Enhydriodon
Enhydriodon
| |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Mioceno Superior ao Pleistoceno Inferior, 10,1–2 Ma | |||||||||||||||||
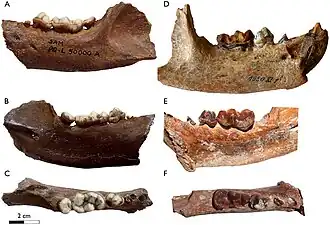 Dentições da mandíbula inferior de Enhydriodon hendeyi (A-C) e Enhydriodon africanus (D-F) | |||||||||||||||||
| Classificação científica | |||||||||||||||||
| |||||||||||||||||
| Espécie-tipo | |||||||||||||||||
| †Enhydriodon sivalensis Falconer, 1868 | |||||||||||||||||
| Outras espécies | |||||||||||||||||
| |||||||||||||||||
| Sinónimos | |||||||||||||||||
Sinonímia de gênero
Sinônimos de E. sivalensis
Sinônimos de E. ekecaman
| |||||||||||||||||
Enhydriodon é um gênero extinto de lontras conhecido da África, Paquistão e Índia, que viveu do final do Mioceno ao início do Pleistoceno. Contém nove espécies confirmadas, duas espécies debatidas e pelo menos algumas outras espécies não descritas da África. O nome do gênero significa "dente de lontra" em grego antigo e é uma referência à sua dentição, e não ao gênero Enhydra, que inclui a lontra-marinha moderna e seus dois parentes pré-históricos. Enhydriodon pertence à tribo Enhydriodontini (que também contém Sivaonyx e Vishnuonyx) na subfamília de lontras Lutrinae.
Os tamanhos e comprimentos exatos das espécies de Enhydriodon são desconhecidos, dada a falta de fósseis completos dela e da maioria das lutrinas fósseis relacionadas. Estima-se que as espécies do subcontinente indiano tivessem pesos semelhantes aos da lontra-marinha existente, mas estima-se que as espécies africanas fossem mais pesadas que as lutrinas existentes. Em particular, várias espécies como Enhydriodon kamuhangirei, E. dikikae e E. omoensis foram estimadas em mais de 100 kg. Dadas essas estimativas de peso, as três espécies provavelmente atingiram tamanhos comparáveis aos de ursos ou leões existentes, tornando-as os maiores mustelídeos conhecidos, embora a falta de espécimes completos torne as estimativas precisas impossíveis.
Sua dentição avançada é bem conhecida, seus carnassiais largos e bunodontes permitindo que a lutrina consumisse presas esmagando-as, em vez de cortá-las como a lontra-marinha moderna e ao contrário da maioria das outras lontras existentes. Como tal, é agrupado entre as lontras bunodontes, um termo categórico que se refere a lutrinas fósseis com carnassiais não laminares nos pré-molares ou molares do Mioceno ao Pleistoceno e à lontra-marinha do único gênero existente, Enhydra. Seus dentes I3 (ou terceiros incisivos) são semelhantes a caninos e muito maiores que seus outros incisivos (embora mais curtos que seus caninos), uma característica não vista em gêneros de lutrinas existentes e extintas. Hipotetiza-se que as espécies indianas de Enhydriodon eram semiaquáticas e consumiam bivalves, porque suas dentições bunodontes teriam permitido que consumissem invertebrados de casca dura. Não se sabe se as espécies africanas eram geralmente aquáticas, semiaquáticas ou terrestres, mas suas dietas potenciais adequadas para dentições bunodontes incluem bivalves, Siluriformes, répteis, ovos e carniça. E. omoensis da Etiópia, em particular, poderia ter sido um locomotor terrestre que, pelo menos semirregularmente, caçava ou se alimentava de presas terrestres com dietas de plantas com fixação de carbono C4, o que, se verdadeiro, torna seu comportamento diferente de qualquer lontra existente. Não se sabe se a espécie é uma exceção entre as espécies de lontras bunodontes africanas, mas foi sugerido que E. dikikae e Sivaonyx beyi também eram grandes lontras bunodontes terrestres da África.
O status taxonômico das espécies de Enhydriodon tem sido complicado por suas afinidades e semelhanças com outros gêneros de lutrinas bunodontes como Sivaonyx e Paludolutra até os dias de hoje, embora Paludolutra seja atualmente considerado um gênero distinto não proximamente relacionado a Enhydriodon. Atualmente, a tribo Enhydriodontini é considerada evolutivamente mais próxima do gênero moderno Enhydra do que qualquer outro gênero de lontra bunodonte conhecido que possa ter adquirido dentição bunodonte como resultado de evolução paralela, mas a extensão de sua relação próxima permanece não resolvida.
Taxonomia
Início da história
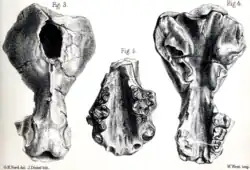
Enhydriodon foi erigido pela primeira vez em 1868 por Hugh Falconer, com base em vários crânios de Sivalik, na Índia, que ele atribuiu a Enhydriodon sivalensis.[1][2] Ele explicou que o nome científico, que significa "dente de lontra", deriva dos termos gregos antigos ἐνυδρίς (lontra) e ὀδούς (dente) e não é uma referência ao gênero Enhydra, que inclui a lontra-marinha moderna (Enhydra lutris). Segundo Falconer, os fósseis de Sivalik pertencentes a Enhydriodon sivalensis foram anteriormente classificados por Falconer e Proby Cautley sob o nome de gênero e espécie Amyxodon sivalensis em uma sinopse de 1835 dos gêneros fósseis de Sivalik que os dois paleontólogos encontraram, na qual o táxon fóssil foi considerado um carnívoro de uma família desconhecida, embora nenhum holótipo ou descrições diagnósticas tenham sido atribuídos a ele. Como resultado da renomeação, Amyxodon foi considerado um "nome morto" ou sinônimo de Enhydriodon, apesar de ser o nome de gênero mais antigo. Usando os espécimes disponíveis de nhydriodon. sivalensis, Falconer calculou que havia quatro pré-molares e molares na mandíbula superior da espécie (a fórmula dentária foi calculada como ), um a menos que no gênero existente Lutra [en], mas correspondendo à contagem total de Enhydra. Ele descreveu o carniceiro superior de Enhydriodon sivalensis como a característica mais única de sua mandíbula superior, sendo quase quadrado e seus lóbulos coronais desenvolvidos a partir de mamelões cônicos, ao contrário dos dois gêneros de lutrinas existentes.[1][3]

Durante os séculos XIX e XX, mais espécies de Enhydriodon, como E. campanii, foram introduzidas e mais gêneros de lutrinas com dentição bunodonte, como Sivaonyx e Vishnuonyx, foram descritos, criando uma história particularmente complicada para o gênero de lontra pré-histórica descrito mais cedo. Em 1931, Guy Pilgrim descreveu mais fósseis descobertos em Sivalik, incluindo uma nova espécie chamada E. falconeri. Ele também implicou que Enhydriodon e Sivaonyx, apesar de suas semelhanças, eram diferenciados pela estrutura do quarto pré-molar maxilar (P4) e pela aparente ausência do pré-molar superior anterior (P1), que se presume ser refletida também na mandíbula inferior (ambos são debatidos até hoje).[4] No mesmo ano em que E. falconeri foi descrito, Ernst Stromer [en] descreveu E. africanus do final do Plioceno, cujos dentes fósseis foram localizados na África do Sul e foi a primeira espécie descrita do continente africano.[5]
Relações percebidas com Paludolutra e Enhydra
Em 1976, Charles Repenning [en] levantou a ideia de que Enhydriodon estava relacionado ao gênero existente Enhydra, devido às supostas espécies do primeiro serem um "ramo" evolutivo de "lontras comedoras de caranguejo" na Itália, Espanha e Califórnia, levando eventualmente à lontra-marinha moderna.[6] Ele introduziu corretamente a ideia de que Enhydra estava relacionado a Enhydriodon, dadas suas dentições bunodontes, mas o suposto "ramo" europeu do gênero Enhydriodon foi posteriormente reclassificado por Johannes Hürzeler e Burkart Engesser para o novo gênero Paludolutra em 1976, embora tenha permanecido relativamente obscuro no registro paleontológico até que pesquisas posteriores revisassem seu estado taxonômico.[7][8]
