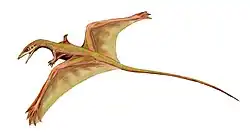
Dinocephalosaurus
Dinocephalosaurus
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Anisiano, 245–244 Ma | |||||||||||||||
.png) Esqueleto completo e articulado (IVPP V20295). | |||||||||||||||
| Classificação científica | |||||||||||||||
| |||||||||||||||
Dinocephalosaurus (que significa "réptil de cabeça terrível") é um gênero de protorossauro [en] aquático de pescoço longo que habitava os mares do Triássico na China. O gênero contém a espécie-tipo e única espécie conhecida, D. orientalis, nomeada por Chun Li em 2003. Diferentemente de outros protorossauros de pescoço longo (que formam um grupo conhecido como tanistrofeídeos), o Dinocephalosaurus evoluiu convergentemente um pescoço longo não por meio do alongamento de vértebras cervicais individuais, mas pela adição de vértebras cervicais de comprimento moderado.
Como indicado por análises filogenéticas, ele pertencia a uma linhagem distinta que também incluía pelo menos seu parente mais próximo, Pectodens, que foi nomeado Dinocephalosauridae em 2021. Como os tanistrofeídeos, no entanto, o Dinocephalosaurus provavelmente usava seu pescoço longo para caçar, utilizando os dentes semelhantes a presas de suas mandíbulas para capturar presas; propostas de que ele empregava alimentação por sucção não foram universalmente aceitas. Provavelmente era um animal marinho por necessidade, como sugerido pelos membros pouco ossificados e semelhantes a nadadeiras, que o teriam impedido de ir para a terra.
Espécimes pertencentes ao gênero foram descobertos pela primeira vez em uma localidade perto de Xinmin, em Guizhou, China, em 2002. Na mesma localidade, datada de 244 milhões de anos atrás, outros répteis marinhos como Mixosaurus [en], Keichousaurus e Wumengosaurus também foram encontrados. Embora o espécime-tipo consistisse apenas de um crânio e a parte frontal do pescoço, espécimes adicionais logo revelaram a forma completa do corpo. Outras descobertas de espécimes de Dinocephalosaurus foram feitas em Luoping, Yunnan, China, a partir de 2008. Nesta localidade, o Dinocephalosaurus teria vivido ao lado de Mixosaurus, Dianopachysaurus e Sinosaurosphargis [en]. Um espécime descoberto na localidade de Luoping preserva um embrião em seu abdômen, indicando que o Dinocephalosaurus dava à luz filhotes vivos, como muitos outros répteis marinhos. O Dinocephalosaurus é o único membro conhecido do grupo Archosauromorpha a dar à luz filhotes vivos, com a possível exceção dos metriorrinquídeos, um grupo de crocodilomorfos marinhos.
Descoberta e nomeação
Panxian
.png)
O espécime-tipo de Dinocephalosaurus foi descoberto pela primeira vez em 2002, durante trabalhos de campo conduzidos na Vila de Yangjuan, Distrito de Xinmin, Condado de Panxian, Guizhou, China. Ele consiste em um crânio quase completo, faltando o lado esquerdo da mandíbula, além de várias vértebras cervicais associadas. Foi posteriormente armazenado no Instituto de Paleontologia de Vertebrados e Paleoantropologia (IVPP) em Pequim, China, sob o número de coleção IVPP V13767, e um artigo de pesquisa descrevendo o espécime foi publicado por Chun Li na "Acta Geologica Sinica" em dezembro de 2003.[1]
Um segundo espécime descoberto na mesma localidade representa um esqueleto parcialmente articulado, faltando apenas a cauda. Também armazenado no IVPP, o espécime possui o número de coleção IVPP V13898. O espécime foi descrito em uma breve correspondência de autoria de Li, Rieppel e LaBarbera, publicada pela Science em setembro de 2004;[2] uma descrição mais detalhada foi posteriormente publicada por Rieppel, Li e Fraser em um artigo de 2008 no Journal of Vertebrate Paleontology.[3]
A localidade de Panxian, de onde esses espécimes se originaram, faz parte do Membro II da formação Guanling anisiana (Triássico Médio),[4] que inicialmente se pensava ter 230 milhões de anos,[1][5] mas foi mais recentemente datada em 244 ± 1,3 milhões de anos com base em datação por urânio-chumbo [en].[6] Os depósitos predominantes nesta localidade são compostos por marga cinza a cinza escura e calcário com cherte, contendo camadas de dolomita e bentonita. O Dinocephalosaurus foi encontrado especificamente na camada 90 dos depósitos de Panxian, uma fina camada de calcário que é tradicionalmente atribuída ao "horizonte superior de répteis" (camadas 87–90). Mais abaixo estão os horizontes "médio de répteis" (camadas 81–85) e "inferior de répteis" (77–79).[4]
Luoping
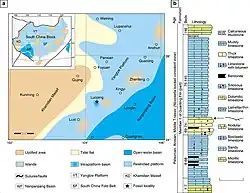
Posteriormente, espécimes adicionais de Dinocephalosaurus foram descobertos na localidade de Luoping, ligeiramente mais antiga, datada de 245–244 milhões de anos com base em bioestratigrafia de conodontes[7] e datação radiométrica preliminar.[8] Os espécimes provêm da camada 74 da localidade de Luoping, em depósitos localizados perto da vila de Dawazi, Condado de Luoping, Yunnan, China. A camada 74 faz parte de uma seção composta por finas camadas de calcário micrítico cinza escuro misturadas com camadas mais espessas de calcário silicioso e calcário siltoso, que se estende para baixo até a camada 67.[4][8]
Um espécime em particular é notável por conter um embrião em sua região abdominal, no qual estão preservadas vértebras cervicais, membros anteriores e vários outros elementos. Ele foi coletado em 2008; no momento de sua coleta, o intemperismo já havia dividido o espécime em três blocos, com os espaços preenchidos por solo moderno. O espécime foi então transferido para o Serviço Geológico da China de Chengdu onde foi preparado e armazenado sob o número de coleção LPV 30280. Posteriormente, uma descrição do espécime, de autoria de Liu e colaboradores, foi publicada em fevereiro de 2017 pela Nature Communications.[8]
