Caliciaceae
Caliciaceae
| |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Calicium viride | |||||||||||||
| Classificação científica | |||||||||||||
| |||||||||||||
| Género-tipo | |||||||||||||
| Calicium Pers. (1794) | |||||||||||||
| Sinónimos[1] | |||||||||||||
| |||||||||||||
Caliciaceae é uma família de fungos predominantemente formadores de líquens, pertencente à classe Lecanoromycetes, na divisão Ascomycota. Embora a classificação da família tenha sofrido diversas alterações ao longo de sua história taxonômica, o uso de métodos modernos de filogenia molecular consolidou sua posição atual na ordem Caliciales. A família Caliciaceae abrange 39 gêneros e cerca de 670 espécies. O gênero mais numeroso é Buellia [en], com aproximadamente 300 espécies; mas também há mais de uma dúzia de gêneros que contêm apenas uma única espécie.
A maioria das espécies de Caliciaceae cresce sobre cascas de árvores, madeira morta ou rochas. Algumas espécies, particularmente do gênero-tipo Calicium, caracterizam-se pela presença de ascos de paredes finas e de curta duração, além de um mazaédio, que é um acúmulo de esporos maduros e soltos que cobrem a superfície do corpo frutífero. Esse mecanismo de dispersão passiva de esporos é relativamente raro entre os Ascomycota. O mazaédio, geralmente preto, pode estar situado no topo de um longo pedúnculo fino ou repousar sobre (ou, por vezes, estar imerso em) a superfície do substrato do líquen. Muitas outras espécies de Caliciaceae produzem esporos em um apotécio, que geralmente se assemelha a um disco preto achatado.
A família inclui espécies com formas de crescimento crustosas (semelhantes a crostas), folhosas (semelhantes a folhas) e, em alguns casos, fruticosas (semelhantes a arbustos). O parceiro fotobionte dos líquens da Caliciaceae é geralmente do gênero de algas verdes Trebouxia. Coletivamente, a família tem uma distribuição cosmopolita, sendo encontrada em todos os continentes habitados. Embora a família seja mais representada em áreas montanhosas de regiões temperadas e tropicais, algumas espécies resistentes conseguem sobreviver ao ambiente hostil dos Vales Secos de McMurdo na Antártica, crescendo em fendas na superfície de rochas. Cinco espécies da Caliciaceae estão incluídas na Lista Vermelha de Espécies Ameaçadas da União Internacional para a Conservação da Natureza. Algumas espécies resistentes à poluição atmosférica do gênero Pyxine foram investigadas para uso como bioindicadores de poluição por metais pesados. Diversos fósseis de Caliciaceae encontrados em inclusões de âmbar foram utilizados para avaliar a história evolutiva da família.
Sistemática
A família Caliciaceae foi circunscrita em 1826 pelo botânico francês François Fulgis Chevallier. Ele descreveu a família: "As Calicineas são um pequeno grupo de plantas cujo crescimento confundiu os botânicos. São pequenos fungos parasitas nas crostas de líquens, principalmente em Variola e porina comum, e em galhos mortos."[2] O gênero-tipo da Caliciaceae é Calicium, originalmente circunscrito em 1794 por Christiaan Hendrik Persoon; este gênero é, por sua vez, tipificado por Calicium viride.[3]
As famílias Buelliaceae e Pyxinaceae são famílias históricas criadas para conter taxa que agora estão incluídos na Caliciaceae.[1] Outras duas famílias 'calicioides' com "Caliciaceae" em seus nomes – Microcaliciaceae (ordem Pertusariales, classe Lecanoromycetes) e Mycocaliciaceae (ordem Mycocaliciales, classe Eurotiomycetes) – contêm espécies anteriormente consideradas estreitamente relacionadas à Caliciaceae. O gênero Microcaliciaceae inclui espécies calicioides não liquenizadas que são parasitas de líquens ou colônias de algas de vida livre, enquanto a Mycocaliciaceae contém fungos calicioides não liquenizados que não possuem mazaédio e utilizam dispersão ativa de esporos.[4]
Classificação