As taxonomias de espécies e gêneros individuais de lutrinas continuaram a ser revisadas no século XXI, à medida que mais espécies de lontras pré-históricas eram descritas, enquanto os paleontólogos revisavam continuamente as espécies de lutrinas bunodontes fósseis para diferentes gêneros. Paludolutra foi originalmente reclassificado como um subgênero de Enhydriodon por Gerard F. Willemsen em 1992.[5] No entanto, em janeiro de 2005, Martin Pickford [en] e colegas diagnosticaram Paludolutra como um sinônimo de Sivaonyx com base no diagnóstico de Pilgrim deste último, rejeitando a sinonímia de Willemsen de Paludolutra com Enhydriodon. Além disso, eles erigiram uma espécie de Enhydriodon chamada Enhydriodon hendeyi da localidade-tipo de Langebaanweg [en], África do Sul, que data do Plioceno inferior e foi nomeada em homenagem ao paleontólogo Quinton B. Hendey, que, segundo eles, descreveu os primeiros espécimes conhecidos que desde então foram atribuídos à espécie.[9] Em dezembro do mesmo ano, Jorge Morales e Pickford, em vez disso, descreveram Paludolutra como um gênero distinto que poderia estar relacionado a Sivaonyx com base em convergências de dentição.[8] Em 2007, os dois paleontólogos reafirmaram que a morfologia dentária de Paludolutra era distinta o suficiente para ser reclassificada como um gênero com base na diferenciação genérica completa, sugerindo que as espécies Paludolutra campanii, P. lluecai e P. maremmana não seriam mais classificadas sob Enhydriodon com base em Paludolutra ser um subgênero.[10][11]
Revisões modernas de espécies africanas
Em 2003, Lars Werdelin [en] erigiu a espécie Enhydriodon ekecaman do sítio paleontológico de Kanapoi [en], na bacia do Turkana [en], no Quênia (início do Plioceno, ca. 5,2-4,0 Ma), descrevendo-a como um dos primeiros membros da linhagem africana de Enhydriodon. A espécie foi nomeada a partir do termo da língua Turkana "ekecaman", que significa "pescador", porque ele sugeriu que peixes podem ter sido uma dieta para a espécie. Ele também declarou a espécie "E. pattersoni ", descrita por R. J. G. Savage [en] em 1978, como nomen nudum de E. ekecaman, uma vez que nenhum espécime-tipo ou diagnóstico válido foi designado para ela, uma visão apoiada por Morales e Pickford em dezembro de 2005.[12][8]
Enhydriodon africanus, E. ekecaman e E. hendeyi foram reclassificados para Sivaonyx por Pickford e Morales em dezembro de 2005, onde eles adicionalmente descreveram uma nova espécie chamada Sivaonyx kamuhangirei.[8] A reclassificação de lontras bunodontes fósseis africanas para Sivaonyx gerou um debate contínuo sobre a praticidade das diferenças entre Enhydriodon e Sivaonyx, com alguns pesquisadores alegando neutralidade devido a focos preferenciais na pesquisa das espécies individuais em vez de suas colocações de gênero. Em 2022, as quatro espécies foram eventualmente reclassificadas para Enhydriodon em um artigo de pesquisa de Camille Grohé et al. E. soriae também foi inicialmente classificada em Sivaonyx, mas foi eventualmente atribuída a Enhydriodon, embora sua colocação de gênero permaneça disputada.[13][14] Em 2005, Morales e Pickford classificaram Enhydriodon na tribo recém-criada Enhydriodontini, que eles descreveram como abrigando gêneros de lontras bunodontes extintas de Sivalik e da África, incluindo Vishnuonyx, Sivaonyx e Paludolutra. Em 2007, Pickford sinonimizou a espécie "E. aethiopicus ", anteriormente descrita por Denis Geraads et al. em 2004, com Pseudocivetta ingens, um membro extinto da família Viverridae.[10] Em 2017, Enhydra foi explicitamente excluída da tribo Enhydriodontini, apesar de suas semelhanças, e Paludolutra foi reclassificada como um táxon irmão da tribo.[8][11]
Em 2011, Denis Geraads e colegas descreveram E. dikikae com base em seus restos de um crânio parcial e fêmures no Baixo Awash de Dikika, Etiópia, com a localidade datando do Plioceno médio. Foi descrito como tendo um crânio notavelmente mais pesado (embora quebrado) do que outras espécies de Enhydriodon ou a lontra-marinha moderna. O nome da espécie foi baseado diretamente no sítio de Dikika.[15] Foi considerada a maior espécie de Enhydriodon até que outra espécie também da Etiópia, E. omoensis, foi descrita do Baixo Vale do Omo em 2022, datando do final do Plioceno até o limite Plio-Pleistoceno. Semelhante a E. dikikae, o nome da espécie foi derivado diretamente do sítio em que foi recuperado.[14] Em uma conferência de setembro de 2022 por Alberto Valenciano, Morales e Pickford (no mesmo mês do artigo de pesquisa sobre E. omoensis), no entanto, eles se referiram a certas espécies de lutrinas anteriormente reclassificadas para Enhydriodon como Sivaonyx, a saber, S. hendeyi e S. africana.[16]
Classificação

Enhydriodon pertence à tribo Enhydriodontini na subfamília Lutrinae, que apareceu pela primeira vez na Eurásia e na África durante o final do Mioceno.[8] É talvez a lontra pré-histórica mais conhecida, dada a sua longa história taxonômica e por ser uma fonte primária de comparações com outros gêneros de lontras bunodontes. Geralmente, pensa-se que Enhydriodon foi o resultado de uma tendência do Mioceno-Pleistoceno que deu às lutrinas pré-históricas dentes bunodontes e grandes tamanhos em comparação com seus parentes existentes. É classificado como um membro do grupo de lontras bunodontes, um termo categórico comumente usado por pesquisadores que também inclui Sivaonyx, Paludolutra, Vishnuonyx, Torolutra, Enhydritherium [en], Djourabus, Paralutra, Tyrrhenolutra, Siamogale [en] e Enhydra.[11][17][18] As lontras bunodontes são definidas como mustelídeos de grande a muito grande porte da América do Norte, Eurásia e África que tinham uma dentição robusta em comparação com a maioria das lontras existentes, geralmente permitindo que eles predatem criaturas com armadura dura.[a][13][17] Apesar de compartilharem a característica de dentição bunodonte, existem pelo menos vários clados de lutrinas pertencentes a esta categoria, em vez de um só, então o termo "lontras bunodontes", portanto, é categórico em vez de taxonômico e abrange lutrinas durante os mesmos períodos com dentições semelhantes, em vez de um que defina diretamente seu estado taxonômico.[11]
O cladograma a seguir de Xiaoming Wang et al. em 2018 define algumas das seguintes espécies e gêneros de lontras existentes e extintas dentro da subfamília Lutrinae, com base em um consenso de maioria de 50% (os gêneros de lontras bunodontes estão em negrito a partir de "Paralutra jaegeri"):[11]
| Lutrinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Como mostrado na filogenia acima, Enhydriodon compartilhava uma morfologia mais próxima com seus outros parentes extintos e com Enhydra do que com as outras lutrinas existentes que não possuem dentes carnassiais bunodontes (Lutra aonychoides foi descrito como não sendo relacionado a Lutra). Embora a árvore de consenso de maioria exiba uma relação morfológica próxima entre Enhydriodon e Enhydra, os autores da árvore de consenso também criaram uma árvore de inferência bayesiana propondo que Enhydra é um clado isolado, separado dos membros típicos de Enhydriodontini ("Paralutra" jaegeri foi proposto como um clado isolado de Siamogale também). Independentemente disso, eles argumentaram que Enhydra está mais próximo do clado composto por Enhydriodon, Sivaonyx e Vishnuonyx do que qualquer outro gênero de lontra bunodonte. Os pesquisadores explicaram que a aquisição da dentição bunodonte ocorreu pelo menos três vezes na evolução das lutrinas, refletida pelos clados da árvore filogenética: em Sivaonyx-Enhydriodon-Enhydra, em Paludolutra-Enhydritherium e em Siamogale.[11] Lontras não-bunodontes provavelmente se ramificaram separadamente das lontras bunodontes durante ou antes do Plioceno, mas seus registros fósseis pobres e restrição a depósitos do Plio-Pleistoceno, em comparação, deixam pouco entendimento em suas filogenias evolutivas.[20]