Pelo menos um outro espécime não descrito de Dinocephalosaurus é conhecido de uma localidade não especificada, tendo sido catalogado como ZMNH M8752 no Museu de História Natural de Zhejiang. Ele foi mencionado brevemente em comparação com Fuyuansaurus por Fraser, Rieppel e Li em 2013.[9] Além disso, outro esqueleto embrionário catalogado como IVPP V22788 foi encontrado em Luoping e foi considerado por eles como intimamente relacionado com o Dinocephalosaurus devido ao grande número de vértebras cervicais curtas e seus membros em forma de pilares. No entanto, eles também notaram algumas diferenças provavelmente não relacionadas ao crescimento, como a presença de menos vértebras cervicais (24 em vez de 33) e a presença de um anel esclerótico, o que os levou a atribuí-lo não ao Dinocephalosaurus, mas a um animal intimamente relacionado.[10]
Nomeação
Na descrição de 2003, Li combinou as raízes din- ("terrível"), cephalo- ("cabeça") e saur ("lagarto") para criar o nome do gênero Dinocephalosaurus. Segundo Li, esse nome refere-se ao "crânio assustador" do holótipo. Enquanto isso, o nome da espécie deriva do latim orientalis ("oriental"), em referência ao D. orientalis representar o único registro conhecido na época dos Tanystropheidae na porção oriental do oceano de Tétis.[1]
Descrição

O Dinocephalosaurus era um arcosauromorfo não crocopodano muito grande, atingindo um comprimento corporal máximo de até 6 m,[11] em comparação com um máximo de 5 a 6 m para Tanystropheus.[12] Os espécimes conhecidos provavelmente eram maduros, dado que possuem ossos cranianos fundidos e não apresentam os dentes multicúspides vistos em juvenis de Tanystropheus.[3]
Crânio

O crânio do Dinocephalosaurus era baixo e estreito, com uma pré-maxila e maxila longas em comparação com as de Tanystropheus. Tanto a pré-maxila quanto a maxila se encontravam na esquina inferior frontal e contribuíam para a borda da narina, que estava localizada na extremidade frontal de um longo recesso que se estendia ao longo do focinho à frente da órbita ocular (a depressão antorbital). Esse recesso também estava presente em Macrocnemus [en] e Fuyuansaurus.[9] Como em Pectodens [en], a espessura da pré-maxila fazia com que as narinas fossem retraídas da ponta do focinho.[13] As margens inferiores dos dois ossos do focinho eram respectivamente alinhadas com cinco e doze dentes longos e semelhantes a pregos; o terceiro dente pré-maxilar e o quarto e quinto dentes maxilares eram distintamente semelhantes a presas. A mandíbula inferior preserva quinze dentes, com três deles sendo semelhantes a presas e formando uma "armadilha para peixes".[3][13] Em comparação, os dentes em adultos de Tanystropheus eram pinos afiados, enquanto eram tricúspides (com três cúspides) em juvenis.[1]
Diferentemente da órbita oval de Tanystropheus, a órbita do Dinocephalosaurus parece ter sido em forma de pêssego, com uma extremidade frontal estreita. No topo do crânio, o osso parietal era largo e achatado, sem vestígios da crista mediana encontrada em Tanystropheus.[1] O jugal possuía apenas dois processos, faltando o terceiro processo projetado para trás presente na maioria dos outros arcosauromorfos,[3] mas compartilhado com Pectodens, Claudiosaurus e Trilophosaurus [en].[13] Também estavam ausentes o processo retroarticular da mandíbula inferior traseira (outro ponto de distinção de Tanystropheus),[1] bem como dentes adicionais e uma cavidade entre os pterigoides no palato.[3]
Pescoço e tronco
.png)
Como o Tanystropheus, o Dinocephalosaurus tinha um pescoço excepcionalmente longo (1.7 m de comprimento) em relação ao seu tronco (1 m excluindo a cauda).[2] Tanystropheus e Dinocephalosaurus alcançaram seus pescoços extremamente alongados de maneiras diferentes. O pescoço do Tanystropheus era composto por 13 vértebras cervicais alongadas,[14] enquanto o pescoço do Dinocephalosaurus era composto por pelo menos 27 vértebras cervicais que não eram tão alongadas. Entre as 27 vértebras do Dinocephalosaurus, a mais longa era a décima nona, que media aproximadamente 91 mm de comprimento. Em comparação, as vértebras mais longas em um espécime chinês de Tanystropheus eram a nona e a décima, que mediam 248 mm de comprimento. Além disso, as vértebras cervicais do Dinocephalosaurus não eram ocas, ao contrário das de Tanystropheus.[15]
Características adicionais das vértebras cervicais que distinguiam o Dinocephalosaurus de outros protorossauros incluíam as espinhas neurais baixas e em forma de quilha com bordas superiores côncavas, e as superfícies articulares frontal e traseira das vértebras sendo ambas côncavas (anficélicas). Nas primeiras dez vértebras cervicais, a margem inferior também era côncava. As costelas cervicais longas e esguias apresentavam projeções frontais livres dos corpos vertebrais, uma característica rara vista apenas em Pectodens, Czatkowiella, Sclerostropheus e Tanytrachelos.[13] Essas costelas estavam alinhadas ao longo do pescoço e conectavam várias articulações vertebrais consecutivas, de duas ou três articulações consecutivas na frente do pescoço a cinco ou seis na parte traseira do pescoço. Não parece haver processos distintos nas vértebras para articulação com as costelas.[3]
