Extinção dos Neandertais
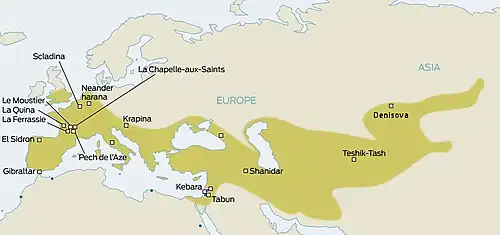
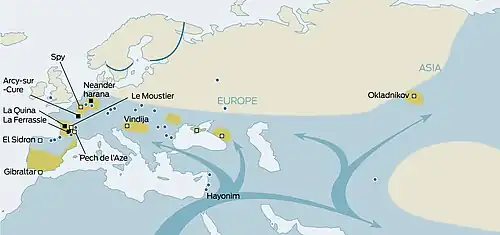
Os Neandertais tornaram-se extintos há cerca de 40.000 anos. As hipóteses sobre as causas de sua extinção incluem violência, transmissão de doenças dos humanos modernos para as quais os neandertais não tinham imunidade, substituição competitiva, extinção por cruzamento com populações humanas modernas iniciais, catástrofes naturais, mudanças climáticas e depressão de consanguinidade. É provável que múltiplos fatores tenham causado o desaparecimento de uma população já reduzida.
Transição para o Paleolítico Superior
A extinção dos neandertais fez parte do evento mais amplo de extinção da megafauna do Pleistoceno Superior [en].[1] Qualquer que tenha sido a causa de sua extinção, os neandertais foram substituídos pelos humanos modernos, indicado pela substituição quase completa da tecnologia lítica do Paleolítico Médio (Musteriense) pela tecnologia lítica do Paleolítico Superior dos humanos modernos (Aurignaciano) em toda a Europa (a Transição do Paleolítico Médio para o Superior) entre 41.000 e 39.000 anos atrás.[2][3][4][5] Entre 44 200 e 40 600 AP, os neandertais desapareceram do noroeste da Europa.[6] No entanto, postula-se que os neandertais ibéricos persistiram até cerca de 35.000 anos atrás, conforme indicado pelo intervalo de datas dos conjuntos líticos de transição—Chatelperronense, Uluzziano, Protoaurinhacense e Aurinhacense Inicial. Os dois últimos são atribuídos a humanos modernos, mas os dois primeiros têm autoria não confirmada, potencialmente produtos de coabitação e transmissão cultural entre neandertais e humanos modernos. Além disso, o aparecimento do Aurinhacense ao sul do Rio Ebro foi datado em aproximadamente 37 500 anos atrás, o que levou à hipótese da "Fronteira do Ebro", que afirma que o rio representava uma barreira geográfica impedindo a imigração de humanos modernos e, assim, prolongando a persistência neandertal.[7][8] No entanto, a datação da Transição Ibérica é debatida, com um tempo contestado de 43.000–40 800 anos atrás na Caverna Bajondillo [en], Espanha.[9][10][11][12] O Chatelperronense aparece no nordeste da Ibéria há cerca de 42 500–41 600 anos.[7]
Alguns fósseis de neandertais [en] foram datados como muito mais recentes—como os encontrados em Zafarraya (30.000 anos atrás)[13] e na Caverna de Gorham (28.000 anos atrás)[14]—o que pode ser impreciso, pois foram baseados em artefatos ambíguos em vez de datação direta.[4] Uma afirmação de neandertais sobrevivendo em um refúgio polar nos Montes Urais[15] é vagamente apoiada por ferramentas de pedra musterenhas datadas de 34.000 anos atrás no sítio siberiano setentrional de Byzovaya, em um momento em que os humanos modernos ainda não teriam colonizado as regiões mais ao norte da Europa;[16] no entanto, restos humanos modernos são conhecidos do sítio próximo de Mamontovaya Kurya [en], datado de 40.000 anos atrás.[17] A datação indireta de restos de neandertais da Caverna de Mezmaiskaya [en] reportou uma data de cerca de 30.000 anos atrás, mas a datação direta, em vez disso, resultou em 39 700±1 100 anos atrás, mais alinhada com as tendências exibidas no resto da Europa.[18]

A primeira indicação de imigração de humanos modernos do Paleolítico Superior para a Europa é uma série de dentes humanos modernos com ferramentas de pedra da indústria Neroniana encontradas na Caverna de Mandrin [en], Malataverne, na França, datadas em 2022 entre 56 800 e 51 700 anos atrás.[19] Os ossos mais antigos na Europa datam de aproximadamente 45–43.000 anos atrás na Bulgária,[20] Itália,[21] e Grã-Bretanha.[22] Esta onda de humanos modernos substituiu os neandertais.[2] No entanto, neandertais e H. sapiens têm uma história de contato muito mais longa. Evidências de DNA indicam contato de H. sapiens com neandertais e mistura genética já há 120–100.000 anos. Uma reanálise em 2019 de fragmentos de crânio de 210.000 anos da Caverna Apidima, na Grécia, presumidos como pertencentes a um neandertal, concluiu que pertenciam a um humano moderno, e um crânio de neandertal datado de 170.000 anos atrás na gruta indica que H. sapiens foram substituídos por neandertais até retornarem há cerca de 40.000 anos.[23] Esta identificação foi refutada por um estudo de 2020.[24]
Possíveis causas da extinção
Violência
Kwang Hyun Ko discute a possibilidade de que a extinção dos neandertais tenha sido precipitada ou acelerada por conflitos violentos com Homo sapiens. A violência nas sociedades de caçadores-coletores primitivos geralmente ocorria como resultado da competição por recursos após desastres naturais. Portanto, é plausível sugerir que a violência, incluindo a guerra primitiva, teria ocorrido entre as duas espécies humanas.[25] A hipótese de que os humanos modernos substituíram violentamente os neandertais foi proposta pela primeira vez pelo paleontólogo francês Marcellin Boule (a primeira pessoa a publicar uma análise de um neandertal) em 1912.[26]
Parasitas e patógenos
Doenças infecciosas carregadas por Homo sapiens podem ter sido transmitidas aos neandertais, que teriam tido pouca proteção contra infecções às quais não haviam sido previamente expostos, levando a consequências devastadoras para as populações neandertais. Homo sapiens eram menos vulneráveis a doenças neandertais, em parte porque haviam evoluído para lidar com uma carga de doenças muito maior nos trópicos e, portanto, eram mais capazes de lidar com patógenos novos, e em parte porque o maior número de Homo sapiens significava que mesmo surtos devastadores ainda deixariam sobreviventes suficientes para uma população viável.[27]
Se os vírus pudessem saltar facilmente entre essas duas espécies semelhantes, possivelmente porque viviam próximas, Homo sapiens pode ter infectado os neandertais e impedido que a epidemia se extinguisse à medida que os números neandertais diminuíam. O mesmo processo também pode explicar a resiliência dos Homo sapiens a doenças e parasitas neandertais. Novas doenças humanas provavelmente se moveram da África para a Eurásia. Essa suposta "vantagem africana" permaneceu até a revolução agrícola há 10.000 anos na Eurásia, após a qual animais domesticados superaram outros primatas como a fonte mais prevalente de novas infecções humanas, substituindo a "vantagem africana" por uma "vantagem eurasiana". O impacto catastrófico dos vírus eurasianos nas populações nativas americanas no passado histórico oferece uma noção de como os humanos modernos podem ter afetado grupos predecessores hominínios na Eurásia há 40.000 anos. Os genomas humanos e neandertais e adaptações a doenças ou parasitas podem fornecer insight sobre isso.[28][29]