Descrição
Crânio
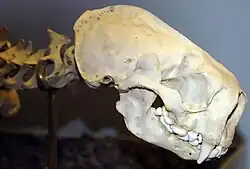
Atualmente, existem apenas dois crânios parciais conhecidos que são atribuídos a Enhydriodon: um de E. sivalensis de Sivalik e o outro de E. dikikae do vale do Awash. Atualmente, não se sabe se as características dos crânios de qualquer uma das espécies são bem representativas de outras espécies de Enhydriodon, mas os crânios conhecidos de E. dikikae e E. sivalensis têm características um tanto diferentes entre si.[15]
O crânio de E. sivalensis, identificado como pertencente a um indivíduo totalmente crescido, está relativamente bem preservado, com cristas temporais, ossos frontais, maxilas, pré-maxilas, ossos nasais, focinho e partes do osso palatino identificáveis. No entanto, também sofreu desgaste e está ligeiramente torcido no sentido horário. Mais notavelmente, a arcada dentária está completa, embora o M1 esquerdo e o I1 esquerdo estejam ambos ausentes e a maioria dos dentes esteja quebrada a partir de suas coroas. Possui uma caixa craniana grande, um focinho largo e curto e uma grande abertura nasal. Contornos das órbitas ao redor dos frontais do crânio também podem ser identificados.[2]
O crânio quebrado pertencente a E. dikikae contém um focinho curto e não prognático, partes das órbitas, um arco dentário superior quase completo ao qual faltam ambos os I1s e um I2 direito, e parte da mandíbula inferior. O focinho no crânio de E. dikikae é curto, com uma pequena borda orbital anterior posicionada logo acima do lado posterior do canino. A parte frontal do focinho é identificada como curta, sendo, portanto, comparável ao focinho de Enhydra. Embora a evolução de lontras bunodontes como Enhydriodon não seja clara, propõe-se que o focinho curto de E. dikikae e o tamanho muito grande de seus caninos tornam a espécie claramente diferente na aparência e mais derivada evolutivamente (ou evolutivamente recente) do que E. sivalensis.[15]
Dentição
A dentição de Enhydriodon é bem definida por seus carnassiais bunodontes extremamente largos nos molares e pré-molares, semelhantes aos da lontra-marinha moderna. As diferenças entre as espécies de Enhydriodon e Sivaonyx são geralmente atribuídas à dentição, então os dentes pré-molares ou molares fósseis são examinados para discernir os dois gêneros de lontras bunodontes. As diferenças genéricas (hipocone do P4 maior, cúspides cônicas pós-protocone e aparente falta de pré-molares superiores anteriores para Enhydriodon) por meio de medições dentárias têm sido difíceis de provar devido à natureza fragmentária dos fósseis e às inconsistências relativas das medições/dimensões dos dentes por espécie.[10][15] A reclassificação de todas as espécies "africanas de Sivaonyx", exceto S. beyi, para Enhydriodon em 2022 foi atribuída a "[um] metaconídeo mais alto que o protoconídeo no M1, presença de uma incisura carnassial e uma ou mais cúspides entre o protocone e o hipocone no P4, e/ou expansão distolingual no M1."[14]
Sugere-se que Enhydriodon, como o gênero de aparição mais tardia, tenha a dentição mais bunodonte da tribo Enhydriodontini, que inclui o Vishnuonyx de aparição mais antiga e depois o Sivaonyx. A dentição de Enhydriodon sugere uma supressão quase total das funções carnassiais em favor do esmagamento como função predominante. O I3 (ou terceiro incisivo superior) de Enhydriodon é muito maior que seu I1 (menor incisivo) e I2, parecendo maior e mais semelhante a um canino em comparação com Paludolutra e Enhydra. Em comparação com outros gêneros de lutrinas bunodontes onde o incisivo superior é conhecido, seus terceiros incisivos são apenas marginalmente maiores que seus primeiros e segundos incisivos.[10] O I1 direito de um crânio de Enhydriodon sivalensis, por exemplo, mede 3 mm de diâmetro anteroposterior (APD) e 4,5 mm de diâmetro transverso (TD). O I2 direito do crânio mede 5,2 mm de APD e 5,5 mm de TD. Em comparação, o I3 direito é o maior incisivo do holótipo, com medidas de 10,5 mm de APD e 8 mm de TD (os caninos são maiores que os incisivos, medindo 17,1 mm de APD e 13,8 mm de TD).[2] A característica do I3 grande também se aplica a E. dikikae, que foi descrito após a descrição geral de Pickford do gênero Enhydriodon como tendo um I3 muito maior que I1 - I2 e sendo de formato mais cônico. O dente I3 do DIK-56 mede 12,4 mm de largura mesiodistal (MD) e 11,6 mm de largura bucolingual (BL), em comparação com as medidas de seu I2 de 5,5 mm de MD e 9,7 mm de BL. Como Enhydriodon sivalensis, o I3 é mais curto que os caninos, com o C1 medindo 16,9 mm de MD mais 15 mm de BL e o C1 medindo 19,5 mm de MD e 15,3 mm de BL.[15]
Membros
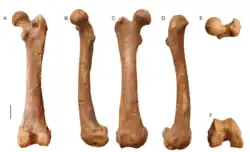
_hendeyi_Aonyx_capensis_femora.jpg)
Restos pós-cranianos de lontras bunodontes, incluindo Enhydriodon, são escassos, deixando pouca informação sobre a anatomia geral de muitos gêneros. As únicas espécies conhecidas de Enhydriodon com restos pós-cranianos são E. hendeyi, E. dikikae e E. omoensis.[13]
Os restos fósseis de Enhydriodon hendeyi incluem um úmero fragmentário, uma ulna, dois fêmures e um astrágalo (também conhecido como osso do talo). Os fêmures de E. hendeyi são menores que os de Sivaonyx beyi e Enhydriodon dikikae, mas também maiores que os de Enydritherium, Satherium e da lontra-africana-sem-garras existente. O astrágalo é semelhante ao de E. omoensis, mas difere pela cabeça menor e pelo pescoço mais fino, bem como por uma projeção distal maior do tubérculo do osso. A tróclea do osso do talo (superfície sulcada que forma as articulações dos ossos) é rasa e mediolateralmente larga, enquanto seu tubérculo é projetado no centro aproximado, ambos produzindo um sulco robusto e profundo dos tendões dos músculos da flexão plantar [en] para extensão do pé no tornozelo, em comparação com a lontra-africana-sem-garras.[13]
Os restos pós-cranianos de E. dikikae são conhecidos pelo fêmur esquerdo proximal (parte superior), fêmur direito distal (parte inferior) e um úmero. O fêmur esquerdo proximal é conhecido por um grande tubérculo ao longo da área posterior do pescoço, trocanter menor [en] de alinhamento médio e uma posição semelhante do côndilo medial do fêmur. Os espécimes de fêmur indicam tamanhos gerais grandes dos fêmures de E. dikikae em comparação até mesmo com as maiores lontras existentes. O úmero (completo, mas desgastado e rachado) é muito mais longo e ligeiramente mais robusto em comparação com o de Sivaonyx beyi, e sua tuberosidade deltoide [en] é bem formada. A crista supracondilar lateral do úmero é mais longa em comparação com S. beyi, enquanto o epicôndilo medial do úmero não é tão proeminente em tamanho. A fossa do olécrano [en] é pequena e mais circular em comparação com S. beyi.[15]
Enhydriodon omoensis é representado apenas por um único fêmur esquerdo completo, que tem um pescoço curto e uma cabeça redonda orientada em uma direção proximal (próxima ao centro) em vez de uma direção medial (no centro), sendo a primeira deslocada a 40° em relação à longitude da seção da diáfise do osso. A largura lateromedial da epífise é estreita. O fêmur também tem uma grande cabeça femoral [en] localizada na cabeça ventromedial (alinhando-se à parte inferior média dela), um trocânter maior [en] que se dobra na parte de trás e é mais baixo que a cabeça femoral, uma fossa trocantérica [en] curta e profunda e um trocânter menor forte que é centrado mais no meio do que na parte ventral (ou inferior) e é, portanto, visível em uma visão posterior. O côndilo medial do fêmur é maior que o côndilo lateral do mesmo osso. A fossa intercondilar do fêmur é retangular e larga.[14]