O tronco do Dinocephalosaurus tinha uma contagem muito alta de pelo menos 26 vértebras.[13] No Dinocephalosaurus, não parece haver vértebras lombares, ou vértebras do tronco sem costelas. As costelas do sacro e da cauda também não parecem estar fundidas às suas vértebras correspondentes. Cada um dos gastralium no Dinocephalosaurus era composto por três elementos em vez de quatro como no Tanystropheus; eles diferiam no fato de que o Dinocephalosaurus tinha apenas um elemento na linha média, enquanto o Tanystropheus tinha dois elementos que se combinavam para formar uma barra na linha média.[3]
Membros
.png)
O Dinocephalosaurus tinha pernas relativamente grandes terminando em pés semelhantes a nadadeiras. Os membros anteriores e posteriores têm aproximadamente o mesmo comprimento, ao contrário do Tanystropheus, onde os membros anteriores eram muito menores.[16] Enquanto a maioria dos protorossauros, como Tanystropheus, Macrocnemus e Langobardisaurus [en], tinham membros relativamente ossificados adaptados para a vida terrestre, os membros robustos do Dinocephalosaurus eram pouco ossificados e se assemelhavam aos dos notossauros. Dos ossos do carpo, apenas seis eram ossificados; da mesma forma, apenas três dos ossos do tarso eram ossificados. Além disso, o astrágalo e o calcâneo também não se articulavam entre si no tornozelo, formando ossificações simples e arredondadas.[2][3] Outra adaptação à vida aquática foi a perda de uma abertura conhecida como fenestra tiroide na pelve devido aos ossos pélvicos arredondados, que também estava presente em Fuyuansaurus.[9]
Essas características eram provavelmente neotênicas, o que também caracteriza traços encontrados em muitos outros tetrapodes aquáticos. Embora o Tanystropheus também fosse provavelmente neotênico, ele não se aproximava do Dinocephalosaurus na extremidade dessa condição.[3] Várias outras características peculiares estavam presentes nos pés do Dinocephalosaurus. Diferentemente do Tanystropheus e da maioria dos outros protorossauros, o quinto metatarso do Dinocephalosaurus era simples e reto em vez de curvo.[2] O Tanystropheus tinha um quinto metatarso ligeiramente engrossado na extremidade superior, e também possuía uma longa falange no quinto dígito que funcionava como outro metatarso.[17] No terceiro dígito do pé do Dinocephalosaurus, havia quatro falanges, mas nenhuma delas parece ter sido a garra terminal [en]. Isso sugere que o Dinocephalosaurus tinha uma contagem acima da média de pelo menos cinco falanges no terceiro dígito.[3]
Classificação

O Dinocephalosaurus foi considerado um membro dos Protorosauria, um grupo de répteis Permo-Triássico ubíquos e diversos.[3] Essa atribuição foi baseada em características como o crânio baixo e estreito com uma região pós-orbital curta e estreita; o osso nasal longo em relação ao osso frontal; a projeção posterior reduzida do jugal; a presença de mais de sete vértebras cervicais, com centros (corpos) mais longos que os das vértebras do tronco; as espinhas neurais baixas das vértebras cervicais; as costelas cervicais longas; a ausência de intercentros articulando com os centros das vértebras do tronco; e a ausência de um forame epicondilar no úmero.[18][19][20][21] Essas características eram compartilhadas com Tanystropheus, Macrocnemus e outros protorossauros.[3]

Os protorossauros foram anteriormente considerados ancestrais dos lagartos,[22] mas análises filogenéticas posteriormente verificaram que eles eram, de fato, arcosauromorfos não arcossauros.[23] Como originalmente definido, o Protorosauria referia-se ao grupo contendo Protorosaurus e Prolacerta [en], e incluía os "Prolacertiformes" como uma subdivisão.[24] Em 1997, Nour-Eddine Jalil conduziu uma análise dos "Prolacertiformes"; essa análise expandiu sua definição para incluir 14 gêneros, incluindo os Tanystropheidae.[18] Pesquisas subsequentes geralmente descobriram que Prolacerta era mais próxima dos Archosauriformes do que Protorosaurus,[25][26] tornando os "Prolacertiformes" não monofiléticos (ou seja, compostos por vários grupos que não eram grupos irmãos).[23][27] Essa pesquisa também sugeriu que os protorossauros restantes – incluindo Protorosaurus, tanistrofeídeos e drepanossauros [en] em alguns estudos – não necessariamente formavam um grupo monofilético. Um número crescente de análises encontrou um Protorosauria não monofilético com Protorosaurus sendo mais basal (menos especializado) do que os Tanystropheidae, e drepanossauros fora dos Archosauromorpha completamente.[23][28][29]
Na descrição original do Dinocephalosaurus, Li reconheceu a região pós-orbital e os centros cervicais alongados como indicativos de uma relação próxima com Tanystropheus, o que o levou a atribuí-lo aos Tanystropheidae.[1] No entanto, Rieppel e colegas posteriormente notaram características que distinguiam o Dinocephalosaurus de protorossauros derivados como Tanystropheus, Macrocnemus, Langobardisaurus e Tanytrachelos; estas incluem a ausência de afunilamento na extremidade frontal do osso nasal e uma contribuição não reduzida do ílio ao acetábulo.[18] Isso levou Rieppel e colegas a considerar o Dinocephalosaurus como um protorossauro indeterminado, com seu alongamento do pescoço sendo convergente ao do Tanystropheus. Sua árvore filogenética, baseada em um conjunto de dados derivado das análises separadas de Jalil (1997),[18] David Dilkes (1998),[21] e Michael Benton & Jackie Allen (1997),[19] é reproduzida abaixo, à esquerda.[3]
|
Topologia A: Rieppel et al. (2008)[3]
|
Topologia B: Liu et al. (2017)[8]