As interações de doenças infecciosas podem expressar o período prolongado de estagnação antes da modificação, conforme a ecologia de doenças. Modelos matemáticos foram usados para fazer previsões para investigações futuras, fornecendo informações sobre interações entre espécies durante a transição entre as eras do Paleolítico Médio e Superior. Isso pode ser útil dada a escassa evidência material deste período e o potencial da tecnologia de sequenciamento e datação de DNA. Tal modelagem, juntamente com tecnologia moderna e metodologias arqueológicas pré-históricas, pode fornecer uma nova compreensão deste período na origem humana.[29]
No final do século XX na Nova Guiné, devido a práticas funerárias canibalísticas, o povo Fore [en] foi dizimado por encefalopatias espongiformes transmissíveis, especificamente o kuru, uma doença altamente virulenta disseminada pela ingestão de príons encontrados no tecido cerebral. No entanto, indivíduos com a variante 129 do gene PRNP eram naturalmente imunes aos príons. O estudo deste gene levou à descoberta de que a variante 129 era difundida entre todos os humanos modernos, o que poderia indicar canibalismo generalizado em algum momento da pré-história humana. Como os neandertais são conhecidos por terem praticado canibalismo até certo ponto e por terem coexistido com humanos modernos, o paleoantropólogo britânico Simon Underdown especulou que os humanos modernos transmitiram uma doença espongiforme semelhante ao kuru aos neandertais e, como a variante 129 parece ter estado ausente nos neandertais, ela os matou rapidamente.[30][31]
Substituição competitiva
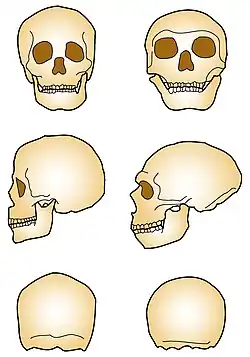
Desvantagens específicas da espécie
Uma ligeira vantagem competitiva por parte dos humanos modernos pode ter sido responsável pelo declínio dos neandertais em uma escala de tempo de milhares de anos.[32][33]
Geralmente, sítios fósseis pequenos e amplamente dispersos sugerem que os neandertais viviam em grupos menos numerosos e socialmente mais isolados do que os Homo sapiens contemporâneos. Ferramentas como lâminas de sílex musterienses e pontas Levallois são notavelmente sofisticadas desde o início, mas têm uma taxa lenta de variabilidade e uma inércia tecnológica geral é perceptível durante todo o período fóssil. Os artefatos são de natureza utilitária, e traços comportamentais simbólicos não são documentados antes da chegada dos humanos modernos na Europa, há cerca de 40.000 a 35.000 anos.[32][34][35]
As diferenças morfológicas perceptíveis na forma do crânio entre as duas espécies humanas também têm implicações cognitivas. Estas incluem os lobos parietais[36][37][38] e cerebelo[39][40] menores dos neandertais, áreas implicadas no uso de ferramentas,[41] integração visuoespacial,[42] habilidades numéricas,[43] criatividade,[44] e conceitualização de ordem superior.[45] As diferenças, embora leves, possivelmente teriam sido suficientes para afetar a seleção natural e podem fundamentar e explicar as diferenças nos comportamentos sociais, na inovação tecnológica e na produção artística.[32]
Jared Diamond, um defensor da substituição competitiva, aponta em seu livro O Terceiro Chimpanzé [en] [The Third Chimpanzee] que a substituição dos neandertais pelos humanos modernos é comparável a padrões de comportamento que ocorrem sempre que pessoas com tecnologia avançada entram em conflito com pessoas com tecnologia menos desenvolvida.[46]
Divisão do trabalho
Em 2006, foi postulado que a divisão do trabalho entre os sexos dos neandertais era menos desenvolvida do que a dos Homo sapiens do Paleolítico Médio. Tanto os neandertais masculinos quanto femininos participavam da ocupação única de caçar grandes animais, como bisões, veados, gazelas e cavalos selvagens. Esta hipótese propõe que a relativa falta de divisão de trabalho dos neandertais resultou em uma extração menos eficiente de recursos do ambiente em comparação com Homo sapiens.[47]
Diferenças anatômicas e habilidade de corrida
Pesquisadores como Karen L. Steudel da Universidade de Wisconsin destacaram a relação entre a anatomia neandertal (mais baixa e atarracada do que a dos humanos modernos) e a capacidade de correr e o requerimento de energia (30% a mais).[48]
No entanto, em um estudo recente, os pesquisadores Martin Hora e Vladimir Sladek da Universidade Carolina de Praga mostram que a configuração dos membros inferiores dos neandertais, particularmente a combinação de joelhos robustos, calcanhares longos e membros inferiores curtos, aumentou a vantagem mecânica efetiva dos extensores do joelho e tornozelo dos neandertais, reduzindo assim significativamente a força necessária e a energia gasta para a locomoção. O custo de caminhada do neandertal masculino agora é estimado em 8–12% maior do que o dos homens anatomicamente modernos, enquanto o custo de caminhada da neandertal feminina é considerado virtualmente igual ao das mulheres anatomicamente modernas.[49]
Outros pesquisadores, como Yoel Rak, da Universidade de Tel Aviv em Israel, notaram que os registros fósseis mostram que as pélvis neandertais, em comparação com as pélvis humanas modernas, teriam tornado muito mais difícil para os neandertais absorverem choques e saltarem de um passo para o seguinte, dando aos humanos modernos outra vantagem sobre os neandertais em capacidade de corrida e caminhada. No entanto, Rak também observa que todos os humanos arcaicos tinham pélvis largas, indicando que esta é a morfologia ancestral e que os humanos modernos sofreram uma mudança para pélvis mais estreitas no final do Pleistoceno.[50]
Humanos modernos e aliança com cães
