- ↑ a b Duckert C, Blandenier Q, McKeown M, Hohaia H, Luketa S, Wilmshurst J, et al. (2021). «Superficially described and ignored for 92 years, rediscovered and emended: Apodera angatakere (Amoebozoa: Arcellinida: Hyalospheniformes) is a new flagship testate amoeba taxon from Aotearoa (New Zealand)». Journal of Eukaryotic Microbiology. 68 (6): e12867. PMC 9292727. PMID 34351666. doi:10.1111/jeu.12867
- ↑ a b Bobrov, Anatoly; Kosakyan, Anush. «A New Species from Mountain Forest Soils in Japan: Porosia paracarinata sp. nov., and Taxonomic Concept of the Genus Porosia Jung, 1942». Acta Protozoologica. 54 (4): 289–294. doi:10.4467/16890027AP.15.024.3538
- ↑ a b Jung W (1942). «Südchilenische Thekamöben (Aus dem südchilenischen Küstengebiet, Beitrag 10)» [Southern Chilean thecamoeba (From the Southern Chilean coastal area, contribution 10)]. Archiv für Protistenkunde (em alemão). 95: 253–356
- ↑ Luketa, Stefan (2015). «Description of the family Padaungiellidae and morphological variability of Padaungiella lageniformis (Amoebozoides: Arcellinida) from the Vlasina Lake area, Serbia». Archives of Biological Sciences, Belgrade. 67 (4): 1331–1337. doi:10.2298/ABS150312110L
- ↑ Bobrov, Anatoly; Duckert, Clément; Mitchell, Edward A. D. (2021). «Certesella larai (Amoebozoa: Arcellinida: Hyalospheniformes) a new soil testate amoeba species from the Dominican Republic and Chile challenges the definition of genera Certesella and Porosia». Acta Protozoologica. 60: 61–75. doi:10.4467/16890027AP.21.007.15381
- ↑ Vucetich, María Cristina (1978). «Comentarios sobre el género Certesella Loeblich & Tappan, 1961 y estudio de la estéreo ultraestructura tecal de tres especies austroamericanas (Rhizopoda Testaceolobosa)» [Comments on the genus Certesella Loeblich & Tappan, 1961 and the study of the thecate stereo ultrastructure of three Austroamerican species (Rhizopoda Testacealobosa)] (PDF). Obra Centenario, Museo de la Plata, Zoología (em espanhol). 6: 305–313. Consultado em 23 de setembro de 2023. Cópia arquivada (PDF) em 20 de fevereiro de 2022
- ↑ a b Luketa, Stefan (2017). «Morphological variability of Gibbocarina galeata and G. penardiana comb. nov. (Arcellinida: Hyalospheniidae) from East Herzegovina». Protistology. 11 (1): 37–47. doi:10.21685/1680-0826-2017-11-1-3
- ↑ Todorov, Milcho; Bankov, Nikola; Ganeva, Anna (2018). «Longinebela ampulla sp. n. (Arcellinida: Hyalospheniidae), a New Testate Amoeba from Sphagnum Peatlands in Bulgaria» (PDF). Acta Zoologica Bulgarica. 70 (3): 285–292. Consultado em 22 de outubro de 2023. Cópia arquivada (PDF) em 13 de março de 2023
- ↑ Bankov, Nikola; Todorov, Milcho; Ganeva, Anna (2021). «An overview of the genus Quadrulella (Arcellinida: Hyalospheniidae) from Bulgaria, with description of Quadrulella deflandrei sp. nov.». Protistology. 15 (2): 53–68