|
|
Liu e colegas publicaram uma análise filogenética separada em 2017. Eles criticaram a análise de Rieppel e colegas por repetir desnecessariamente vários caracteres em seus dados, dando assim peso indevido aos caracteres repetidos na análise. Em sua própria análise, Liu e colegas usaram os mesmos conjuntos de dados de origem, mas excluiram caracteres repetidos, adicionaram dois novos caracteres de uma análise de Sean Modesto e Hans-Dieter Sues (2004),[25] e removeram táxons mal preservados ou potencialmente quiméricos como Cosesaurus, Kadimakara, Trachelosaurus e Malerisaurus. Análises baseadas em máxima parcimônia e métodos bayesianos descobriram que o Dinocephalosaurus era um membro dos Tanystropheidae, sendo o grupo irmão de um grupo monofilético contendo Tanystropheus, Macrocnemus, Langobardisaurus e Tanytrachelos. As análises também recuperaram um Protorosauria monofilético, embora excluindo Prolacerta. A árvore da análise baseada em parcimônia é reproduzida acima, à direita.[8]
Vários estudos subsequentes descobriram que o Dinocephalosaurus era intimamente relacionado, mas fora dos Tanystropheidae. Ezcurra e Butler publicaram uma análise filogenética baseada em um conjunto de dados diferente em 2018, que incluía todos os arcosauromorfos do Permiano Médio ao início do Triássico Superior conhecidos na época. Eles também descobriram que o Dinocephalosaurus estava em uma politomia com Tanystropheidae e Trachelosaurus. Jesairosaurus foi encontrado como o gênero mais próximo dessa politomia.[30] Em 2020, Tiane De-Oliveira e colegas adicionaram Dinocephalosaurus, Jesairosaurus e Elessaurus [en] a outro conjunto de dados publicado por Pritchard e colegas em 2018, enquanto também introduziram modificações baseadas no conjunto de dados de Ezcurra e Butler.[31] Embora sua análise filogenética tenha recuperado uma árvore pouco resolvida com uma grande politomia, eles descobriram que Dinocephalosaurus e Jesairosaurus formavam um grupo basal aos Tanystropheidae e outros arcosauromorfos.[32]

Em 2021, Spiekman e colegas introduziram outro conjunto de dados especificamente para testar as relações filogenéticas dos protorossauros. Diferentes análises foram realizadas usando conjuntos de dados que incorporavam diferentes espécies e características anatômicas. Eles descobriram que Dinocephalosaurus e Pectodens consistentemente formavam um grupo, que eles nomearam como Dinocephalosauridae. Algumas análises (especificamente, aquelas que excluíam caracteres baseados em proporções ou com ordenações) também descobriram que Sclerostropheus e Tanystropheus antiquus caíam dentro desse grupo. Fuyuansaurus ou Jesairosaurus foram encontrados como o parente mais próximo dos dinocephalossaurídeos em algumas análises. Com exceção de Jesairosaurus, esses gêneros de colocação incerta foram encontrados como profundamente aninhados dentro dos Tanystropheidae por outras análises. Fornecendo suporte formal à hipótese original de Rieppel e colegas, Spiekman e colegas descobriram que colocar Dinocephalosaurus como o grupo irmão de Tanystropheus exigia uma árvore que era seis passos mais longa, e portanto menos provável.[13]
Em 2024, Spiekman e colegas publicaram uma redescrição do Dinocephalosaurus, incluindo uma descrição de cinco novos espécimes adicionais. Usando uma versão atualizada do conjunto de dados anterior de Spiekman, eles recuperaram relações filogenéticas semelhantes, com Dinocephalosaurus formando um clado com Pectodens fora dos Tanystropheidae. O seguinte cladograma mostra os resultados de sua análise que incluiu caracteres de proporção e ordenados, excluindo "T. antiquus", T. "conspicuus" e Czatkowiella devido à sua instabilidade:[11]
| Archosauromorpha |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia
Pescoço e alimentação

O pescoço longo do Dinocephalosaurus provavelmente desempenhava um papel funcional. Em particular, o comprimento do pescoço colocava uma longa distância entre a cabeça e o restante do corpo. Isso teria permitido ao Dinocephalosaurus se aproximar de presas potenciais sem que a maior parte de seu volume fosse detectada, o que teria sido eficaz nas águas turvas de seu habitat.[2]
Em 2004, Li e colegas sugeriram que o Dinocephalosaurus também poderia ter usado seu pescoço para capturar presas por meio de alimentação por sucção [en]. Após flexionar o pescoço para o lado (o que teria sido facilitado pela esguiez das costelas cervicais), o ato de endireitar o pescoço teria feito com que as costelas cervicais se espalhassem para fora devido à ação dos músculos do pescoço ligados às costelas. Conforme a cabeça avançava, o volume do esôfago teria aumentado, criando sucção. Uma vez que a presa fosse capturada, os dentes semelhantes a presas teriam segurado a presa na boca.[2] No entanto, em uma resposta, Demes e Krause sugeriram que a alimentação por sucção teria envolvido o animal engolir uma grande quantidade de água salgada, bem como expandir rapidamente seu esôfago. Sem evidências de adaptações para nenhum dos dois comportamentos (glândulas de sal para o primeiro, um grande osso hioide ou costelas especializadas para o segundo), eles consideraram a sugestão de Li e colegas improvável.[33] No entanto, como outros amniotas aquáticos, o Dinocephalosaurus teria engolido e digerido sua presa de cabeça primeiro, como evidenciado pela preservação de um peixe perleidídeo na região abdominal de LPV 30280 de Luoping.[8][34]