Os Caliciaceae e outros líquens piniformes morfologicamente semelhantes, com ascomas formadores de mazaédio e dispersão passiva de esporos, costumavam ser agrupados na ordem Caliciales, que por muito tempo foi considerada um exemplo modelo de agrupamento monofilético de taxa.[5] Historicamente, a família também foi colocada por diversos autores nas ordens Coryneliales, Helotiales e na agora obsoleta Sphaeriales, dependendo de quais características fenotípicas eram consideradas mais importantes.[6]
O liquenologista sueco Leif Tibell dedicou grande parte de sua carreira ao estudo de líquens calicioides, utilizando diversas técnicas para compreender as relações entre os taxa desse grupo,[7] incluindo análises fenéticas e cladísticas de morfologia e investigações de metabólitos secundários. Em 1984, ele sugeriu que a ordem era polifilética (ou seja, não originada de um único ancestral comum) e que as principais características identificadoras dos Caliciales haviam evoluído independentemente em vários grupos não relacionados.[8] Ele restringiu a ordem a três famílias (Caliciaceae, Mycocaliciaceae e Sphinctrinaceae) que considerava o "núcleo" do grupo.[8][9] Análises filogenéticas preliminares mostraram que Mycocaliciaceae e Sphinctrinaceae pertenciam à classe Eurotiomycetes, enquanto a Caliciaceae parecia se agrupar com a ordem Lecanorales.[5] A proposição de que fungos mazaediados estão dispersos pelos Ascomycota foi confirmada posteriormente em vários estudos filogenéticos moleculares.[4][5][10] A proposição de que os fungos caliciformes estão dispersos em toda a Ascomycota foi confirmada posteriormente em vários estudos filogenéticos moleculares. Os seis gêneros que foram incluídos por Tibell na família Caliciaceae em sua classificação proposta de 1984 para os fungos caliciformes (Acroscyphus, Calicium, Cyphelium, Texosporium, Thelomma e Tholurna) ainda estão na família atualmente.[11]
Filogenética
As famílias Caliciaceae e Physciaceae foram demonstradas como estreitamente relacionadas em estudos moleculares.[12][13] Ambas as famílias foram temporariamente colocadas no grupo dos Teloschistales como parte da subordem Physciineae.[14] Como as espécies produtoras de mazaédio dos Caliciaceae estavam aninhadas nos gêneros Dirinaria, Pyxine e Physcia (todos anteriormente contidos na família Physciaceae), alguns autores trataram todos os membros de Caliciaceae e Physciaceae como uma única família, e o nome Physciaceae foi proposto para conservação em 2002.[15] Com o surgimento de estudos filogenéticos adicionais desde então, no entanto, um conceito de duas famílias, Caliciaceae e Physciaceae, tem sido preferido pela maioria das autoridades. Em 2012, a subordem Physciineae foi elevada ao status de ordem, e o nome Caliciales foi ressuscitado.[16] Na sua circunscrição moderna, a ordem Caliciales contém essas duas famílias.[17]
Um grande estudo molecular do clado Caliciaceae-Physciaceae publicado em 2016 ajudou a esclarecer as relações naturais nesse grupo e a definir melhor as delimitações genéticas.[18] O gênero Culbersonia, anteriormente classificado dentro do grupo Physciaceae devido à sua semelhança morfológica com Physconia, foi demonstrado como membro dos Caliciaceae em 2019. Em análises filogenéticas, ele se agrupa em um clado com Pyxine e Dirinaria. Esses três gêneros distinguem-se de outros Caliciaceae pela forma de crescimento folhoso apressada (como pequenas folhas pressionadas contra o substrato), pela ausência de mazaédio e, ecologicamente, por sua predominância em regiões subtropicais e tropicais.[11]
Descrição

.jpg)
O talo, quando presente, é crustoso, com textura verrucosa (semelhante a verrugas), granular (semelhante a grãos) ou areolado. Sua cor é tipicamente cinza, amarela ou esverdeada. Às vezes, está imerso nas camadas externas da casca de seu hospedeiro. Os ascomas são mais ou menos esféricos ou hemisféricos, situados no topo de um longo pedúnculo em algumas espécies, mas sésseis em outras. Quando presente, o pedúnculo é geralmente melanizado (escurecido).[19] A forma dos ascomas é apoteciada (constituída por um apotécio) ou mazaediada.[1]