Pat Shipman argumenta que a domesticação do cão deu aos humanos modernos uma vantagem ao caçar.[51] Evidências mostram que os restos mais antigos de cães domesticados foram encontrados na Bélgica (31 700 AP) e na Sibéria (33.000 AP).[52][53] Um levantamento de sítios antigos de humanos modernos e neandertais com restos faunísticos em toda Espanha, Portugal e França forneceu uma visão geral do que humanos modernos e neandertais comiam.[54] Coelhos tornaram-se mais frequentes, enquanto grandes mamíferos – principalmente consumidos pelos neandertais – tornaram-se cada vez mais raros. Em 2013, testes de DNA no "cão de Altai", restos de um cão Paleolítico da Caverna Razboinichya (Montes Altai), ligaram este cão de 33.000 anos com a linhagem atual de Canis familiaris.[55]
Cruzamento
Na época dos últimos neandertais, aproximadamente 45 a 40 mil anos atrás, a análise genética sugere que houve um fluxo gênico dos neandertais para os humanos modernos de cerca de 10%, mas quase nenhum fluxo dos humanos modernos para os neandertais. Isso pode ser um artefato devido ao pequeno número de genomas neandertais tardios, ou porque os híbridos não eram viáveis em grupos neandertais, ou porque neandertais férteis estavam sendo absorvidos em grupos humanos modernos, mas não o contrário. Se o efeito foi real durante um período prolongado, teria aumentado o tamanho do pool gênico humano moderno e reduzido o dos neandertais já esparsos, contribuindo para reduzir seus números abaixo de uma população viável e, assim, para sua extinção.[56][57]
Endogamia
De acordo com um estudo de Rios et al, os padrões de parentesco entre os restos de neandertais recuperados sugerem que houve endogamia,[58] como acasalamentos entre meio-irmãos e/ou tio/tia e sobrinha/sobrinho.[59] Os pesquisadores hipotetizam que os neandertais podem ter ficado isolados em pequenos grupos durante condições climáticas severas, o que contribuiu para comportamentos de endogamia.[60] Devido à falta de diversidade genética, as populações neandertais teriam se tornado mais vulneráveis a mudanças climáticas, doenças e outros estressores, o que pode ter contribuído para sua extinção.[61][58] Um modelo semelhante à hipótese da endogamia pode ser visto entre os gorilas das terras baixas ameaçados de extinção. Suas populações são tão pequenas que causaram endogamia, tornando-os ainda mais vulneráveis à extinção.[62][63]
Mudanças climáticas
Sua extinção final coincide com o evento de Heinrich 4 [en], um período de intensa sazonalidade; eventos de Heinrich posteriores também estão associados a grandes mudanças culturais quando as populações humanas europeias entraram em colapso.[64][65] Essa mudança climática pode ter despovoado várias regiões de neandertais, como picos de frio anteriores, mas essas áreas foram repovoadas por humanos imigrantes, levando à extinção dos neandertais.[66] No sul da Ibéria, há evidências de que as populações neandertais declinaram durante o H4 e a proliferação associada de estepes desérticas dominadas por Artemisia.[67]
Os dados revelam que a mudança climática súbita, embora crucial localmente, teve um efeito limitado na população neandertal mundial. O cruzamento e a assimilação, que foram hipotetizados como causas na morte das populações neandertais europeias, são bem-sucedidos apenas para baixos níveis de competição por alimentos. Pesquisas futuras examinarão modelos de cruzamento, e a hibridização pode ser avaliada usando registros genômicos da última idade do gelo (Fu et al., 2016).[68]
Catástrofe natural
Vários pesquisadores argumentaram que a Erupção ignimbrítica da Campânia, uma erupção vulcânica perto de Nápoles, Itália, há cerca de 39 280 ± 110 anos (estimativa mais antiga ~37.000 anos), que eruptou cerca de 200 km3 (48 cu mi) de magma (500 km3 (120 cu mi) de volume bruto) contribuiu para a extinção dos neandertais.[69] O argumento foi desenvolvido por Golovanova et al.[70][71] A hipótese postula que, embora os neandertais tenham encontrado vários interglaciais durante 250.000 anos na Europa,[72] a incapacidade de adaptar seus métodos de caça causou sua extinção ao enfrentar a competição de Homo sapiens quando a Europa se transformou em uma estepe e semi-deserto esparsamente vegetado durante a última Idade do Gelo.[73] Estudos de camadas de sedimentos na Caverna de Mezmaiskaya sugerem uma severa redução do pólen de plantas.[71] O dano à vida vegetal teria levado a um declínio correspondente nos mamíferos herbívoros caçados pelos neandertais.[71][74][75]
Inversão magnética
Alguns pesquisadores sugeriram que a excursão geomagnética de Laschamps, uma breve inversão do campo magnético da Terra há cerca de 41.000 anos, pode ter contribuído para a extinção dos neandertais. A excursão causou um enfraquecimento da intensidade do campo magnético que protege a Terra da radiação nociva, incluindo a radiação ultravioleta, que é perigosa para os humanos. Argumenta-se que os humanos modernos podem ter sido menos suscetíveis aos efeitos prejudiciais da radiação do que os neandertais porque usavam ocre como protetor solar e usavam roupas costuradas, o que fornece mais proteção do que as roupas simples drapeadas dos neandertais.[76]
Ver também
- Lista de fósseis da evolução humana
- Extinções do final do Pleistoceno [en]
Referências
- ↑ Hortolà, Policarp; Martínez-Navarro, Bienvenido (8 de maio de 2013). «The Quaternary megafaunal extinction and the fate of Neanderthals: An integrative working hypothesis» [A extinção da megafauna do Quaternário e o destino dos Neandertais: Uma hipótese de trabalho integrativa]. Quaternary International. East meets West: First settlements and human evolution in Eurasia. 295: 69–72. Bibcode:2013QuInt.295...69H. ISSN 1040-6182. doi:10.1016/j.quaint.2012.02.037
- ↑ a b Higham, T.; Douka, K.; Wood, R.; Ramsey, C. B.; Brock, F.; Basell, L.; Camps, M.; Arrizabalaga, A.; Baena, J.; Barroso-Ruíz, C.; C. Bergman; C. Boitard; P. Boscato; M. Caparrós; N.J. Conard; C. Draily; A. Froment; B. Galván; P. Gambassini; A. Garcia-Moreno; S. Grimaldi; P. Haesaerts; B. Holt; M.-J. Iriarte-Chiapusso; A. Jelinek; J.F. Jordá Pardo; J.-M. Maíllo-Fernández; A. Marom; J. Maroto; M. Menéndez; L. Metz; E. Morin; A. Moroni; F. Negrino; E. Panagopoulou; M. Peresani; S. Pirson; M. de la Rasilla; J. Riel-Salvatore; A. Ronchitelli; D. Santamaria; P. Semal; L. Slimak; J. Soler; N. Soler; A. Villaluenga; R. Pinhasi; R. Jacobi (2014). «The timing and spatiotemporal patterning of Neanderthal disappearance» [O momento e o padrão espaço-temporal do desaparecimento dos Neandertais]. Nature. 512 (7514): 306–309. Bibcode:2014Natur.512..306H. PMID 25143113. doi:10.1038/nature13621. hdl:1885/75138
 .