Não é provável que o Dinocephalosaurus usasse seu pescoço longo para respirar estendendo-o verticalmente. Se tentasse fazer isso, a diferença de pressão entre a superfície e seu torso seria suficientemente extrema para que seus pulmões não se inflassem. Assim, para respirar, o Dinocephalosaurus precisaria se aproximar da superfície com o pescoço quase horizontal. A postura horizontal do pescoço também teria facilitado a locomoção na superfície, devido ao seu longo perfil aumentando seu "comprimento do casco" e reduzindo o efeito da resistência das ondas.[2]
Reprodução

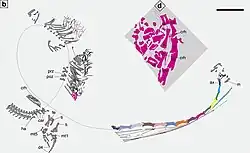
O Dinocephalosaurus representa o ápice das adaptações aquáticas entre os protorossauros.[2] Dado seu pescoço longo e membros semelhantes a nadadeiras, provavelmente era incapaz de funcionar confortavelmente em um ambiente terrestre. Em 2021, Motani e Vermeij categorizaram isso como o quarto em uma sequência de cinco etapas de adaptações crescentes a ambientes marinhos com base em animais modernos como análogos, enquanto os tanistrofeídeos estavam apenas na segunda etapa (alimentação no oceano).[35] Uma consequência dessas adaptações é que o Dinocephalosaurus seria incapaz de construir ninhos em terra. Isso também o teria impedido de possuir ovos reptilianos de casca dura; tais ovos necessitam da troca de gases com o ambiente externo, e esse processo é significativamente mais lento na água do que no ar.[36] Assim, o Dinocephalosaurus não poderia ter sido ovíparo. Ao mesmo tempo, no entanto, os arcossauros são conhecidos pela ausência total de viviparidade, ou nascimento vivo, entre seus membros vivos.[8][37]

O indivíduo embrionário do Dinocephalosaurus preservado dentro de LPV 30280 de Luoping pode ser identificado como tal por várias razões. Primeiro, ele está completamente contido na cavidade corporal do adulto. Suas costelas cervicais – que são longas, como as do adulto – estão voltadas para a mesma direção que as vértebras dorsais do adulto, o que está em contradição com o método típico de engolir presas de cabeça primeiro entre os amniotas. Ele também está preservado com seu pescoço curvado em direção aos membros anteriores, uma postura vista entre embriões de vertebrados, mas não em espécimes adultos de Dinocephalosaurus, que geralmente têm o pescoço curvado para cima.[3] A ausência de mãos associadas aos membros anteriores no embrião pode ser um artefato de preservação, mas também pode ser devido à sequência na qual os ossos dos membros dos tetrapodes ossificam.[8]
Embora a presença de um embrião per se seja equívoca em relação a essa questão, as condições em que o embrião foi preservado fornecem forte evidência de que o Dinocephalosaurus era vivíparo. Não há uma casca de ovo calcificada preservada ao redor do embrião, apesar da presença de elementos calcificados delicadamente preservados de outros animais em Luoping.[4] Isso é consistente com as cascas de ovos envolvendo os embriões de répteis vivíparos sendo membranas não calcificadas.[38] Além disso, as proporções relativas do úmero e da fíbula no embrião, comparadas com o indivíduo materno e IVPP V13898 de Panxian, indicam que o embrião tem cerca de 12% do tamanho de sua mãe. Combinado com seus ossos ossificados, isso sugere que o embrião estava em um estágio avançado de desenvolvimento, enquanto crocodilianos, pássaros, tartarugas e tuataras depositam ovos em estágios de desenvolvimento muito iniciais.[8][39]
No geral, as evidências fornecidas pelo embrião sugerem que o Dinocephalosaurus era vivíparo, tornando-o o primeiro arcosauromorfo vivíparo[8] exceto, possivelmente, pelos metriorrinquídeos.[40] Isso é consistente com a separação de suas costelas sacrais do sacro,[3] o que indica uma pelve móvel que poderia ter funcionado no parto.[41][42] Embora a ausência de arcossauros vivíparos tenha sido historicamente atribuída a atributos comuns herdados da linhagem-tronco arcossauriana, a descoberta de que o Dinocephalosaurus era vivíparo sugere que esse fenômeno é devido a restrições específicas de estilo de vida da linhagem. Embora os sistemas de determinação de sexo entre os arcosauromorfos vivos sejam diversos, com crocodilianos e tartarugas usando determinação de sexo dependente de temperatura,[43] modelagens filogenéticas sugerem que o Dinocephalosaurus manteve a condição basal de determinação genotípica do sexo dos primeiros diapsídeos, e que esse sistema facilitou sua transição para um estilo de vida obrigatoriamente marinho, juntamente com a viviparidade.[8][44]
Paleoecologia
Pelo menos até o final do Triássico Médio, os altos níveis do mar permitiam que águas rasas cobrissem grande parte do Bloco do Sul da China,[45][46] uma placa tectônica que hoje consiste no estável Cráton do Yangtze e no menos estável Fold Belt do Sul da China. Um evento de construção de montanhas conhecido como orogenia Indosiniana elevou rochas pré-cambrianas para formar quatro grandes massas de terra no Bloco do Sul da China: Khamdian a oeste, Jiangnan no centro, Yunkai ao sul e Cathaysia a leste. As cadeias de ilhas também se estendiam entre Yunkai e Cathaysia, a leste. Localizada entre Khamdian e Jiangnan, havia uma profunda bacia oceânica conhecida como Bacia de Nanpanjiang.[46][47][48] Ao longo da borda ocidental dessa bacia, sedimentos contendo fósseis foram depositados para se tornarem o que hoje são os Lagerstätte (depósitos sedimentares caracterizados por preservação exemplar) de Panxian, Luoping e outras localidades.[4]