O apotécio consiste nos ascos (células portadoras de esporos) e no himênio associado, além da parte estéril, estrutural e de suporte do apotécio, chamada excípulo. Em espécies apoteciadas de Caliciaceae, o apotécio é tipicamente lecidino (sem margem ao redor do talo ou com margem formada pelo próprio excípulo), em vez de lecanorino (arredondado com margem proeminente). A estrutura tecidual do excípulo é prosoplectenquimatosa (composta por hifas longas, estreitas, onduladas e paralelas) ou paraplectenquimatosa (um arranjo celular onde as hifas estão orientadas em todas as direções) e é hialina (translúcida) a marrom-escura.[1] O gênero Acroscyphus, que contém a única espécie rara, mas amplamente distribuída, A. sphaerophoroides, é uma exceção peculiar à morfologia típica dos Caliciaceae: possui um talo dactiliforme (semelhante a dedos), ascocarpos imersos em podécios (pedúnculos ocos) e uma medula amarela a laranja.[20]
Os ascos das espécies de Caliciaceae são cilíndricos a amplamente claviformes. Eles estão fixados a croziers, que são estruturas em forma de gancho nas células que se formam na base dos ascos. Os ascos têm paredes finas e não possuem estruturas internas na ponta. Como resultado, duram pouco tempo antes de se degradarem, e os ascosporos dentro deles são liberados passivamente em uma massa mazaedial.[19] O hamatécio (termo que se refere aos tecidos entre os ascos no himênio) consiste em paráfises não ramificadas ou ligeiramente ramificadas que são amiloides.[1] Geralmente, há oito esporos por asco, embora às vezes esse número seja reduzido a quatro ou aumentado para 16–32 por asco.[1] Os ascosporos são marrom-escuros, com superfície lisa ou ornamentada com remanescentes da ruptura das camadas externas da parede. Eles possuem zero ou um septo.[19] Os esporos dos líquens Texosporium têm uma ornamentação única criada por paráfises que aderem à superfície; essa característica pode ajudar a proteger contra dessecação ou danos ao DNA causados por radiação em habitats ensolarados.[21] Os conidiomas produzidos pelas espécies de Caliciaceae têm a forma de picnídios. Os conídios não têm septo, têm formato de bastonete (bacilar) a filiforme (filiforme),geralmente curvado, e são hialinos.[1]

A química secundária das espécies de Caliciaceae é variável. Compostos comumente relatados na família incluem depsides [en] (como atranorina), terpenos, depsidonas (por exemplo, ácido norstíctico) e lichexantonas. Às vezes, antraquinonas estão presentes quando o talo é pigmentado.[1] O gênero Acroscyphus é novamente uma exceção, pois contém compostos secundários não encontrados em outras Caliciaceae, incluindo cloroatranorina, rugulosina, zeorina e ácido crisofânico.[20]
Gêneros

.jpg)


Até junho de 2024, o Index Fungorum reconhecia 39 gêneros e 669 espécies na família Caliciaceae.[22] A seguir, apresenta-se uma lista dos gêneros da família Caliciaceae, baseada em uma revisão e resumo de classificação fúngica de 2020 por Wijayawardene e colegas.[17] Após o nome do gênero, consta a autoridade taxonômica (aqueles que primeiro circunscreveram o gênero; abreviações de autores padronizadas são usadas), o ano de publicação e o número de espécies:

.jpg)

.jpg)
- Acolium (Ach.) Gray (1821)[23] – 5 spp.
- Acroscyphus Lév. (1846)[24] – 1 sp.
- Allocalicium M.Prieto & Wedin (2016)[18] – 1 sp.
- Amandinea M.Choisy ex Scheid. & M.Mayrhofer (1993)[25] – 35 spp.
- Australiaena Matzer, H.Mayrhofer & Elix (1997)[26] – 1 sp.
- Baculifera Marbach & Kalb (2000) – 14 spp.
- Buellia De Not. (1846)[27] – 300 spp.
- Burrowsia Fryday & I.Medeiros (2020)[28] – 1 sp.
- Calicium Pers. (1794)[3] – ca. 30 spp.
- Chrismofulvea Marbach (2000) – 4 spp.
- Ciposia Marbach (2000)[29] – 1 sp.
- Cratiria Marbach (2000) – ca. 20 spp.
- Culbersonia Esslinger (2000)[32] – 1 sp.
- Dermatiscum Nyl. (1867)[33] – 3 spp.