. Mostramos que o Musteriense [a tradição de fabricação de ferramentas dos Neandertais] terminou há 41 030–39 260 anos calibrados AP (com 95,4% de probabilidade) em toda a Europa. Também demonstramos que as indústrias arqueológicas 'transicionais' subsequentes, uma das quais foi ligada aos Neandertais (Chatelperronense), terminam em um período similar.
- ↑ Higham, T. (2011). «European Middle and Upper Palaeolithic radiocarbon dates are often older than they look: problems with previous dates and some remedies» [Datas de radiocarbono do Paleolítico Médio e Superior Europeu são frequentemente mais antigas do que parecem: problemas com datas anteriores e alguns remédios]. Antiquity. 85 (327): 235–249. doi:10.1017/s0003598x00067570.
Poucos eventos da pré-história europeia são mais importantes do que a transição de humanos antigos para modernos há cerca de 40 000 anos, um período que, infelizmente, está próximo do limite da datação por radiocarbono. Este artigo mostra que até 70% das datas de radiocarbono mais antigas na literatura podem ser jovens demais, devido à contaminação por carbono moderno.
- ↑ a b Galván, B.; Hernández, C. M.; Mallol, C.; Mercier, N.; Sistiaga, A.; Soler, V. (2014). «New evidence of early Neanderthal disappearance in the Iberian Peninsula» [Nova evidência do desaparecimento precoce dos Neandertais na Península Ibérica]. Journal of Human Evolution. 75: 16–27. Bibcode:2014JHumE..75...16G. PMID 25016565. doi:10.1016/j.jhevol.2014.06.002
- ↑ Agusti, J.; Rubio-Campillo, X. (2017). «Were Neanderthals responsible for their own extinction?» [Os Neandertais foram responsáveis por sua própria extinção?]. Quaternary International. 431: 232–237. Bibcode:2017QuInt.431..232A. doi:10.1016/j.quaint.2016.02.017
- ↑ Devièse, Thibaut; Abrams, Grégory; Hajdinjak, Mateja; Pirson, Stéphane; De Groote, Isabelle; Di Modica, Kévin; Toussaint, Michel; Fischer, Valentin; Comeskey, Dan; Spindler, Luke; Meyer, Matthias; Semal, Patrick; Higham, Tom (23 de março de 2021). «Reevaluating the timing of Neanderthal disappearance in Northwest Europe» [Reavaliando o momento do desaparecimento dos Neandertais no Noroeste da Europa]. Proceedings of the National Academy of Sciences of the United States of America. 118 (12). Bibcode:2021PNAS..11822466D. ISSN 0027-8424. PMC 7999949. PMID 33798098. doi:10.1073/pnas.2022466118
- ↑ a b Zilhão, J.; Anesin, D.; Aubry, T. (2017). «Precise dating of the Middle-to-Upper Paleolithic transition in Murcia (Spain) supports late Neandertal persistence in Iberia» [Datação precisa da transição do Paleolítico Médio para o Superior em Múrcia (Espanha) apoia a persistência tardia dos Neandertais na Ibéria]. Heliyon. 3 (1). Bibcode:2017Heliy...300435Z. PMC 5696381. PMID 29188235. doi:10.1016/j.heliyon.2017.e00435
- ↑ Morales, J. I; Cebrià, A.; Burguet-Coca, A. (2019). «The Middle-to-Upper Paleolithic transition occupations from Cova Foradada (Calafell, NE Iberia)» [As ocupações da transição do Paleolítico Médio para o Superior na Cova Foradada (Calafell, NE da Ibéria)]. PLOS ONE. 14 (5). Bibcode:2019PLoSO..1415832M. PMC 6522054. PMID 31095578. doi:10.1371/journal.pone.0215832
- ↑ Cortés-Sánches, M.; Jiménez-Espejo, F. J.; Simón-Vallejo, M. D.; Stringer, C. (2019). «An early Aurignacian arrival in southwestern Europe» [Uma chegada precoce do Aurinhacense no sudoeste da Europa]. Nature Ecology & Evolution. 3 (2): 207–212. Bibcode:2019NatEE...3..207C. PMID 30664696. doi:10.1038/s41559-018-0753-6. hdl:10630/34212
- ↑ Anderson, L.; Reynolds, N.; Teyssandier, N. (2019). «No reliable evidence for a very early Aurignacian in Southern Iberia» [Nenhuma evidência confiável para um Aurinhacense muito precoce no Sul da Ibéria]. Nature Ecology and Evolution. 3 (5): 713. Bibcode:2019NatEE...3..713A. PMID 30988496. doi:10.1038/s41559-019-0885-3
- ↑ de la Peña, P. (2019). «Dating on its own cannot resolve hominin occupation patterns» [A datação por si só não pode resolver os padrões de ocupação dos hominínios]. Nature Ecology and Evolution. 3 (5): 712. Bibcode:2019NatEE...3..712D. PMID 30988497. doi:10.1038/s41559-019-0886-2
- ↑ Cortés-Sánches, M.; Jiménez-Espejo, F. J.; Simón-Vallejo, M. D.; Stringer, C. (2019). «Reply to 'Dating on its own cannot resolve hominin occupation patterns' and 'No reliable evidence for a very early Aurignacian in Southern Iberia'» [Resposta a 'A datação por si só não pode resolver os padrões de ocupação dos hominínios' e 'Nenhuma evidência confiável para um Aurinhacense muito precoce no Sul da Ibéria']. Nature Ecology and Evolution. 3 (5): 714–715. Bibcode:2019NatEE...3..714C. PMID 30988498. doi:10.1038/s41559-019-0887-1
- ↑ Fontugne, M.; Reyss, J. L.; Ruis, C. B.; Lara, P. M. (1995). «The Mousterian site of Zafarraya (Granada, Spain): dating and implications on the palaeolithic peopling processes of Western Europe» [O sítio musterenho de Zafarraya (Granada, Espanha): datação e implicações nos processos de povoamento paleolítico da Europa Ocidental]. Comptes Rendus de l'Académie des Sciences. 321 (10): 931–937