Em Panxian, parece ter havido uma transição na fauna endêmica de répteis marinhos, possivelmente causada pelo vulcanismo. Esse vulcanismo é indicado pela presença de uma camada de bentonita entre os horizontes médio e superior de répteis.[49] Na parte inferior do horizonte superior de répteis, também foram encontrados fósseis do onipresente ictiossauro mixossauro [en] Mixosaurus panxianensis (que ocorre em todas as camadas), dos paquipleurossauros Keichousaurus sp. e Wumengosaurus delicatomandibularis e de peixes, embora os fósseis deste último sejam fragmentários. Os horizontes Inferior Médio e Inferior de répteis também incluem o mixossauro Phalarodon [en] cf. fraasi, o ictiossauro primitivo Xinminosaurus catactes, o placodonte Placodus inexpectatus, os notossauros Lariosaurus [en] hongguoensis e Nothosaurus yangjuanensis, e o arcossauro Qianosuchus mixtus, além de bivalves e peixes saurictiídeos.[4][50]
A fauna de Luoping parece ter sido preservada em uma pequena bacia intraplataforma, em vez de em águas abertas ao redor, a julgar pelos sedimentos anóxicos presentes no local.[4] Dos 19.759 espécimes, 93,7% dos fósseis encontrados em Luoping são artrópodes: decápodes, isópodes, ciclóides semelhantes a caranguejos, misidáceos semelhantes a camarões e ostracodes, bem como raros milípedes e caranguejos-ferradura. Em comparação, apenas 0,07% dos espécimes provêm de répteis marinhos,[51] que incluem o Dinocephalosaurus e o Pectodens, juntamente com os mixossaurianos Mixosaurus cf. panxianensis e Phalarodon atavus; os paquipleurossauros Dianmeisaurus gracilis e Dianopachysaurus dingi; os saurosphargídeos [en] Largocephalosaurus polycarpon e Sinosaurosphargis yunguiensis; os notossauros Nothosaurus zhangi e uma espécie de Lariosaurus; outros sauropterígeos Atopodentatus unicus, Dawazisaurus brevis e Diandongosaurus acutidentatus; e um arcossauro relacionado a Qianosuchus.[4][52]
Por outro lado, peixes como saurichthyiformes, palaeoniscídeos, birgerídeos, perleidídeos, eugnathídeos, semionotiformes, pholidopleurídeos, peltopleurídeos e coelacanthiformes foram encontrados em Luoping, formando 3,66% dos fósseis com 25 taxa em 9 famílias. Moluscos como bivalves e gastrópodes, juntamente com amonoides e belemnoides, representam 1,69% dos espécimes. Os fósseis raros e provavelmente não endêmicos também incluem equinodermos, como crinoides, estrelas-do-mar e ouriços-do-mar; branquiópodes; e ramos e folhas de coníferas relativamente completos, que provavelmente se originaram de florestas costeiras a menos de 10 km de distância.[51]
Referências
- ↑ a b c d e f g Li, C. (2003). «Primeiro registro de réptil protorossaurídeo (Ordem Protorosauria) do Triássico Médio da China» 4 ed. Acta Geologica Sinica - English Edition. 77: 419–423. doi:10.1111/j.1755-6724.2003.tb00122.x
- ↑ a b c d e f g h Li, C.; Rieppel, O.; LaBarbera, M.C. (2004). «Um Protorossauro Aquático Triássico com um Pescoço Extremamente Longo» 5692 ed. Science. 305: 1931. PMID 15448262. doi:10.1126/science.1100498
- ↑ a b c d e f g h i j k l m n o p Rieppel, O.; Li, C.; Fraser, N.C (2008). «A Anatomia Esquelética do Protorossauro Triássico Dinocephalosaurus orientalis Li, do Triássico Médio da Província de Guizhou, Sul da China» 1 ed. Journal of Vertebrate Paleontology. 28: 95–110. JSTOR 30126337. doi:10.1671/0272-4634(2008)28[95:TSAOTT]2.0.CO;2
- ↑ a b c d e f g h Benton, M.J.; Zhang, Q.; Hu, S.; Chen, Z.-Q.; Wen, W.; Liu, J.; Zhou, C.; Xie, T.; Tong, J.; Choo, B. (2013). «Biota Vertebrada Excepcional do Triássico da China e a Expansão dos Ecossistemas Marinhos Após a Extinção em Massa Permo-Triássica». Earth-Science Reviews. 123: 199–243. Bibcode:2013ESRv..125..199B. doi:10.1016/j.earscirev.2013.05.014
- ↑ Sun, Z.; Sun, Y.; Hao, W.; Jiang, D. (2006). «Evidências de Conodontes para a Idade da Fauna de Panxian, Guizhou, China» 5 ed. Acta Geologica Sinica - English Edition. 80: 621–630. doi:10.1111/j.1755-6724.2006.tb00284.x
- ↑ Wang, Y.; Yang, D.; Han, J.; Wang, L.; Yao, J.; Liu, D. (2014). «A Idade U-Pb Triássica para o Protorossauro Aquático de Pescoço Longo de Guizhou, China» 4 ed. Geological Magazine. 151: 749–754. doi:10.1017/S001675681400003X
- ↑ Zhang, Q.-Y.; Zhou, C.-Y.; Lu, T.; Xie, T.; Lou, X.-Y.; Liu, W.; Sun, Y.-Y.; Huang, J.-Y.; Zhao, L.-S. (2009). «Uma Atribuição de Idade do Triássico Médio Baseada em Conodontes para a Biota de Luoping, Yunnan, China» 10 ed. Science in China Series D: Earth Sciences. 52: 1673–1678. Bibcode:2009ScChD..52.1673Z. doi:10.1007/s11430-009-0114-z
- ↑ a b c d e f g h i j k Liu, J.; Organ, C.L.; Benton, M.J.; Brandley, M.C.; Aitchison, J.C. (2017). «Nascimento Vivo em um Réptil Arcosauromorfo». Nature Communications. 8. 14445 páginas. Bibcode:2017NatCo...814445L. ISSN 2041-1723. PMC 5316873
 . PMID 28195584. doi:10.1038/ncomms14445
. PMID 28195584. doi:10.1038/ncomms14445
- ↑ a b c Fraser, N.C.; Rieppel, O.; L., C. (2013). «Um Protorossauro de Focinho Longo do Triássico Médio do Sul da China» 5 ed. Journal of Vertebrate Paleontology. 33: 1120–1126. doi:10.1080/02724634.2013.764310
- ↑ Li, C.; Rieppel, O.; Fraser, N.C. (2017). «Viviparidade em um Réptil Arcosauromorfo Marinho Triássico» 3 ed. Vertebrata PalAsiatica. 55: 210–217. Consultado em 2 de setembro de 2025
- ↑ a b Spiekman, Stephan N. F.; Wang, Wei; Zhao, Lijun; Rieppel, Olivier; Fraser, Nicholas C.; Li, Chun (23 de fevereiro de 2024). «Dinocephalosaurus orientalis Li, 2003: um Notável Arcosauromorfo Marinho do Triássico Médio do Sudoeste da China». Earth and Environmental Science Transactions of the Royal Society of Edinburgh (em inglês): 1–33. ISSN 1755-6910. doi:10.1017/S175569102400001X