Massa Corporal

Algumas espécies de Enhydriodon, particularmente algumas que residiram na África, são os maiores mustelídeos conhecidos [en] que já existiram com base em estimativas de peso, mas seus tamanhos e pesos precisos permanecem desconhecidos, dada a falta de espécimes completos em seus registros fósseis. Algumas espécies como E. latipes(?) são pouco estudadas em comparação com outras e, portanto, carecem de estimativas confirmadas de tamanho ou peso.[21] Geralmente, estima-se que algumas espécies de Enhydriodon tenham um peso semelhante ao de lontras de grande porte modernas, enquanto outras são estimadas como muito maiores (deve-se notar também que as estimativas de peso são feitas mais frequentemente para lontras bunodontes como Enhydriodon do que as estimativas de tamanho, embora comparações de tamanho com animais modernos possam ser referenciadas).[14]
As duas espécies de Enhydriodon nativas do subcontinente indiano tinham estimativas de peso modestas, comparáveis à maioria dos outros gêneros de lontras bunodontes, bem como aos gêneros de lontras existentes. O memorial de Falconer de 1868 descreveu E. sivalensis como uma lutrina do tamanho de uma pantera.[1] Em 1932, Pilgrim diagnosticou E. falconeri como sendo menor que E. sivalensis, embora não tenha oferecido estimativas de tamanho ou peso para ela.[4] Em 2007, Pickford estimou que E. sivalensis era a maior lontra pré-histórica da Índia, variando de 22 kg no mínimo a 25 kg no máximo de peso corporal, com o crânio possivelmente do tamanho de um lobo. Ele também estimou o corpo de E. falconeri, com base nas dimensões de seus dentes M1 inferiores, como sendo semelhante ao da çontra‑do‑cabo (Aonyx capensis), com uma média de 16 kg.[10]
As espécies de Enhydriodon da África são estimadas como algumas das maiores espécies de lontras que já existiram, refletindo a tendência do Mioceno-Pleistoceno de as lontras bunodontes crescerem mais do que suas primas não-bunodontes. Pickford descreveu E. kamuhangirei do vale do Rift Ocidental, Uganda (na época Sivaonyx kamuhangirei), como podendo exceder 100 kg de peso, tornando-a a maior lontra pré-histórica conhecida na época, embora tenha mencionado que as lontras fósseis não descritas na Etiópia (provavelmente classificadas mais tarde sob E. dikikae e/ou E. omoensis) poderiam ter sido maiores.[10] E. hendeyi (então Sivaonyx hendeyi) foi estimado como do tamanho de um lobo e com cerca de 40 kg, enquanto E. africanus e E. ekecaman são considerados de tamanhos semelhantes.[22][14] E. dikikae da Etiópia foi estimado em um peso mínimo de 100 kg e máximo de 200 kg (sendo este último mencionado como mais provável), com seu holótipo sugerindo um tamanho semelhante ao de um urso. Comparado com a maioria das outras espécies de Enhydriodon ou Enhydra, tinha um comprimento de crânio estimado em cerca de 25 cm.[15] E. omoensis foi posteriormente estimado em mais de 200 kg de peso, tornando-o mais pesado que E. dikikae e os leões modernos. Segundo Grohé et al., E. omoensis era potencialmente "do tamanho de um leão", tornando-se a maior espécie de mustelídeo que já existiu.[14]
Paleobiologia
.jpg)
Como os gêneros de lontras bunodontes fósseis, incluindo Enhydriodon, geralmente carecem de espécimes completos e elementos pós-cranianos, sua locomoção e nichos ecológicos permanecem incertos. Uma teoria comum sobre as espécies do subcontinente indiano de Enhydriodon é que, com base em sua dentição robusta e bunodonte semelhante à de Enhydra, Enhydriodon falconeri e E. sivalensis eram ambos especializados em comer mariscos com frequência.[5] Essa afirmação foi feita pela primeira vez por Willemsen a partir de analogias da dieta de Enhydra (abalones e bivalves marinhos) e Aonyx (caranguejos de água doce), mas há pouca evidência paleontológica para apoiar diretamente essa afirmação. Independentemente disso, sugere-se que o esmalte espesso na dentição posterior das espécies indianas de Enhydriodon as torna mais moluscívoras do que cancrivoras (em contraste, sugere-se que as espécies indianas de Sivaonyx tenham combinado funções de corte dos carnassiais com coroas bunodontes gerais para predar mais crustáceos, embora os bivalves pudessem ter sido presas secundárias). A possibilidade de Enhydriodon predar bivalves é apoiada pela presença de gêneros de bivalves de água doce fossilizados, Parreysia e Lamellidens, nos mesmos locais que eles, ambos comuns em toda a coluna sedimentar de Sivalik, que abrange de 15 a 2 Ma, coincidindo com a presença da tribo Enhydriodontini no subcontinente indiano (Índia e Paquistão).[10]
Sugere-se que as espécies maiores de Enhydriodon no continente africano tenham predado uma variedade maior de alimentos, além de suas presas primárias, incluindo presas mais macias, apesar de suas dentições bunodontes, tornando suas dietas potenciais distintas das de seus homólogos do subcontinente indiano. Um tipo de presa sugerido eram peixes grandes com coberturas externas duras, como Siluriformes.[23] Vários gêneros de bagres estavam presentes na África desde suas primeiras aparições durante o final do Mioceno, coincidindo com a presença de Enhydriodon, incluindo os gêneros existentes Clarotes [en], Bagrus [en], Auchenoglanis [en] e Chrysichthys e o gênero extinto Nkondobagrus.[24] Em contraste com os bagres de movimento lento e abundantes, os caranguejos na África foram excluídos como presas potenciais para as espécies africanas de Enhydriodon, dada a falta de caranguejos fossilizados em Dikika, a improbabilidade de as biomassas de caranguejos sustentarem populações de grandes lontras e a aparente incompatibilidade com a dentição de esmalte. Peixes de natação rápida provavelmente não eram fontes de alimento regulares, devido à dentição especializada para esmagar alimentos duros, além de grandes animais provavelmente não terem a capacidade de capturar presas rápidas. Outras presas com armadura, como crocodilos juvenis, tartarugas e ovos de avestruz, também foram sugeridas como presas de E. dikikae.[15]

Os fêmures e restos dentários de Enhydriodon africano poderiam possivelmente sugerir um estilo de vida semiaquático e terrestre, o que significa que poderia ter comido tanto presas aquáticas quanto terrestres. As especulações sobre o estilo de vida de Enhydriodon, no entanto, têm sido contraditórias entre si, então não há, portanto, um consenso majoritário sobre isso. Em 2008, especulou-se que as espécies africanas menores de Enhydriodon, com base em seus tamanhos de fêmur menores, eram mais generalistas de locomoção, semelhantes à maioria dos mustelídeos, enquanto as espécies maiores eram totalmente aquáticas, já que suas estruturas de fêmur compartilhavam semelhanças com Enhydra. No entanto, as extremidades proximais dos fêmures de Omo e Hadar apontavam para uma natureza mais aquática do que a maioria das lutrinas, enquanto seus comprimentos relativos se assemelhavam aos de mustelídeos generalistas terrestres, incluindo lontras semiaquáticas.[23][14] Sivaonyx beyi do Chade, especulado em pesar de 56,4 kg a 60,1 kg, tinha proporções de membros não especializadas que implicavam locomoção generalista-terrestre e adaptações aquáticas pobres.[25] Devido à hipótese de que S. beyi era um predador terrestre, especula-se que Enhydriodon dikikae era principalmente terrestre, com base em sua localização fóssil compartilhada com fauna aquática e terrestre em Dikika.[15] As paleobiologias e a partição de nicho de E. ekecaman e E. cf. dikikae em Kanapoi [en], Quênia, permanecem incertas, pois seus materiais fósseis, descobertos na década de 1960, não foram especificamente pronunciados além de "Kanapoi", o que pesquisas futuras teriam que cobrir.[26] Também é apontado que as espécies africanas de lontras bunodontes como Enhydriodon e Sivaonyx foram sempre encontradas em locais em associação com corpos de água permanentes, em oposição aos leitos Superiores de Laetoli em Laetoli, Tanzânia, que não possuíam tal característica, colocando em questão a extensão do estilo de vida possivelmente terrestre das espécies africanas de Enhydriodon e Sivaonyx.[27]