- ↑ Finlayson, C.; Pacheco, F. G. (2006). «Late survival of Neanderthals at the southernmost extreme of Europe» [Sobrevivência tardia dos Neandertais no extremo sul da Europa]. Nature. 443 (7,113): 850–853. Bibcode:2006Natur.443..850F. PMID 16971951. doi:10.1038/nature05195
- ↑ Slimak, L.; Svendsen, J. I.; Mangerud, J.; Plisson, H. (2011). «Late Mousterian persistence near the Arctic Circle» [Persistência musterenha tardia próximo ao Círculo Polar Ártico]. Science. 332 (6031): 841–845. Bibcode:2011Sci...332..841S. JSTOR 29784275. PMID 21566192. doi:10.1126/science.1203866
- ↑ Zwyns, N. (2012). «Comment on Late Mousterian persistence near the Arctic Circle» [Comentário sobre a persistência musterenha tardia próximo ao Círculo Polar Ártico]. Science. 335 (6065): 167. Bibcode:2012Sci...335..167Z. PMID 22246757. doi:10.1126/science.1209908
- ↑ Pavlov, P.; Svendsen, J. I.; Indrelid, S. (2001). «Human presence in the European Arctic nearly 40,000 years ago» [Presença humana no Ártico europeu há quase 40 000 anos]. Nature. 413 (6,851): 64–67. Bibcode:2001Natur.413...64P. PMID 11544525. doi:10.1038/35092552
- ↑ Pinhasi, R.; Higham, T. F. G.; Golovanova, L. V.; Doronichev, V. B. (2011). «Revised age of late Neanderthal occupation and the end of the Middle Palaeolithic in the northern Caucasus» [Idade revisada da ocupação neandertal tardia e o fim do Paleolítico Médio no norte do Cáucaso]. Proceedings of the National Academy of Sciences. 108 (21): 8611–8616. Bibcode:2011PNAS..108.8611P. PMC 3102382. PMID 21555570. doi:10.1073/pnas.1018938108.
A data direta do fóssil (39 700±1 100 14C AP) está em boa concordância com a função de distribuição de probabilidade, indicando com alto nível de probabilidade que os Neandertais não sobreviveram na Gruta de Mezmaiskaya após 39 ka cal AP. [...] Isso desafia alegações anteriores de sobrevivência tardia dos Neandertais no norte do Cáucaso. [...] Nossos resultados confirmam a falta de fósseis de Neandertais confiavelmente datados com menos de ≈40 ka cal AP em qualquer outra região da Eurásia Ocidental, incluindo o Cáucaso.
- ↑ Slimak, L.; Zanolli, C.; Higham, T. (2022). «Modern human incursion into Neanderthal territories 54,000 years ago at Mandrin, France» [Incursão de humanos modernos em territórios neandertais há 54 000 anos em Mandrin, França]. Science Advances. 8 (6). Bibcode:2022SciA....8J9496S. PMC 8827661. PMID 35138885. doi:10.1126/sciadv.abj9496
- ↑ Hublin, J.-J.; Sirakov, N. (2020). «Initial Upper Palaeolithic Homo sapiens from Bacho Kiro Cave, Bulgaria» [Homo sapiens do Paleolítico Superior Inicial da Gruta Bacho Kiro, Bulgária]. Nature. 581 (7808): 299–302. Bibcode:2020Natur.581..299H. PMID 32433609. doi:10.1038/s41586-020-2259-z. hdl:11585/770553
- ↑ Benazzi, S.; Douka, K.; Fornai, C.; Bauer, C.C.; Kullmer, O.; Svoboda, J.Í.; Pap, I.; Mallegni, F.; Bayle, P.; Coquerelle, M.; Condemi, S.; Ronchitelli, A.; Harvati, K.; Weber, G.W. (2011). «Early dispersal of modern humans in Europe and implications for Neanderthal behaviour» [Dispersão precoce de humanos modernos na Europa e implicações para o comportamento neandertal]. Nature. 479 (7374): 525–528. Bibcode:2011Natur.479..525B. PMID 22048311. doi:10.1038/nature10617
- ↑ Higham, T.; Compton, T.; Stringer, C.; Jacobi, R.; Shapiro, B.; Trinkaus, E.; Chandler, B.; Gröning, F.; Collins, C.; Hillson, S.; o'Higgins, P.; Fitzgerald, C.; Fagan, M. (2011). «The earliest evidence for anatomically modern humans in northwestern Europe» [A evidência mais antiga de humanos anatomicamente modernos no noroeste da Europa]. Nature. 479 (7374): 521–524. Bibcode:2011Natur.479..521H. PMID 22048314. doi:10.1038/nature10484
- ↑ Harvati, K. (2019). «Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia» [Fósseis da Gruta de Apidima fornecem a evidência mais antiga de Homo sapiens na Eurásia]. Nature. 571 (7766): 500–504. Bibcode:2019Natur.571..500H. PMID 31292546. doi:10.1038/s41586-019-1376-z. hdl:10072/397334
- ↑ de Lumley, M.-A.; Guipert, G.; de Lumley, H.; Protopapa, N.; Pitsios, T. (2020). «Apidima 1 and Apidima 2: two anteneandertal skulls in the Peloponnese, Greece» [Apidima 1 e Apidima 2: dois crânios anteneandertais no Peloponeso, Grécia]. L'Anthropologie. 124 (1). doi:10.1016/j.anthro.2019.102743
- ↑ Ko, Kwang Hyun (2016). «Hominin interbreeding and the evolution of human variation» [Cruzamento entre hominínios e a evolução da variação humana]. Journal of Biological Research-Thessaloniki. 23 (17). PMC 4947341. PMID 27429943. doi:10.1186/s40709-016-0054-7
- ↑ Boule, Marcellin (1920). Les hommes fossiles [Os homens fósseis]. Paris: Masson
- ↑ Kennedy, Jonathan (2023). Pathogenesis: how Germs Made History [Patogênese: como os Germes Fizeram a História]. London, UK: Torva. pp. 42–43
- ↑ Houldcroft, Charlotte J.; Underdown, Simon J. (julho de 2016). «Neanderthal genomics suggests a pleistocene time frame for the first epidemiologic transition: Neanderthal Infectious Diseases Genetics» [A genômica neandertal sugere um período pleistocênico para a primeira transição epidemiológica: Genética de Doenças Infecciosas Neandertais]. American Journal of Physical Anthropology. 160 (3): 379–388. PMID 27063929. doi:10.1002/ajpa.22985