- ↑ Spiekman, Stephan N. F.; Scheyer, Torsten M. (16 de dezembro de 2019). «Uma Revisão Taxonômica do Gênero Tanystropheus (Archosauromorpha, Tanystropheidae)» 3 ed. Palaeontologia Electronica (em inglês). 22: 1–46. ISSN 1094-8074. doi:10.26879/1038. Consultado em 2 de setembro de 2025
- ↑ a b c d e f Spiekman, S.N.F.; Fraser, N.C.; Scheyer, T.M. (2021). «Uma Nova Hipótese Filogenética dos Tanystropheidae (Diapsida, Archosauromorpha) e Outros "Protorossauros", e suas Implicações para a Evolução Inicial dos Arcossauros-Tronco». PeerJ. 9: e11143. PMC 8101476. PMID 33986981. doi:10.7717/peerj.11143
- ↑ Rieppel, O.; Jiang, D.-Y.; Fraser, N.C.; Hao, W.-C.; Motani, R.; Sun, Y.-L.; Sun, Z.-Y. (2010). «Tanystropheus cf. T. longobardicus do Início do Triássico Superior de Guizhou, Sudoeste da China» 4 ed. Journal of Vertebrate Paleontology. 30: 1082–1089. doi:10.1080/02724634.2010.483548
- ↑ Rieppel, O. (2001). «Uma Nova Espécie de Tanystropheus (Reptilia: Protorosauria) do Triássico Médio de Makhtesh Ramon, Israel» 2 ed. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 221: 271–287. doi:10.1127/njgpa/221/2001/271
- ↑ Renesto, S. (2005). «Um Novo Espécime de Tanystropheus (Reptilia Protorosauria) do Triássico Médio da Suíça e a Ecologia do Gênero» 3 ed. Rivista Italiana di Paleontologia e Stratigrafia. 111: 377–394. doi:10.13130/2039-4942/6327
- ↑ Renesto, S. (1994). «Um Novo Réptil Prolacertiforme do Triássico Superior do Norte da Itália» 2 ed. Rivista Italiana di Paleontologia e Stratigrafia. 100: 285–306. doi:10.13130/2039-4942/8615
- ↑ a b c d Jalil, N.-E. (1997). «Um Novo Diápsido Prolacertiforme do Triássico do Norte da África e as Inter-relações dos Prolacertiformes» 3 ed. Journal of Vertebrate Paleontology. 17: 506–525. doi:10.1080/02724634.1997.10010998
- ↑ a b Benton, M.J.; Allen, J.L. (1997). «Boreopricea do Triássico Inferior da Rússia e as Relações dos Répteis Prolacertiformes». Palaeontology. 40: 931–953. Consultado em 2 de setembro de 2025
- ↑ Evans, S.E. (1988). «A História Inicial e as Relações dos Diápsidos». In: Benton, M.J. A Filogenia e Classificação dos Tetrapodes. 1. Oxford: Clarendon Press. pp. 221–253
- ↑ a b Dilkes, D.W. (1998). «O Rincossauro Triássico Inicial Mesosuchus browni e as Inter-relações dos Répteis Arcosauromorfos Basais» 1368 ed. Philosophical Transactions of the Royal Society B. 353: 501–541. PMC 1692244. doi:10.1098/rstb.1998.0225
- ↑ Parrington, F.R. (1935). «Sobre Prolacerta broomi, gen. et sp. n., e a Origem dos Lagartos» 92 ed. Annals and Magazine of Natural History. 10. 16: 197–205. doi:10.1080/00222933508655037
- ↑ a b c Ezcurra, M.D. (2016). «As Relações Filogenéticas dos Arcosauromorfos Basais, com Ênfase na Sistemática dos Arcossauriformes Proterossuquianos». PeerJ. 4: e1778. PMC 4860341. PMID 27162705. doi:10.7717/peerj.1778
- ↑ Camp, C.L. (1945). «Prolacerta e os Répteis Protorossaurídeos; Parte I» 1 ed. American Journal of Science. 243: 17–32. Bibcode:1945AmJS..243...17C. doi:10.2475/ajs.243.1.17
- ↑ a b Modesto, S.P.; Sues, H.D. (2004). «O Crânio do Réptil Arcosauromorfo Triássico Inicial Prolacerta broomi e sua Significância Filogenética» 3 ed. Zoological Journal of the Linnean Society. 140: 335–351. doi:10.1111/j.1096-3642.2003.00102.x. Consultado em 2 de setembro de 2025
- ↑ Gottmann-Quesada, A.; Sander, P.M. (2009). «Uma Redescrição do Arcosauromorfo Inicial Protorosaurus speneri Meyer, 1832, e suas Relações Filogenéticas» 4–6 ed. Palaeontographica Abteilung A. 287: 123–200. doi:10.1127/pala/287/2009/123
- ↑ Ezcurra, M.D.; Scheyer, T.M.; Butler, R.J. (2014). «A Origem e Evolução Inicial dos Sauria: Reavaliando o Registro Fóssil Sauriano do Permiano e o Momento da Divergência Crocodilo-Lagarto» 5 ed. PLOS ONE. 9: e89165. Bibcode:2014PLoSO...989165E. PMC 3937355. PMID 24586565. doi:10.1371/journal.pone.0089165
- ↑ Pritchard, A.C; Turner, A.H.; Nesbitt, S.J.; Irmis, R.B.; Smith, N.D. (2015). «Tanistrofeídeos do Triássico Superior (Reptilia, Archosauromorpha) do Norte do Novo México (Membro da Floresta Petrificada, Formação Chinle) e a Biogeografia, Morfologia Funcional e Evolução dos Tanystropheidae» 2 ed. Journal of Vertebrate Paleontology. 35: e911186. doi:10.1080/02724634.2014.911186
- ↑ Pritchard, A.C.; Nesbitt, S.J. (2017). «Um Crânio Semelhante ao de Pássaro em um Réptil Diápsido Triássico Aumenta a Heterogeneidade da Radiação Morfológica e Filogenética dos Diápsidos» 10 ed. Royal Society Open Science. 4. 170499 páginas. Bibcode:2017RSOS....470499P. ISSN 2054-5703. PMC 5666248. PMID 29134065. doi:10.1098/rsos.170499
- ↑ Ezcurra, M.D.; Butler, R.J. (2018). «A Ascensão dos Répteis Dominantes e a Recuperação do Ecossistema da Extinção em Massa Permo-Triássica» 1880 ed. Proceedings of the Royal Society B. 285: 20180361. PMC 6015845. PMID 29899066. doi:10.1098/rspb.2018.0361
- ↑ Pritchard, A.C.; Gauthier, J.A.; Hanson, M.; Bever, G.S.; Bhullar, B.-A. S. (2018). «Um Sauriano Triássico Minúsculo de Connecticut e a Evolução Inicial do Aparelho de Alimentação dos Diápsidos» 1 ed. Nature Communications. 9: 1213. PMC 5865133. PMID 29572441. doi:10.1038/s41467-018-03508-1
- ↑ De-Oliveira, T.M.; Pinheiro, F.L.; Stock Da-Rosa, A.A.; Dias-Da-Silva, S.; Kerber, L. (2020). «Um Novo Arcosauromorfo da América do Sul Fornece Insights sobre a Diversificação Inicial dos Tanistrofeídeos» 5 ed. PLOS ONE. 15: e0233216. PMC 7213688. PMID 32392264. doi:10.1371/journal.pone.0233216