E. hendeyi foi analisado com base no índice de robustez femoral (FRI) e no índice epicondilar femoral (FEI), nos quais seu valor de FRI é comparável ao extinto S. beyi, Enhydritherium e Satherium [en](os dois últimos são análogos à grande lontra-marinha e à ariranha, respectivamente, e têm valores maiores nos índices femorais do que a maioria das outras lontras existentes), enquanto seu valor de FEI é análogo à lontra-africana-sem-garras e à lontra-anã-oriental (Aonyx cinereus) existentes, hipotetiza-se que Enhydriodon hendeyi e Sivaonyx beyi eram ambos locomotores semiaquáticos que tinham associações menores com a água do que os locomotores aquáticos Enhydritherium e Satherium, embora se diga que Sivaonyx beyi era mais terrestre que Enhydriodon hendeyi. Enquanto isso, os valores mais baixos correspondem a E. dikikae, que tem valores semelhantes aos musteloides terrestres semifossoriais (adaptados para cavar e viver um pouco no subsolo), como o texugo-americano e a doninha-fedorenta, reforçando assim a hipótese de que E. dikikae era um mustelídeo terrestre mais generalista, semelhante a Sivaonyx beyi.[13]
Com a falta geral de consenso sobre o estilo de vida das espécies africanas de Enhydriodon considerada, um estudo de 2022 sobre Enhydriodon omoensis mediu as razões de isótopos estáveis de carbono e oxigênio [en] de espécies de Enhydriodon em comparação com mamíferos terrestres existentes, como felídeos, hienídeos e bovídeos, juntamente com mamíferos semiaquáticos, como hipopotamídeos. Os autores explicaram que o uso de razões isotópicas de oxigênio, ou δ18O, pode ser usado para entender a dependência de um táxon da água, na qual os táxons aquáticos e semiaquáticos existentes, que incluem lontras de rio e marinhas, têm desvios isotópicos de oxigênio significativamente menores em comparação com os carnívoros terrestres. Os pesquisadores que estudaram Enhydriodon omoensis descobriram que os valores de δ18O do esmalte de seus dentes tinham um desvio padrão de 2,7%, caindo fora dos desvios padrão de δ18O da lontra-marinha e da lontra-norte-americana, que foram registrados como 0,6% e 0,3%-0,9%, respectivamente. O desvio padrão de Enhydriodon de Omo se alinha mais dentro da faixa de carnívoros terrestres existentes, como os hienídeos, sugerindo que E omoensis não era tão semiaquático quanto se pensava inicialmente. Os resultados do estudo contradizem a suposição de 2008 de que a espécie de Enhydriodon de Omo era aquática.[14]
Grohe et al. consideraram inicialmente que a dieta de Enhydriodon poderia ter sido a ostra Etheria elliptica [en], que estava presente no continente no mesmo intervalo de tempo. Com base em investigações usando isótopos estáveis de carbono, uma dieta de ostras puras resultaria em um valor de δ13C do esmalte de -11,3%. A dieta de E. omoensis, no entanto, não se baseava puramente em Etheria, pois seus valores mínimo-máximo de carbono (-9,7% a -4,7%) são ~2-7% mais positivos que o valor esperado para uma dieta de ostras puras. Seus valores de δ13C do esmalte caem na faixa de alimentadores mistos de C3-C4, caindo apenas parcialmente na faixa de dietas de alimentadores aquáticos de plantas C3, como peixes, tartarugas ou bivalves. O desvio padrão de δ13C de Enhydriodon de Omo, no entanto, cai fora da faixa de populações de lontras de água doce existentes estudadas. Em vez disso, considera-se que E. omoensis consumia presas terrestres com uma dieta C4 pelo menos semirregularmente, por meio de caça e/ou carniça. A grande dentição bunodonte da espécie sugere habilidades durofágicas [en] que lhe permitiam se alimentar de carniça, incluindo ossos, de maneira potencialmente semelhante a hienas ou mustelídeos esmagadores de ossos.[14]
Paleoecologia
Paquistão e Índia
Enhydriodon falconeri e E. sivalensis, embora ambas espécies de Enhydriodon que estiveram presentes em Sivalik, na Índia e no Paquistão, durante o período Neogeno, não coexistiram nas mesmas épocas, com base em suas aparições em depósitos de formação. Restos de E. falconeri estavam presentes na formação Nagri (Dhok Milan e Sethi Nagri, Paquistão) e na formação Dhok Pathan (Dhok Pathan e Hasnot, Paquistão), ambas as formações datando dos Sivaliks médios, representando o final do Mioceno. A espécie também estava presente na formação Tatrot (Tatrot, Índia) dos Sivaliks Superiores, do início ou meio do Plioceno. Nas formações Nagri e Dhok Pathan, foi demonstrado que E. falconeri existiu com várias famílias de mamíferos carnívoros arcaicos que se extinguiram antes do Plioceno, como os hienodontes [en] hienailurinos [en] e os anficíonídeos. As espécies de lontras primitivas também existiram com vários membros carnívoros extintos de famílias existentes durante o final do Mioceno, representando outros mustelídeos, ursídeos, felídeos (felinos e macairodontíneos), hienídeos (hienídeos percrocutíneos, ictiteres e hieninos), viverrídeos e herpestídeos. Sugere-se que a extinção dos anficíonídeos e percrocutídeos deixou nichos predatórios vazios que foram rapidamente preenchidos por outros gêneros de hienídeos, que se tornaram altamente diversificados e coexistiram com os felídeos no subcontinente.[28]
Outros membros extintos de famílias de mamíferos existentes e extintas foram encontrados na formação Nagri e, portanto, existiram com Enhydriodon falconeri, incluindo bovídeos, girafídeos, antracotérios, tragulídeos, suídeos, equídeos hiparioninos, rinocerotídeos, calicotérios, gonfotérios, hominídeos e espalacídeos.[29] Uma espécie de réptil extinta de gavial, Gavialis lewisi (?), é relatada da formação Dhok Pathan do Paquistão e é de idade Pliocena.[30] Gêneros de mamíferos encontrados na formação Dhok Pathan são geralmente consistentes com os gêneros de mamíferos encontrados na formação Nagri, mas também incluem outros bovídeos, girafídeos, cervídeos cervinos, antracotérios, suídeos, hiparioninos, rinocerotídeos, "gonfotérios tetralofodontes", cercopitecídeos e histricídeos.[31][32][33][34]