- Dermiscellum Hafellner, H.Mayrhofer & Poelt (1979)[34] – 1 sp.
- Dimelaena Norman (1852)[35] – 10 spp.
- Diploicia A.Massal. (1852)[36] – ca. 12 spp.
- Diplotomma Flot. (1849) [37] – ca. 30 spp.
- Dirinaria (Tuck.) Clem. (1909)[38] – ca. 35 spp.
- Endohyalina Marbach (2000) – 10 spp.
- Fluctua Marbach (2000) – 1 sp.
- Gassicurtia Fée (1825)[39] – 30 spp.
- Hypoflavia Marbach (2000)[29] – 3 spp.
- Monerolechia Trevis. (1857)[40] – 4 spp.
- Orcularia (Malme) Kalb & Giralt (2011)[41] – 4 spp.
- Pseudothelomma M.Prieto & Wedin (2016)[18] – 2 spp.
- Pyxine Fr. (1825)[42] – ca. 75 spp.
- Redonia C.W.Dodge (1973)[43] – 2 spp.
- Santessonia Hale &; Vobis (1978)[44] – 10 spp.
- Sculptolumina Marbach (2000)[29] – 4 spp.
- Sphinctrinopsis Woron. (1927)[45] – 1 sp.
- Stigmatochroma Marbach (2000) – 9 spp.
- Tetramelas Norman (1852)[35] – 16 spp.
- Texosporium Nádv. ex Tibell & Hofsten (1968)[46] – 1 sp.
- Texosporium Nádv. ex Tibell & Hofsten (1968) – 1 sp.
- Thelomma A.Massal. (1860)[47] – 5 spp.
- Tholurna Norman (1861)[48] – 1 sp.
- Tylophoropsis Sambo (1938)[49] – 1 sp.[nota 1]
Alguns gêneros que até recentemente eram classificados como membros da família Caliciaceae tiveram seu status reavaliado à luz de estudos filogenéticos moleculares. Por exemplo, Cyphelium Ach. foi sinonimizado com Calicium em 2016, quando se descobriu que sua espécie-tipo se agrupava com este último gênero.[18] Hafellia foi colocada em sinonímia com Buellia quando uma proposta para substituir o tipo conservado de Buellia, B. disciformis, por B. aethalea,[50] foi recusada pelo Comitê de Nomenclatura de Fungos.[51] Essa proposta tentou manter B. disciformis como tipo para Hafellia, para que o nome pudesse continuar a ser usado para um grupo distinto de líquens crustosos com paredes de ascosporos espessadas, que historicamente foram tratados em Buellia.[50]
Habitat e distribuição
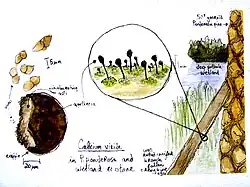
Coletivamente, as espécies de Caliciaceae têm uma distribuição cosmopolita, embora sejam especialmente predominantes em áreas temperadas e tropicais montanhosas. A maioria das espécies forma líquens que crescem em cascas ou madeira morta,[19] embora algumas crescem em rochas.[1] Em contraste, Texosporium faz parte das comunidades de crosta biológica do solo encontradas em áreas campestres, crescendo em esterco de vaca, solo e detritos.[18] Uma espécie rara conhecida apenas de uma única localidade na Suécia, Calicium episcalare, é parasita do líquen comum Hypocenomyce scalaris.[52]
Como a maioria das espécies calicioides, os líquens de Caliciaceae tendem a evitar competição com outros líquens e frequentemente crescem em micro-habitats negligenciados por outros líquens. Exemplos incluem: o lado de uma árvore não habitado por outras espécies cortícolas, em fendas profundas na casca ou buracos de pica-pau. Devido a essa preferência por locais negligenciados e seu tamanho geralmente pequeno, os líquens calicioides tendem a ser ignorados por coletores, e o grupo, em geral, é pouco conhecido.[53]
A Caliciaceae é uma das famílias cujas espécies são mais frequentemente encontradas em florestas de manguezais de Calabarzon (Filipinas).[54] Resultados semelhantes foram relatados em estudos sobre florestas de manguezais na Índia[55] e no Golfo da Tailândia,[56] onde os gêneros Dirinaria e Pyxine estavam entre os líquens folhosos mais comuns. Os líquens encontrados nos ecossistemas de manguezais tendem a ser tolerantes ao sal e sensíveis à umidade.[54] Usando técnicas de sequenciamento de última geração para detectar a assinatura genética dos organismos, as espécies de Caliciaceae foram identificadas como alguns dos líquens endolíticos mais comuns (líquens crustosos que crescem dentro de rochas sólidas) encontrados nos Vales Secos de McMurdo, na Antártica. Essa região apresenta um dos ambientes mais severos da Terra, com grandes variações de temperatura, aridez extrema, nutrientes limitados e altos níveis de radiação solar e UV.[57] O parceiro fotobionte do Caliciaceae é geralmente do gênero de algas verdes Trebouxia,[19] embora Stichococcus também tenha sido registrado com Calicium.[58]