- ↑ a b Greenbaum, Gili; Getz, Wayne M.; Rosenberg, Noah A.; Feldman, Marcus W.; Hovers, Erella; Kolodny, Oren (1 de novembro de 2019). «Disease transmission and introgression can explain the long-lasting contact zone of modern humans and Neanderthals» [Transmissão de doenças e introgressão podem explicar a zona de contato duradoura entre humanos modernos e Neandertais]. Nature Communications. 10 (1): 5003. Bibcode:2019NatCo..10.5003G. ISSN 2041-1723. PMC 6825168. PMID 31676766. doi:10.1038/s41467-019-12862-7
- ↑ Underdown, S. (2008). «A potential role for transmissible spongiform encephalopathies in Neanderthal extinction» [Um papel potencial para as encefalopatias espongiformes transmissíveis na extinção dos Neandertais]. Medical Hypotheses. 71 (1): 4–7. PMID 18280671. doi:10.1016/j.mehy.2007.12.014
- ↑ Liberski, P. (2013). «Kuru: a journey back in time from Papua New Guinea to the Neanderthals' extinction» [Kuru: uma jornada de volta no tempo da Papua Nova Guiné à extinção dos Neandertais]. Pathogens. 2 (3): 472–505. PMC 4235695. PMID 25437203. doi:10.3390/pathogens2030472
- ↑ a b c Wynn, Thomas; Overmann, Karenleigh A; Coolidge, Frederick L (2016). «The false dichotomy: A refutation of the Neandertal indistinguishability claim» [A falsa dicotomia: Uma refutação da alegação de indistinguibilidade dos Neandertais]. Journal of Anthropological Sciences. 94 (94): 201–221. PMID 26708102. doi:10.4436/jass.94022
- ↑ Banks, William E.; d'Errico, Francesco; Peterson, A. Townsend; Kageyama, Masa; Sima, Adriana; Sánchez-Goñi, Maria-Fernanda (24 de dezembro de 2008). Harpending, Henry, ed. «Neanderthal Extinction by Competitive Exclusion» [Extinção Neandertal por Exclusão Competitiva]. PLOS ONE. 3 (12). Bibcode:2008PLoSO...3.3972B. ISSN 1932-6203. PMC 2600607. PMID 19107186. doi:10.1371/journal.pone.0003972
- ↑ «Homo neanderthalensis Brief Summary» [Homo neanderthalensis Resumo Breve]. EOL. Consultado em 30 de dezembro de 2025
- ↑ Peresani, M; Dallatorre, S; Astuti, P; Dal Colle, M; Ziggiotti, S; Peretto, C (2014). «Symbolic or utilitarian? Juggling interpretations of Neanderthal behavior: new inferences from the study of engraved stone surfaces» [Simbólico ou utilitário? Malabarismo de interpretações do comportamento neandertal: novas inferências do estudo de superfícies de pedra gravadas]. J Anthropol Sci. 92 (92): 233–55. PMID 25020018. doi:10.4436/JASS.92007
- ↑ Bruner, Emiliano (2004). «Geometric Morphometrics and Paleoneurology: Brain Shape Evolution in the Genus Homo» [Morfometria Geométrica e Paleoneurologia: Evolução da Forma do Cérebro no Gênero Homo]. Journal of Human Evolution. 47 (5): 279–303. Bibcode:2004JHumE..47..279B. PMID 15530349. doi:10.1016/j.jhevol.2004.03.009
- ↑ Bruner, Emiliano (2010). «Morphological Differences in the Parietal Lobes with the Human Genus: A Neurofunctional Perspective» [Diferenças Morfológicas nos Lobos Parietais dentro do Gênero Humano: Uma Perspectiva Neurofuncional]. Current Anthropology. 51 (1): S77–S88. doi:10.1086/650729
- ↑ Bruner, Emiliano; Manzi, Giorgio; Arsuaga, Juan Luis (2003). «Encephalization and Allometric Trajectories in the Genus Homo: Evidence from the Neandertal and Modern Lineages» [Encefalização e Trajetórias Alométricas no Gênero Homo: Evidências das Linhagens Neandertal e Moderna]. Proceedings of the National Academy of Sciences of the United States of America. 100 (26): 15335–15340. Bibcode:2003PNAS..10015335B. PMC 307568. PMID 14673084. doi:10.1073/pnas.2536671100
- ↑ Hublin, Jean-Jacques; Neubauer, Simon; Gunz, Philipp (2015). «Brain Ontogeny and Life History in Pleistocene Hominins» [Ontogenia Cerebral e História de Vida em Hominínios do Pleistoceno]. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1663): 1–11. PMC 4305163. PMID 25602066. doi:10.1098/rstb.2014.0062
- ↑ Weaver, Anne (2005). «Reciprocal Evolution of the Cerebellum and Neocortex in Fossil Humans» [Evolução Recíproca do Cerebelo e Neocórtex em Humanos Fósseis]. Proceedings of the National Academy of Sciences of the United States of America. 102 (10): 3576–3580. Bibcode:2005PNAS..102.3576W. PMC 553338. PMID 15731345. doi:10.1073/pnas.0500692102
- ↑ Orban, Guy A; Caruana, Fausto (2014). «The Neural Basis of Human Tool Use» [A Base Neural do Uso de Ferramentas Humanas]. Frontiers in Psychology. 5: 310. PMC 3988392. PMID 24782809. doi:10.3389/fpsyg.2014.00310
- ↑ Bruner, Emiliano; Iriki, Atsushi (2016). «Extending Mind, Visuospatial Integration, and the Evolution of the Parietal Lobes in the Human Genus» [Estendendo a Mente, Integração Visuoespacial e a Evolução dos Lobos Parietais no Gênero Humano]. Quaternary International. 405: 98–110. Bibcode:2016QuInt.405...98B. doi:10.1016/j.quaint.2015.05.019
- ↑ Piazza, Manuela; Izard, Véronique (2009). «How Humans Count: Numerosity and the Parietal Cortex» [Como os Humanos Contam: Numerosidade e o Córtex Parietal]. Neuroscientist. 15 (3): 261–273. PMID 19436075. doi:10.1177/1073858409333073
- ↑ Vandervert, Larry R; Schimpf, Paul H; Liu, Hesheng (2007). «How Working Memory and the Cerebellum Collaborate to Produce Creativity and Innovation» [Como a Memória de Trabalho e o Cerebelo Colaboram para Produzir Criatividade e Inovação]. Creativity Research Journal. 19 (1): 1–18. doi:10.1080/10400410709336873