A transição do Mioceno médio para o Mioceno superior refletiu um período em que as florestas tropicais perenes a decíduas, que antes cobriam grande parte do subcontinente indiano, encolheram e foram substituídas por pastagens devido ao resfriamento global, condições mais secas e à intensificação das monções asiáticas.[35] Uma mudança das planícies de inundação de Nagri para as de Dhok Pathan sugere menos drenagem no sistema fluvial desta última em comparação com a primeira, com os rios menores de Dhok Pathan tendo um fluxo mais sazonal do que antes. Isso reflete a tendência geral do forçamento climático do final do Mioceno, resultando em mais sazonalidade, trazendo grandes mudanças faunísticas. Os climas mais secos e sazonais, juntamente com as mudanças fluviais, gradualmente deram origem a florestas abertas maiores, predominantemente compostas por plantas C4 perto dos rios do Mioceno de Potwar, enquanto as comunidades exclusivamente ou predominantemente compostas por plantas C3 diminuíram muito e eventualmente desapareceram por volta de 7,0 Ma, juntamente com os alimentadores de C3 que dependiam mais de vegetação fechada. Essas mudanças ocorreram logo após a chegada dos hiparioninos e marcaram diminuições em grupos de mamíferos no subcontinente indiano, como as extinções do hominídeo Sivapithecus e do dinotério Deinotherium, como resultado da fragmentação de habitats fechados em favor de habitats abertos que eliminariam o alimento para os navegadores e frugívoros de C3.[36][37][38]
.jpg)
Os registros fósseis de carnívoros da formação Tatrot, na Índia, são escassos, mas entre os membros extintos que existiram com Enhydriodon falconeri no Plioceno estavam outras lutrinas, macairodontíneos e hienídeos.[28] Os mamíferos herbívoros encontrados na formação Tatrot, no planalto de Potwar, contêm assembleias altamente diversas de bovídeos, mas também incluem cervídeos, suídeos, elefantídeos, estegodontídeos, hiparioninos, antracotérios, hipopotamídeos, girafídeos e tragulídeos.[39][40][41] Os crocodilianos Crocodylus e Rhamphosuchus, o pelicano Pelecanus, tartarugas (Batagur, Geoclemys [en], Hardella [en]e Pangshura [en]) e o caranguejo de água doce Acanthopotamon são relatados de pelo menos as formações Tatrot ou Pinjor da Índia também, indicando um habitat de água doce ativo no qual E. falconeri e, posteriormente, E. sivalensis estavam presentes.[30][42][43][44]
Entre os táxons de carnívoros, Enhydriodon é o gênero de caniforme de maior duração que já existiu nos Sivaliks do subcontinente indiano, identificado desde as formações Nagri-Pinjor. No entanto, a espécie identificada na formação Pinjor das épocas Plio-Pleistoceno é E. sivalensis, o que sugere que E. falconeri, após um longo tempo de sucesso relativo, pode ter passado por anagênese no Plioceno. Outros gêneros de carnívoros encontrados na formação Pinjor incluíam os recém-chegados canídeos, bem como mustelídeos, ursídeos, felídeos (macairodontíneos, panterinos e felinos), hienídeos e viverrídeos.[28] Outros gêneros de mamíferos encontrados na formação Pinjor incluem hominídeos, cercopitecídeos, roedores de várias famílias, proboscídeos, equinos da tribo Equini [en], rinocerotídeos, suídeos, cervídeos, girafídeos e bovídeos.[45]
Etiópia
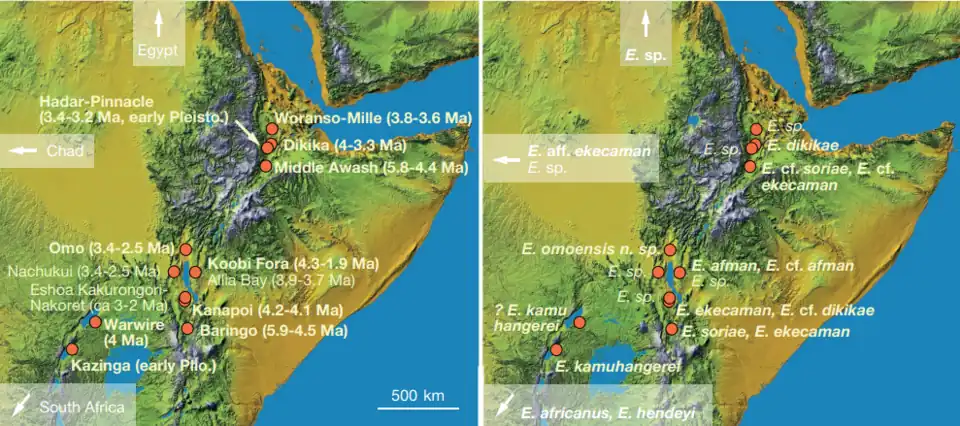
Enhydriodon dikikae e E. omoensis eram grandes espécies de lutrinas encontradas em diferentes locais na Etiópia moderna. Fósseis de E. dikikae foram encontrados nas duas sequências inferiores da formação Hadar, no vale do Baixo Awash, Etiópia, indicando que seus fósseis variam de 4 Ma a 3,2 Ma. Fósseis de E. omoensis foram localizados na formação Usno [en] e na formação Shungura [en], no vale do Baixo Omo, na Etiópia, com os fósseis variando de 3,44 Ma a 2,53 Ma. E. dikikae foi nomeado em homenagem ao Membro Basal de Dikikae da formação Hadar, enquanto Enhydriodon omoensis teve seu nome derivado do vale do Baixo Omo.[15][14]
Existem quatro membros [en] da sequência composta de Dikika como parte da formação Hadar do Plioceno, da base ao topo: os membros Basal, Sidi Hakoma, Denen Dora e Kada Hadar. Todos juntos, são datados de ca. 3,5-2,9 Ma e são mais conhecidos pelos numerosos restos de Australopithecus afarensis.[46] Fósseis de Enhydriodon dikikae são conhecidos dos membros Basal e Sigi Hakoma da formação e são desconhecidos nos outros dois membros superiores.[15]
Com base em métodos para determinar paleoambientes, como análise ecomorfológica, microdesgaste dentário de bovídeos e isótopos de carbono e oxigênio do esmalte, o Membro Basal (BM) tem a maior abundância de bovídeos e suídeos na formação Hadar, sugerindo que os ambientes em que estavam presentes eram possivelmente pastagens arborizadas, bem como florestas ribeirinhas. Aepycerotini eram comuns dentro do membro, o que se encaixa na preferência da tribo por habitats ecotonais entre pastagens e matas.[47]
O Submembro 1 de Sidi Hakoma (SH-1), que varia de ~3,45 a 3,35 Ma, tinha fauna e, portanto, habitats semelhantes a outros membros da formação Hadar, mas também provavelmente incluía zonas húmidas em certas regiões. Táxons como uma espécie da tribo Cephalophini, que habita florestas, e cinco espécies de primatas foram recuperados do membro, indicando ainda uma grande floresta ribeirinha com, predominantemente, matas na área circundante. Aepyceros era o bovídeo mais abundante, e SH-1 tinha a menor proporção de bovídeos pastadores de qualquer submembro da formação Hadar. A vegetação de SH-1 pode ter se assemelhado muito à das savanas da Guiné ou Sudanesas que se interdigitam com a floresta tropical da África Central, o que cria mosaicos de habitats de pastagens, matas e algumas faixas de floresta. A assembleia de ostracodes dos membros Basal e Sidi Hakoma indica fontes de entrada de água doce, cujas conchas também indicam apenas uma estação seca de três meses, característica das savanas da África Central. A única estação seca, indicando uma estação chuvosa de nove meses, é um fator distintivo do membro Sidi Hakoma em relação ao clima moderno na África Oriental, que tem um formato de estação seca bimodal (duas estações secas) em vez de uma única. O Submembro 2 de Sidi Hakoma (SH-2) é semelhante a SH-1 e acredita-se que tenha sido associado a matas com algumas planícies gramadas, das quais Aepycerotini eram os mais comuns.[48]
O Submembro 3 de Sidi Hakoma (SH-3) indica a presença de matas e pastagens com mais zonas húmidas à beira do lago em comparação com os submembros anteriores, com presenças aumentadas de bovídeos Reduncinae e a maior abundância de bovídeos Tragelaphini, que indicam habitats mais fechados ou zonas húmidas. Ele também contém as maiores assembleias de micromamíferos de gêneros de murídeos existentes, como os existentes Acomys, Golunda e Oenomys e o extinto Saidomys [en], dos quais Golunda está agora extinto na África. O Submembro 4 de Sidi Hakoma indica habitats de zonas húmidas que cercam lagos dentro de ambientes mais secos. Um aumento adicional de bovídeos Reduncinae e uma diminuição de bovídeos Alcelaphinae indicam os referidos ambientes de margem de lago e zonas húmidas circundantes. Os dados de abundância de bovídeos sugerem quantidades semelhantes de cobertura arbórea para SH-3 e SH-4, com a diferença sendo que este último era ligeiramente mais seco que o primeiro.[48]
_National_Museum_of_Ethiopia.jpg)
A formação Hadar representa muitos fósseis de Australopithecus afarensis, mais notavelmente o esqueleto parcial conhecido como "Lucy". O intervalo de tempo agregado da espécie é de pelo menos 0,7 milhões de anos, de 3,7 Ma a 3,0 Ma.[47] A formação Hadar também é conhecida por sua representação de uma grande diversidade de espécies de bovídeos que representavam a maioria das principais tribos da África. As tribos de bovídeos encontradas na formação incluíam Aepycerotini, Alcelaphini, Antilopini, Bovini, Caprini, Cephalophini, Hippotragini, Neotragini, Reduncini e Tragelaphini. Outros grupos, representados por espécies extintas de gêneros existentes ou extintos, incluem girafídeos, hipopótamos, suídeos, canídeos, felídeos (macairodontíneos eram os mais comuns), hienídeos, outros mustelídeos, viverrídeos, rinocerontes, equídeos, catarrinos, dinotérios e elefantídeos, todos também encontrados na localidade. Grupos de pequenos mamíferos incluem morcegos, leporídeos, porcos-espinhos do velho mundo, roedores murídeos, espalacídeos, esquilos e porcos-formigueiros.[46][48]