- ↑ Demes, B.; Krause, D.W. (2005). «Alimentação por Sucção em um Protorossauro Triássico?» 5725 ed. Science. 308: 1112–1113. doi:10.1126/science.308.5725.1112c
- ↑ Caldwell, M.W.; Lee, M.S.Y. (2001). «Nascimento Vivo em Lagartos Marinhos Cretáceos (Mosassauróides)» 1484 ed. Proceedings of the Royal Society B. 268: 2397–240. PMC 1088892. PMID 11747556. doi:10.1098/rspb.2001.1796
- ↑ Motani, R.; Vermeij, G.J. (2021). «Etapas Ecofisiológicas de Adaptação Marinha em Tetrapodes Não Avianos Extintos e Existentes» 5 ed. Biological Reviews. 96: 1769–1798. PMID 33904243. doi:10.1111/brv.12724. Consultado em 2 de setembro de 2025
- ↑ Shine, R. (1985). «A Evolução da Viviparidade em Répteis: Uma Análise Ecológica». In: Gans, C.; Billett, F. Biologia dos Reptilia. 15. Nova York: John Wiley & Sons. pp. 605–694. Consultado em 2 de setembro de 2025
- ↑ Blackburn, D.G.; Evans, H.E. (1986). «Por que não há pássaros vivíparos?» 2 ed. The American Naturalist. 128: 165–190. JSTOR 2461543. doi:10.1086/284552
- ↑ Stewart, J.R.; Thompson, M.B. (2009). «Evolução Paralela da Placentação em Lagartos Scincídeos Australianos» 6 ed. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 312B: 590–602. PMID 19012323. doi:10.1002/jez.b.21245
- ↑ Andrews, R.M. (2004). «Desenvolvimento Embrionário». In: Deeming, D.C. Incubação Reptiliana: Ambiente, Evolução e Comportamento. Nottingham: Nottingham University Press. pp. 75–102
- ↑ Herrera, Y.; Fernandez, M.S.; Lamas, S.G.; Campos, L.; Talevi, M.; Gasparini, Z. (2017). «Morfologia da Região Sacral e Estratégias Reprodutivas dos Metriorhynchidae: Uma Abordagem Contra-indutiva» (PDF) 4 ed. Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 106: 247–255. doi:10.1017/S1755691016000165. Consultado em 2 de setembro de 2025
- ↑ Cheng, Y.-N.; Wu, X.-C.; Ji, Q. (2004). «Répteis Marinhos Triássicos Deram à Luz Filhotes Vivos» (PDF) 7015 ed. Nature. 432: 383–386. Bibcode:2004Natur.432..383C. PMID 15549103. doi:10.1038/nature03050. Consultado em 2 de setembro de 2025
- ↑ O'Keefe, F.R.; Chiappe, L.M. (2011). «Viviparidade e História de Vida K-Selecionada em um Plesiossauro Marinho Mesozóico (Reptilia, Sauropterygia)» 6044 ed. Science. 333: 870–873. Bibcode:2011Sci...333..870O. PMID 21836013. doi:10.1126/science.1205689
- ↑ Organ, C.L.; Janes, D.E. (2008). «Evolution of sex chromosomes in Sauropsida». Integrative and Comparative Biology. 48 (4): 512–519. PMC 4553705. PMID 21669812. doi:10.1093/icb/icn041
- ↑ Organ, C.L.; Janes, D.E.; Meade, A.; Pagel, M. (2009). «Genotypic sex determination enabled adaptive radiations of extinct marine reptiles». Nature. 461 (7262): 389–392. Bibcode:2009Natur.461..389O. PMID 19759619. doi:10.1038/nature08350
- ↑ Lehrmann, D.J.; Enos, P.; Payne, J.L.; Montgomery, P.; Wei, J.; Yu, Y.; Xiao, J.; Orchard, M.J. (2005). «Permian and Triassic depositional history of the Yangtze platform and Great Bank of Guizhou in the Nanpanjiang basin of Guizhou and Guangxi, South China» (PDF). Albertiana. 33: 149–168
- ↑ a b Lehrmann, D.J.; Minzoni, M.; Enos, P.; Yu, Y.-Y.; Wei, J.-Y.; Li, R.-X. (2009). «Triassic depositional history of the Yangtze platform and Great Bank of Guizhou in the Nanpanjiang basin of South China». Journal of Earth Sciences and Environment. 31 (4): 344–367
- ↑ Lehrmann, D.J.; Payne, J.L.; Felix, S.V.; Dillett, P.M.; Wang, H.; Yu, Y.; Wei, J. (2003). «Permian–Triassic Boundary Sections from Shallow-Marine Carbonate Platforms of the Nanpanjiang Basin, South China: Implications for Oceanic Conditions Associated with the End-Permian Extinction and Its Aftermath». PALAIOS. 18 (2): 138–152. Bibcode:2003Palai..18..138L. CiteSeerX 10.1.1.486.1486. doi:10.1669/0883-1351(2003)18<138:PBSFSC>2.0.CO;2
- ↑ Lehrmann, D.J.; Payne, J.L.; Pei, D.; Enos, P.; Druke, D.; Steffen, K.; Zhang, J.; Wei, J.; Orchard, M.J.; Ellwood, B. (2007). «Record of the end-Permian extinction and Triassic biotic recovery in the Chongzuo-Pingguo platform, southern Nanpanjiang basin, Guangxi, South China». Palaeogeography, Palaeoclimatology, Palaeoecology. 252 (1): 200–217. Bibcode:2007PPP...252..200L. doi:10.1016/j.palaeo.2006.11.044
- ↑ Motani, R.; Jiang, D.-Y.; Tintori, A.; Sun, Y.-L.; Hao, W.-C.; Boyd, A.; Sanja, H.-F.; Schmitz, L.; Shin, Y.-J.; Sun, Z.-Y. (2008). «Horizons and Assemblages of Middle Triassic Marine Reptiles from Panxian, Guizhou, China». Journal of Vertebrate Paleontology. 28 (3): 900–903. doi:10.1671/0272-4634(2008)28[900:HAAOMT]2.0.CO;2
- ↑ Jiang, D.; Motani, R.; Hao, W.; Rieppel, O.; Sun, Y.; Tintori, A.; Sun, Z.; Schmitz, L. (2009). «Biodiversity and Sequence of the Middle Triassic Panxian Marine Reptile Fauna, Guizhou Province, China». Acta Geologica Sinica - English Edition. 83 (3): 451–459. doi:10.1111/j.1755-6724.2009.00047.x
- ↑ a b Hu, S.-X.; Zhang, Q.-Y.; Chen, Z.-Q.; Zhou, C.-Y.; Lü, T.; Xie, T.; Wen, W.; Huang, J.-Y.; Benton, M.J. (2011). «The Luoping biota: exceptional preservation, and new evidence on the Triassic recovery from end-Permian mass extinction». Proceedings of the Royal Society B. 278 (1716): 2274–2282. PMC 3119007. PMID 21183583. doi:10.1098/rspb.2010.2235
- ↑ Liu, Q.; Yang, T.; Cheng, L.; Benton, M.J.; Moon, B.C.; Yan, C.; An, Z.; Tian, L. (2021). «An injured pachypleurosaur (Diapsida: Sauropterygia) from the Middle Triassic Luoping Biota indicating predation pressure in the Mesozoic». Scientific Reports. 11 (1): 21818. PMC 8575933. PMID 34750442. doi:10.1038/s41598-021-01309-z
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||