- ↑ Balsters, Joshua H; Whelan, Christopher D; Robertson, Ian H; Ramnani, Narender (2013). «Cerebellum and Cognition: Evidence for the Encoding of Higher Order Rules» [Cerebelo e Cognição: Evidências para a Codificação de Regras de Ordem Superior]. Cerebral Cortex. 23 (6): 1433–1443. PMID 22617850. doi:10.1093/cercor/bhs127. hdl:2262/72974
- ↑ Diamond, Jared (1992). The Third Chimpanzee: The Evolution and Future of the Human Animal [O Terceiro Chimpanzé: A Evolução e o Futuro do Animal Humano]. New York: Harper Collins. p. 45
- ↑ Kuhn, Steven L.; Stiner, Mary C. (2006). «What's a Mother to Do? The Division of Labor among Neandertals and Modern Humans in Eurasia» [O que uma Mãe Deve Fazer? A Divisão do Trabalho entre Neandertais e Humanos Modernos na Eurásia]. Current Anthropology. 47 (6): 953–981. doi:10.1086/507197
- ↑ Steudel-Numbers, Karen L; Tilkens, Michael J (2004). «The Effect of Lower Limb Length on the Energetic Cost of Locomotion: Implications for Fossil Hominins» [O Efeito do Comprimento dos Membros Inferiores no Custo Energético da Locomoção: Implicações para Hominínios Fósseis]. Journal of Human Evolution. 47 (1): 95–109. Bibcode:2004JHumE..47...95S. PMID 15288526. doi:10.1016/j.jhevol.2004.06.002
- ↑ Hora, M; Sládek, V (2014). «Influence of lower limb configuration on walking cost in Late Pleistocene humans» [Influência da configuração dos membros inferiores no custo de caminhada em humanos do Pleistoceno Superior]. Journal of Human Evolution. 67: 19–32. Bibcode:2014JHumE..67...19H. PMID 24485350. doi:10.1016/j.jhevol.2013.09.011
- ↑ Lewin, Roger (27 de abril de 1991). «Science: Neanderthals puzzle the anthropologists» [Ciência: Neandertais intrigam os antropólogos]. New Scientist. Consultado em 30 de dezembro de 2025
- ↑ Shipman, P (2012). «Dog domestication may have helped humans thrive while Neandertals declined» [A domesticação do cão pode ter ajudado os humanos a prosperarem enquanto os Neandertais declinavam]. American Scientist. 100 (3): 198. doi:10.1511/2012.96.198
- ↑ Ovodov, ND; Crockford, SJ; Kuzmin, YV; Higham, TFG; Hodgins, GWL (2011). «A 33,000-Year-Old Incipient Dog from the Altai Mountains of Siberia: Evidence of the Earliest Domestication Disrupted by the Last Glacial Maximum» [Um Cão Incipiente de 33.000 Anos das Montanhas Altai da Sibéria: Evidência da Domesticação Mais Antiga Interrompida pelo Último Máximo Glacial]. PLOS ONE. 6 (7). Bibcode:2011PLoSO...622821O. PMC 3145761. PMID 21829526. doi:10.1371/journal.pone.0022821
- ↑ Germonpré, M.; Sablin, M.V.; Stevens, R.E.; Hedges, R.E.M.; Hofreiter, M.; Stiller, M.; Jaenicke-Desprese, V. (2009). «Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: osteometry, ancient DNA and stable isotopes» [Cães e lobos fósseis de sítios paleolíticos na Bélgica, Ucrânia e Rússia: osteometria, DNA antigo e isótopos estáveis]. Journal of Archaeological Science. 36 (2): 473–490. Bibcode:2009JArSc..36..473G. doi:10.1016/j.jas.2008.09.033
- ↑ Fa, John E.; Stewart, John R.; Lloveras, Lluís; Vargas, J. Mario (2013). «Rabbits and hominin survival in Iberia» [Coelhos e sobrevivência de hominínios na Ibéria]. Journal of Human Evolution. 64 (4): 233–241. Bibcode:2013JHumE..64..233F. PMID 23422239. doi:10.1016/j.jhevol.2013.01.002
- ↑ Druzhkova, AS; Thalmann, O; Trifonov, VA; Leonard, JA; Vorobieva, NV (2013). «Ancient DNA Analysis Affirms the Canid from Altai as a Primitive Dog» [Análise de DNA Antigo Afirma que o Canídeo de Altai é um Cão Primitivo]. PLOS ONE. 8 (3). Bibcode:2013PLoSO...857754D. PMC 3590291. PMID 23483925. doi:10.1371/journal.pone.0057754
- ↑ Li, Liming (12 de julho de 2024). «Recurrent gene flow between Neanderthals and modern humans over the past 200,000 years» [Fluxo gênico recorrente entre Neandertais e humanos modernos nos últimos 200.000 anos]. Science. 185 (158): 8. Bibcode:2024Sci...385i1768L. PMID 38991054. doi:10.1126/science.adi1768
- ↑ Stringer, Chris; Crete, Lucia (2022). «Mapping Interactions of Homo neanderthalis and Homo sapiens From the Fossil and Genetic Records» [Mapeando Interações de Homo neanderthalensis e Homo sapiens a Partir dos Registros Fóssil e Genético]. PaleoAnthropology (2): 407. ISSN 1545-0031
- ↑ a b Ríos, L; Kivell, R.L; Lalueza-Fox, C (8 de fevereiro de 2019). «Skeletal Anomalies in The Neandertal Family of El Sidrón (Spain) Support A Role of Inbreeding in Neandertal Extinction» [Anomalias Esqueléticas na Família Neandertal de El Sidrón (Espanha) Apoiam um Papel da Endogamia na Extinção dos Neandertais]. Sci Rep. 9 (1): 1697. Bibcode:2019NatSR...9.1697R. PMC 6368597. PMID 30737446. doi:10.1038/s41598-019-38571-1
- ↑ Prüfer, K (5 de outubro de 2017). «A high-coverage Neandertal genome from Vindija Cave in Croatia.» [Um genoma neandertal de alta cobertura da Caverna Vindija, na Croácia.]. Science. 356 (6363): 655–658. Bibcode:2017Sci...358..655P. PMC 6185897. PMID 28982794. doi:10.1126/science.aao1887