Outras formações da idade Pliocena na Etiópia mostram tendências semelhantes de grande diversidade na família Bovidae de suas múltiplas tribos, juntamente com suídeos, hipopotamídeos, cercopitecídeos, hominídeos e equídeos, geralmente dos mesmos gêneros da formação Hadar. A maioria dos herbívoros presentes na formação Shungura mostra dietas consistentes de C4 ou geralmente mudou de dietas mistas C3-C4 para dietas geralmente de C4, como indicado por mudanças na dentição por membro da formação. Essas tendências sugerem que os herbívoros africanos no Plioceno estavam mudando cada vez mais para a herbivoria C4, em oposição à navegação e à alimentação mista, como resultado da crescente dominância de pastagens C4 na África. Houve algumas exceções, no entanto, já que Giraffidae e Deinotheriidae eram ambos navegadores consistentes de C3 dentro da formação, enquanto as tribos de bovídeos Aepycerotini e Tragelaphini eram predominantemente alimentadores mistos com pouca mudança na dieta.[49][50][51] Restos de peixes fósseis também são conhecidos da formação Shungura, nomeadamente os gêneros Polypterus [en], Sindacharax, Synodontis, Auchenoglanis [en] e Lates [en].[52]
Notas
Referências
- ↑ a b c Hugh Falconer (1868). Palæontological Memoirs and Notes of the Late Hugh Falconer: Fauna Antiqua Sivalensis. Reino Unido: R. Hardwicke. ISBN 112-0-015839
- ↑ a b c Verma, B.C.; Gupta, S.S. (1992). «Enhydriodon sivalensis, a giant fossil otter from the Saketi Formation (Upper Pliocene), Siwalik group, Sirmur district, Himachal Pradesh». Journal of the Palaeontological Society of India. 37 (1): 31–36. Bibcode:1992JPalS..37...31V. doi:10.1177/0971102319920102
- ↑ Cautley, Proby Thomas; Falconer, Hugh (1835). «Synopsis of Fossil Genera and Species from the upper deposits of the tertiary strata of the Sivalik Hills, in the collection of the authors.». Journal of the Asiatic Society of Bengal. 4 (12)
- ↑ a b Guy E. Pilgrim (1932). The Fossil Carnivora of India. Índia: Palaeontologia Indica. pp. 80–88
- ↑ a b c Willemsen, Gerard F. (1992). «A revision of the Pliocene and Quaternary Lutrinae from Europe». Scripta Geologica. 101: 89–94
- ↑ Repenning, Charles A. (1976). «Enhydra and Enhydriodon From The Pacific Coast of North America». Journal of Research of the U.S. Geological Survey. 4 (3): 305–315. Bibcode:1976JRUGS...4..305R
- ↑ Hürzeler, Johannes; Engesser, Burkart (1976). «Les faunes de mammifères néogènes du Bassin de Baccinello (Grosseto, Italie)». Comptes Rendus de l'Académie des Sciences de Paris (em francês). 283: 333–336
- ↑ a b c d e f Morales, Jorge; Pickford, Martin (2005). «Giant bunodont Lutrinae from the Mio-Pliocene of Kenya and Uganda». Estudios Geológicos (em espanhol). 61 (3): 233–245. doi:10.3989/egeol.05613-666

- ↑ Pickford, Martin; Soria, Dolore; Morales, Jorge (2005). «Carnivores from the Late Miocene and Basal Pliocene of the Tugen Hills, Kenya». Revista de la Sociedad Geológica de España. 18 (1–2): 39–61
- ↑ a b c d e f g h Pickford, Martin (2007). «Revision of the Mio-Pliocene bunodont otter-like mammals of the Indian Subcontinent». Estudios Geológicos (em espanhol). 63 (2): 108–124. doi:10.3989/egeol.07631192
- ↑ a b c d e f Wang, Xiaoming; Grohé, Camille; Su, Denise F.; White, Stuart C.; Ji, Xueping; Kelley, Jay; Jablonski, Nina G.; Deng, Tao; You, Youshan; Yang, Xin (2017). «A new otter of giant size, Siamogale melilutra sp. nov. (Lutrinae: Mustelidae: Carnivora), from the latest Miocene Shuitangba site in north-eastern Yunnan, south-western China, and a total-evidence phylogeny of lutrines». Journal of Systematic Palaeontology. 16 (1): 1–24. Bibcode:2018JSPal..16...39W. doi:10.1080/14772019.2016.1267666
- ↑ Werdelin, Lars (2003). «Carnivores from the Kanapoi Hominid site, Turkana Basin, northern Kenya». Contributions in Science. 498: 115–132. doi:10.5962/p.214388
- ↑ a b c d e Valenciano, Alberto; Govender, Romala (2020). «New insights into the giant mustelids (Mammalia, Carnivora, Mustelidae) from Langebaanweg fossil site (West Coast Fossil Park, South Africa, early Pliocene)». PeerJ. 8. PMC 7271888. PMID 32547866. doi:10.7717/peerj.9221
- ↑ a b c d e f g h i j k Grohé, Camille; Uno, Kevin; Boisserie, Jean-Renaud (2022). «Lutrinae Bonaparte, 1838 (Carnivora, Mustelidae) from the Plio-Pleistocene of the Lower Omo Valley, southwestern Ethiopia: systematics and new insights into the paleoecology and paleobiogeography of the Turkana otters». Comptes Rendus Palevol (em francês). 30 (30): 684–693. doi:10.5852/cr-palevol2022v21a30
- ↑ a b c d e f g h i j k Geraads, Denis; Alemseged, Zeresenay; Bobe, René; Reed, Denné (2011). «Enhydriodon dikikae, sp. nov. (Carnivora: Mammalia), a gigantic otter from the Pliocene of Dikika, Lower Awash, Ethiopia». Journal of Vertebrate Paleontology. 31 (2): 447–453. Bibcode:2011JVPal..31..447G. doi:10.1080/02724634.2011.550356
- ↑ Valenciano, Alberto; Morales, Jorge; Pickford, Martin (2022). "Lutra" libyca from the Mio/Pliocene of Wadi Natrun (Egypt): a new systematics proposal. The 2nd International Symposium on Vertebrate Paleontology 20th-22nd September 2022. pp. 31–32
- ↑ a b Kargopoulos, Nikolaos; Valenciano, Alberto; Kampouridis, Panagiotis; Lechner, Thomas; Böhme, Madelaine (2021). «New early late Miocene species of Vishnuonyx (Carnivora, Lutrinae) from the hominid locality of Hammerschmiede, Bavaria, Germany». Journal of Vertebrate Paleontology. 41 (3). Bibcode:2021JVPal..41E8858K. doi:10.1080/02724634.2021.1948858
- ↑ «Enhydritherium terraenovae» (em inglês). Florida Museum of Natural History. 30 de março de 2017. Consultado em 10 de outubro de 2022
- ↑ Constantino, Paul J.; Lee, James J.-W.; Morris, Dylan J.; Lucas, Peter W.; Hartstone-Rose, Adam; Lee, Wah-Keat; Dominy, Nathaniel J.; Cunningham, Andrew; Wagner, Mark; Lawn, Brian R. (2011). «Adaptation to hard-object feeding in sea otters and hominins». Journal of Human Evolution. 61 (1): 89–96. Bibcode:2011JHumE..61...89C. PMID 21474163. doi:10.1016/j.jhevol.2011.02.009
- ↑ Raghavan, P.; Pickford, M.; Patnaik, R.; Gayathri, P. (2007). «First fossil small-clawed otter, Amblonyx, with a note on some specimens of Lutra, from the Upper Siwaliks, India». Estudios Geológicos (em espanhol). 63 (2): 144–145. doi:10.3989/egeol.07632200
- ↑ Koufos, George D. (2011). «The Miocene carnivore assemblage of Greece». Estudios Geológicos (em espanhol). 67 (2): 296–297. doi:10.3989/egeol.40560.190
- ↑ Alberto Valenciano Vaquero; Romala Govender (21 de junho de 2020). «Texugos gigantes, lontras do tamanho de lobos: fósseis oferecem novas perspectivas sobre o passado». The Conversation. Consultado em 27 de novembro de 2022
- ↑ a b Lewis, Margaret E. (2008). «The femur of extinct bunodont otters in Africa (Carnivora, Mustelidae, Lutrinae)». Comptes Rendus Palevol (em francês). 7 (8): 607–627. Bibcode:2008CRPal...7..607L. doi:10.1016/j.crpv.2008.09.010
- ↑ Stewart, Kathlyn M (2001). «The freshwater fish of Neogene Africa (Miocene–Pleistocene): systematics and biogeography». Fish and Fisheries. 2 (3): 177–230. Bibcode:2001AqFF....2..177S. doi:10.1046/j.1467-2960.2001.00052.x
- ↑ Peigné, Stéphane; De Bonis, Louis; Likius, Andossa; Mackaye, Hassane Taïsso; Vignaud, Patrick; Brunet, Michel (2008). «Late Miocene Carnivora from Chad: Lutrinae (Mustelidae)». Zoological Journal of the Linnean Society. 152 (4): 793–846. doi:10.1111/j.1096-3642.2008.00377.x