Conservação

As espécies de Caliciaceae avaliadas para a Lista Vermelha global da IUCN incluem: Buellia asterella (criticamente em perigo, 2015),[59] Buellia gypsyensis (vulnerável, 2020),[60] Buellia sharpiana (vulnerável, 2020),[61] Calicium sequoiae (em perigo, 2021),[62] Texosporium sancti-jacobi (em perigo, 2020),[63] e Thelomma carolinianum (em perigo, 2017).[64] Na lista vermelha de macrofungos da China, Acroscyphus sphaerophoroides (em perigo, 2020) é o único representante dos Caliciaceae.[65]
Usos humanos
Embora não haja espécies de Caliciaceae conhecidas por terem significância econômica,[19] algumas foram exploradas para fins científicos. Em um estudo sueco de 1992, o líquen arbustivo (Allocalicium adaequatum, então um membro de Calicium) foi proposto como uma das várias espécies bioindicadoras que poderiam ser usadas para avaliar a continuidade florestal em florestas coníferas boreal.[66] Um estudo semelhante conduzido na ecorregião florestal acadiana do Canadá Atlântico encontrou várias espécies de Caliciaceae adequadas para determinar a continuidade em florestas primárias.[67]
Estudos realizados na Índia sugerem que Pyxine cocoes, um líquen folhoso comum e tolerante à poluição, é um candidato para biomonitoramento da poluição atmosférica local.[68] Ele bioacumula metais pesados tóxicos adquiridos do ar e retém os poluentes no talo, que podem ser amostrados e analisados para determinar sua concentração.[69] Outras espécies de Pyxine na Índia com propriedades semelhantes de resistência à poluição são P. hispidula e P. subcinerea.[70][71]
Buellia frigida, uma espécie crustosa que cresce em rochas no rigoroso clima antártico, tem sido frequentemente usada como um organismo modelo em pesquisas de astrobiologia. Este líquen extremófilo foi exposto a condições que simulam aquelas encontradas no espaço e em corpos celestes como Marte, incluindo vácuo, radiação UV e secura extrema. A espécie demonstrou resiliência a esses estressores relacionados ao espaço, tornando-a uma candidata para estudar como a vida pode se adaptar e potencialmente sobreviver em ambientes extraterrestres.[72]
Registro fóssil
Um fóssil pertencente a Calicium foi descrito a partir de âmbar báltico datado de 55–35 milhões de anos atrás (Ma).[73] Este e outros fósseis foram usados para estimar a data da separação entre os Caliciales (ou seja, o clado Caliciaceae-Physciaceae, conforme proposto por Gaya e colegas em 2012[16]) e os Teloschistales. Em sua análise, isso ocorreu no Jurássico Médio, cerca de 171 Ma.[18] Outros fósseis foram encontrados desde então. Em 2018, três fósseis atribuídos a Calicium foram registrados em âmbar europeu do Paleógeno.[74] Um fóssil de fungo em âmbar báltico, originalmente coletado por Robert Caspary e atribuído ao gênero Stilbum (família Chionosphaeraceae) em 1886, foi reavaliado como membro dos Caliciales em 2019, como Calicium succini. Por ser considerado "bem preservado e identificável de forma confiável", é valioso para estudos evolutivos que estimam os tempos de divergência de linhagens fúngicas.[75]
Notas
- ↑ a b De acordo com o Index Fungorum,[30][31] os gêneros Ciposia e Tylophoropsis não são nomes corretos, pois suas publicações foram precedidas por homônimos, os gêneros de plantas Ciposia (Myrtaceae) e Tylophoropsis (Apocynaceae). Os nomes publicados mais cedo têm precedência de acordo com o princípio da prioridade.