- ↑ Vasesen, K; Scherjob, F; Hemerik, L; Verpoorte, A (27 de novembro de 2019). «Inbreeding, Allee effects and stochasticity might be sufficient to account for Neanderthal extinction» [Endogamia, efeitos Allee e estocasticidade podem ser suficientes para explicar a extinção dos Neandertais]. PLOS ONE. 14 (11). Bibcode:2019PLoSO..1425117V. PMC 6880983. PMID 31774843. doi:10.1371/journal.pone.0225117
- ↑ Vaesen, Krist (27 de novembro de 2019). «Inbreeding, Allee effects and stochasticity might be sufficient to account for Neanderthal extinction» [Endogamia, efeitos Allee e estocasticidade podem ser suficientes para explicar a extinção dos Neandertais]. PLOS ONE. 14 (11). Bibcode:2019PLoSO..1425117V. PMC 6880983. PMID 31774843. doi:10.1371/journal.pone.0225117
- ↑ Hedrick, Philip W.; Kalinowski, Steven T. (2000). «Inbreeding Depression in Conservation Biology» [Depressão Endogâmica em Biologia da Conservação]. Annual Review of Ecology and Systematics. 31 (1): 139–162. Bibcode:2000AnRES..31..139H. JSTOR 221728. doi:10.1146/annurev.ecolsys.31.1.139
- ↑ Macdonald, James (28 de janeiro de 2019). «When Endangered Wildlife Gets Inbred» [Quando a Vida Selvagem Ameaçada se Torna Endogâmica]. JSTOR Daily. Consultado em 30 de dezembro de 2025
- ↑ Bradtmöller, M.; Pastoors, A.; Weninger, B.; Weninger, G. (2012). «The repeated replacement model – Rapid climate change and population dynamics in Late Pleistocene Europe» [O modelo de substituição repetida – Mudança climática rápida e dinâmica populacional no Pleistoceno Superior da Europa]. Quaternary International. 247: 38–49. Bibcode:2012QuInt.247...38B. doi:10.1016/j.quaint.2010.10.015
- ↑ Wolf, D.; Kolb, T.; Alcaraz-Castaño, M.; Heinrich, S. (2018). «Climate deteriorations and Neanderthal demise in interior Iberia» [Deteriorações climáticas e desaparecimento dos Neandertais no interior da Ibéria]. Scientific Reports. 8 (1): 7048. Bibcode:2018NatSR...8.7048W. PMC 5935692. PMID 29728579. doi:10.1038/s41598-018-25343-6
- ↑ Staubwasser, M.; Drăgușin, V.; Onac, B. P. (2018). «Impact of climate change on the transition of Neanderthals to modern humans in Europe» [Impacto das mudanças climáticas na transição dos Neandertais para humanos modernos na Europa]. Proceedings of the National Academy of Sciences of the United States of America. 115 (37): 9116–9121. Bibcode:2018PNAS..115.9116S. PMC 6140518. PMID 30150388. doi:10.1073/pnas.1808647115
- ↑ d'Errico, Francesco; Sánchez Goñi, Marı́a Fernanda (abril de 2003). «Neandertal extinction and the millennial scale climatic variability of OIS 3» [Extinção neandertal e a variabilidade climática em escala milenar do OIS 3]. Quaternary Science Reviews. 22 (8–9): 769–788. Bibcode:2003QSRv...22..769D. doi:10.1016/S0277-3791(03)00009-X
- ↑ Timmermann, Axel (junho de 2020). «Quantifying the potential causes of Neanderthal extinction: Abrupt climate change versus competition and interbreeding» [Quantificando as causas potenciais da extinção dos Neandertais: Mudança climática abrupta versus competição e cruzamento]. Quaternary Science Reviews. 238 (106331). Bibcode:2020QSRv..23806331T. doi:10.1016/j.quascirev.2020.106331
- ↑ Fisher, Richard V.; Giovanni Orsi; Michael Ort; Grant Heiken (junho de 1993). «Mobility of a large-volume pyroclastic flow – emplacement of the Campanian ignimbrite, Italy» [Mobilidade de um fluxo piroclástico de grande volume – colocação da ignimbrita campaniana, Itália]. Journal of Volcanology and Geothermal Research. 56 (3): 205–220. Bibcode:1993JVGR...56..205F. doi:10.1016/0377-0273(93)90017-L. Consultado em 20 de setembro de 2008
- ↑ Golovanova, Liubov Vitaliena; Doronichev, Vladimir Borisovich; Cleghorn, Naomi Elansia; Koulkova, Marianna Alekseevna; Sapelko, Tatiana Valentinovna; Shackley, M. Steven (2010). «Significance of Ecological Factors in the Middle to Upper Paleolithic Transition» [Significado dos Fatores Ecológicos na Transição do Paleolítico Médio para o Superior]. Current Anthropology. 51 (5): 655. doi:10.1086/656185
- ↑ a b c Than, Ker (22 de setembro de 2010). «Volcanoes Killed Off Neanderthals, Study Suggests» [Vulcões Mataram os Neandertais, Sugere Estudo]. National Geographic. Consultado em 23 de setembro de 2010
- ↑ Gilligan, I (2007). «Neanderthal extinction and modern human behaviour: the role of climate change and clothing» [Extinção neandertal e comportamento humano moderno: o papel das mudanças climáticas e das roupas]. World Archaeology. 39 (4): 499–514. Bibcode:2007WoArc..39..499G. doi:10.1080/00438240701680492. hdl:1885/28508
- ↑ «Climate Change Killed Neandertals, Study Says» [Mudanças Climáticas Mataram os Neandertais, Diz Estudo]. National Geographic News. Consultado em 30 de dezembro de 2025
- ↑ «Volcanoes wiped out Neanderthals, new study suggests (ScienceDaily)» [Vulcões dizimaram os Neandertais, sugere novo estudo (ScienceDaily)]. University of Chicago Press Journals. 7 de outubro de 2010. Consultado em 7 de outubro de 2010
- ↑ «Neanderthal Apocalypse» [Apocalipse Neandertal]. ZDF Enterprises. 2015. Consultado em 30 de dezembro de 2025
- ↑ Mukhopadhyay, Agnit (16 de abril de 2025). «Wandering of the auroral oval 41,000 years ago» [Variação do oval auroral há 41.000 anos]. Science Advances. 11 (16). Bibcode:2025SciA...11.7275M. PMC 12002135. PMID 40238891. doi:10.1126/sciadv.adq7275