- ↑ Werdelin, Lars; Manthi, Fredrick Kyalo (2012). «Carnivora from the Kanapoi hominin site, northern Kenya». Journal of African Earth Sciences. 64: 1–8. Bibcode:2012JAfES..64....1W. doi:10.1016/j.jafrearsci.2011.11.003
- ↑ Werdelin, Lars; Lewis, Margaret E. (2020). «A contextual review of the Carnivora of Kanapoi». Journal of Human Evolution. 140. Bibcode:2020JHumE.14002334W. PMID 28625408. doi:10.1016/j.jhevol.2017.05.001
- ↑ a b c Jasinski, Steven E.; Abbas, Sayyed Ghyour; Mahmood, Khalid; Babar, Muhammad Adeeb; Khan, Muhammad Akbar (2022). «New Carnivoran(Mammalia: Carnivora) specimens from the Siwaliks of Pakistan and India and their faunal and evolutionary implications». Historical Biology: An International Journal of Paleobiology. 35 (11): 1–36. doi:10.1080/08912963.2022.2138376
- ↑ Khan, Muhammad Akbar; Akhtar, Muhammad; Ikram, Tasneem. «True Ungulates From The Nagri Type Locality (Late Miocene), Northern Pakistan». Journal of Animal and Plant Sciences. 22: 1–59
- ↑ a b Martin, Jérémy (2018). «The taxonomic content of the genus Gavialis from the Siwalik Hills of India and Pakistan» (PDF). Papers in Palaeontology. 5 (3): 483–497. doi:10.1002/spp2.1247
- ↑ Khan, Muhammad Akbar; Akhtar, Muhammad; Iliopoulos, George (2012). «Tragulids (Artiodactyla, Ruminantia, Tragulidae) from the middle Siwaliks of Hasnot (Late Miocene), Pakistan». Rivista Italiana di Paleontologia e Stratigrafia. 118 (2): 325–341. doi:10.13130/2039-4942/6009
- ↑ Ghaffar, Abdul; Akhtar, Muhammad; Babar, Muhammad Adeeb; Roohi, Ghazala (2018). «Cervidae from Dhok Pathan and Hasnot areas (Dhok Pathan Formation, Northern Pakistan) with comments on the age of these sediments». Revista Brasileira de Paleontologia. 21 (2): 132–140. Bibcode:2018RvBrP..21..132G. doi:10.4072/rbp.2018.2.04
- ↑ Abbas, Ghyour; Babar, Muhammad Adeeb; Khan, Muhammad Akbar; Un Nisa, Badar (2021). «New proboscidean material from the Siwalik Group of Pakistan with remarks on some species». PaleoBios. 38 (1): 1–12. doi:10.5070/P9381054113
- ↑ Babar, Muhammad Adeeb; Aftab, Kiran; Khan, Muhammad Akbar; Abbas, Ghyour (2021). «Tetraconodon and Sivachoerus (Suidae) from Dhok Pathan and Hasnot area of Potwar Plateau, northern Pakistan». Arabian Journal of Geosciences. 14 (2473): 1–6. Bibcode:2021ArJG...14.2473B. doi:10.1007/s12517-021-08684-x
- ↑ Wang, Xiaoming; Flynn, Lawrence; Fortelius, Mikael (2013). «Chapter 17: Indian Neogene Siwalik Mammalian Biostratigraphy: An Overview». In: Wang, Xiaoming; Flynn, Lawrence J.; Fortelius, Mikael. Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology (em inglês). [S.l.]: Columbia University Press. pp. 423–444. ISBN 9780231520829. doi:10.7312/wang15012-017
- ↑ Barry, John C.; Morgan, Michèle E.; Flynn, Lawrence J.; Pilbeam, David; Behrensmeyer, Anna K.; Raza, S. Mahmood; A. Khan, Imran; Badgley, Catherine; Hicks, Jason; Kelley, Jay (2002). «Faunal and Environmental Change in the Late Miocene Siwaliks of Northern Pakistan». Paleobiology. 28 (2): 1–71. doi:10.1666/0094-8373(2002)28[1:FAECIT]2.0.CO;2
- ↑ Nelson, Sherry V. (2007). «Isotopic reconstructions of habitat change surrounding the extinction of Sivapithecus, a Miocene hominoid, in the Siwalik Group of Pakistan» (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 243 (1–2): 204–222. Bibcode:2007PPP...243..204N. doi:10.1016/j.palaeo.2006.07.017
- ↑ Patnaik, Rajeev; Singh, Ningthoujam Premjit; Paul, Debajyoti; Sukumar, Raman (2019). «Dietary and habitat shifts in relation to climate of Neogene-Quaternary proboscideans and associated mammals of the Indian subcontinent». Quaternary Science Reviews. 224. Bibcode:2019QSRv..22405968P. doi:10.1016/j.quascirev.2019.105968
- ↑ Jukar, Advait Muhesh; Sun, Boyang; Nanda, Avinash C.; Bernor, Raymond L. (2019). «The first occurrence of Eurygnathohippus Van Hoepen, 1930 (Mammalia, Perissodactyla, Equidae) outside Africa and its biogeographic significance». Bollettino della Società Paleontologica Italiana. 58 (2): 171–179. doi:10.4435/BSPI.2019.13 (inativo 12 de Julho de 2025)
- ↑ Khan, Muhammad Akbar; Jamil, Madeeha; Mahmood, Khalid; Babar, Muhammad Adeeb (2014). «Some New Remains of Cervids (Cervidae, Ruminantia) from Tatrot Formation of Northern Pakistan». Biologia. 60 (2): 225–230
- ↑ Kumar, Suresh (2022). «New Fossils of Sivatherium Giganteum (Giraffidae, Mammalia) from the Upper Siwaliks of the Indian Subcontinent». Acta Palaeontologica Romaniae. 18 (2): 85–92. doi:10.35463/j.apr.2022.02.04
- ↑ Thomas A., Stidham; Krishan, Kewal; Singh, Bahadur; Ghosh, Abhik; Patnaik, Rajeev (2014). «A Pelican Tarsometatarsus (Aves: Pelecanidae) from the Latest Pliocene Siwaliks of India». PLOS ONE. 9 (11): 1–8. Bibcode:2014PLoSO...9k1210S. PMC 4218845. PMID 25365300. doi:10.1371/journal.pone.0111210
- ↑ Garbin, Rafaella C.; Bandyopadhyay, Saswati; Joyce, Walter G. (2020). «A taxonomic revision of geoemydid turtles from Siwalik-age of India and Pakistan». European Journal of Taxonomy (652): 1–67. doi:10.5852/ejt.2020.724.1165
- ↑ Klaus, Sebastian; Singh, Bahadur; Hartmann, Lukas; Krishan, Kewal; Ghosh, Abhik; Patnaik, Rajeev (2017). «A fossil freshwater crab from the Pliocene Tatrot Formation (Siwalik Group) in Northern India (Crustacea, Brachyura, Potamidae)». Palaeoworld. 26 (3): 566–571. doi:10.1016/j.palwor.2016.08.003
- ↑ Nanda, Avinash C.; Sehgal, Ramesh K.; Chauhan, Parth R. (2018). «Siwalik-age faunas from the Himalayan Foreland Basin of South Asia». Journal of Asian Earth Sciences. 162: 54–68. Bibcode:2018JAESc.162...54N. doi:10.1016/j.jseaes.2017.10.035
- ↑ a b Geraads, Denis; Alemseged, Zeresenay; Bobe, René; Reed, Denné (2015). «Pliocene Carnivora (Mammalia) from the Hadar Formation at Dikika, Lower Awash Valley, Ethiopia». Journal of African Earth Sciences. 107: 28–35. Bibcode:2015JAfES.107...28G. doi:10.1016/j.jafrearsci.2015.03.020
- ↑ a b Alemseged, Zeresenay; Wynn, Jonathan G.; Kimbel, William H.; Reed, Denné; Geraads, Denis; Bobe, René (2005). «A new hominin from the Basal Member of the Hadar Formation, Dikika, Ethiopia, and its geological context». Journal of Human Evolution. 49 (4): 499–514. Bibcode:2005JHumE..49..499A. PMID 16026815. doi:10.1016/j.jhevol.2005.06.001
- ↑ a b c Campisano, Christopher J.; Rowan, John; Reed, Kaye E. (2022). «Chapter 18: The Hadar Formation, Afar Regional State, Ethiopia: Geology, Fauna, and Paleoenvironmental Reconstructions». In: Reynolds, Sally C.; Bobe, René. African Paleoecology and Human Evolution (em inglês). [S.l.]: Cambridge University Press. pp. 214–228. doi:10.1017/9781139696470.018
- ↑ Bobe, René (2011). «Fossil Mammals and Paleoenvironments in the Omo-Turkana Basin». Evolutionary Anthropology. 20 (6): 254–263. PMID 22170694. doi:10.1002/evan.20330
- ↑ Negash, Enquye W.; Alemseged, Zeresenay; Bobe, René; Grine, Frederick; Sponheimer, Matt; G. Wynn, Jonathan (2020). «Dietary trends in herbivores from the Shungura Formation, southwestern Ethiopia». Proceedings of the National Academy of Sciences. 117 (36): 21921–21927. Bibcode:2020PNAS..11721921N. PMC 7486712. PMID 32839326. doi:10.1073/pnas.2006982117
- ↑ Bibi, Faysal; Souron, Antoine; Bocherens, Hervé; Uno, Kevin; Boisserie, ean-Renaud (2013). «Ecological change in the lower Omo Valley around 2.8 Ma». Biology Letters. 9 (1): 1–4. PMC 3565503. PMID 23234862. doi:10.1098/rsbl.2012.0890
- ↑ Stewart, Kathlyn M.; Murray, Alison (2008). «Fish remains from the Plio-Pleistocene Shungura Formation, Omo River basin, Ethiopia». Geobios. 41 (2): 283–295. Bibcode:2008Geobi..41..283S. doi:10.1016/j.geobios.2007.06.004