Ligações externas
Referências
- ↑ a b c d e f g h i Jaklitsch, Walter; Baral, Hans-Otto; Lücking, Robert; Lumbsch, H. Thorsten (2016). Frey, Wolfgang, ed. Syllabus of Plant Families: Adolf Engler's Syllabus der Pflanzenfamilien. 1/2 13 ed. Berlin Stuttgart: Gebr. Borntraeger Verlagsbuchhandlung, Borntraeger Science Publishers. pp. 118–119. ISBN 978-3-443-01089-8. OCLC 429208213
- ↑ Chevallier, François-Fulgis (1826). Flore Générale des Environs de Paris (em francês). 1. Paris: Chez Ferra Jeune, librairie, rue des Grands-Augustins. p. 314
- ↑ a b Persoon, C.H. (1794). «Einige Bemerkungen über die Flechten: Nebst Beschreibungen einiger neue Arten aus dieser». Annalen der Botanik (em alemão). 7: 1–32
- ↑ a b Prieto, Maria; Baloch, Elisabeth; Tehler, Anders; Wedin, Mats (2013). «Mazaedium evolution in the Ascomycota (Fungi) and the classification of mazaediate groups of formerly unclear relationship». Cladistics. 29 (3): 296–308. PMID 34818827. doi:10.1111/j.1096-0031.2012.00429.x

- ↑ a b c Wedin, Mats; Tibell, Leif (1997). «Phytogeny and evolution of Caliciaceae, Mycocaliciaceae, and Sphinctrinaceae (Ascomycota), with notes on the evolution of the prototunicate ascus». Canadian Journal of Botany. 75 (8): 1236–1242. doi:10.1139/b97-837
- ↑ Kimbrough, James W. (1970). «Current Trends in the Classification of Discomycetes». Botanical Review. 36 (2): 96–161 (see pp. 125–126. Bibcode:1970BotRv..36...91K. JSTOR 4353790. doi:10.1007/BF02858958
- ↑ Wedin, Mats. «Acharius Medallists: Leif Tibell». International Association for Lichenology. Consultado em 20 de janeiro de 2021. Arquivado do original em 2016
- ↑ a b Tibell, Leif (1984). A Reappraisal of the Taxonomy of Caliciales. Col: Beihefte zur Nova Hedwigia. 79. [S.l.]: J. Cramer. pp. 597–713
- ↑ Tibell, Leif (1996). Caliciales. Col: Flora Neotropica. 69. [S.l.]: New York Botanical Garden Press. pp. 1–78. JSTOR 4393866
- ↑ Gargas, A.; DePriest, P.; Grube, M.; Tehler, A. (1995). «Multiple origins of lichen symbioses in fungi suggested by SSU rDNA phylogeny». Science. 268 (5216): 1492–1495. Bibcode:1995Sci...268.1492G. PMID 7770775. doi:10.1126/science.7770775
- ↑ a b Aptroot, André; Maphangwa, Khumbudzo Walter; Zedda, Luciana; Tekere, Memory; Alvarado, Pablo; Sipman, Harrie J. M. (2019). «The phylogenetic position of Culbersonia is in the Caliciaceae (lichenized ascomycetes)». The Lichenologist. 51 (2): 187–191. doi:10.1017/S0024282919000033
- ↑ Wedin, Mats; Döring, Heidi; Nordin, Anders; Tibell, Leif (2000). «Small subunit rDNA phylogeny shows the lichen families Caliciaceae and Physciaceae (Lecanorales, Ascomycotina) to form a monophyletic group». Canadian Journal of Botany. 78 (2): 246–254. doi:10.1139/b99-185
- ↑ Wedin, Mats; Baloch, Elisabeth; Grube, Martin (2002). «Parsimony analyses of mtSSU and nITS rDNA sequences reveal the natural relationships of the lichen families Physciaceae and Caliciaceae». Taxon. 51 (4): 655–660. JSTOR 1555020. doi:10.2307/1555020
- ↑ Miadlikowska, J.; Kauff, F.; Hofstetter, V.; Fraker, E.; Grube, M.; Hafellner, J.; et al. (2007). «New insights into classification and evolution of the Lecanoromycetes (Pezizomycotina, Ascomycota) from phylogenetic analyses of three ribosomal RNA- and two protein-coding genes». Mycologia. 98 (6): 1088–1103. PMID 17486983. doi:10.1080/15572536.2006.11832636
- ↑ Wedin, Mats; Grube, Martin (2002). «(1555) Proposal to conserve Physciaceae nom. cons. against an additional name Caliciaceae (Lecanorales, Ascomycota)». Taxon. 51 (4): 802. JSTOR 1555043. doi:10.2307/1555043
- ↑ a b Gaya, Ester; Högnabba, Filip; Holguin, Ángela; Molnar, Katalin; Fernández-Brime, Samantha; Stenroos, Soili; Arup, Ulf; Søchting, Ulrik; Boom, Pieter Van den; Lücking, Robert; Sipman, Harrie J.M.; Lutzoni, François (2012). «Implementing a cumulative supermatrix approach for a comprehensive phylogenetic study of the Teloschistales (Pezizomycotina, Ascomycota)». Molecular Phylogenetics and Evolution. 63 (2): 374–387. PMID 22306043. doi:10.1016/j.ympev.2012.01.012
- ↑ a b Wijayawardene, Nalin; Hyde, Kevin; Al-Ani, LKT; Dolatabadi, S; Stadler, Marc; Haelewaters, Danny; et al. (2020). «Outline of Fungi and fungus-like taxa». Mycosphere. 11: 1060–1456. doi:10.5943/mycosphere/11/1/8. hdl:10481/61998
- ↑ a b c d e f Prieto, Maria; Wedin, Mats (2016). «Phylogeny, taxonomy and diversification events in the Caliciaceae». Fungal Diversity. 82 (1): 221–238. doi:10.1007/s13225-016-0372-y
- ↑ a b c d e f Cannon, Paul F.; Kirk, Paul M. (2007). Fungal Families of the World. [S.l.]: CAB International. pp. 48–49. ISBN 978-0851998275
- ↑ a b Joneson, Suzanne; Glew, Katherine A. (2003). «Acroscyphus (Caliciaceae) in North America». The Bryologist. 106 (3): 443–446. doi:10.1639/10
- ↑ McCune, Bruce; Rosentreter, Roger (1992). «Texosporium sancti-jacobi, a rare western North American lichen». The Bryologist. 95 (3): 329–333. JSTOR 3243492. doi:10.2307/3243492
- ↑ Species Fungorum. «Caliciaceae». Catalog of Life. Consultado em 23 de junho de 2024. Arquivado do original em 5 de junho de 2023
- ↑ Gray, Samuel F. (1821). A Natural Arrangement of British Plants. 1. London: Baldwin, Cradock, and Joy. p. 482
- ↑ Léveillé, J.H. (1846). «Descriptions des champignons de l'herbier du Muséum de Paris». Annales des Sciences Naturelles Botanique. 3 (em francês). 5: 262
- ↑ Scheidegger, Christoph (2007). «A revision of European saxicolous species of the genus Buellia De Not. and formerly included genera». The Lichenologist. 25 (4): 315–364. doi:10.1006/lich.1993.1001
- ↑ Matzer, M.; Mayrhofer, H.; Elix, J.A. (2007). «Australiaena streimannii, a new genus and species in the Physciaceae from tropical Australasia». The Lichenologist. 29 (1): 39–79. doi:10.1017/S0024282997000078
- ↑ De Notaris, G. (1846). «Framennti lichenographici di un lavoro inedito». Parlatore, Giornale Botanico Italiano (em italiano). 2: 174–224
- ↑ Fryday, Alan M.; Medeiros, Ian D.; Siebert, Stefan J.; Pope, Nathaniel; Rajakaruna, Nishanta (2020). «Burrowsia, a new genus of lichenized fungi (Caliciaceae), plus the new species B. cataractae and Scoliciosporum fabisporum, from Mpumalanga, South Africa». South African Journal of Botany. 132: 471–481. doi:10.1016/j.sajb.2020.06.001
- ↑ a b c Marbach 2000.
- ↑ Kirk, Paul (ed.). «Record Details: Ciposia Marbach». Index Fungorum. Consultado em 15 de janeiro de 2021
- ↑ Kirk, Paul (ed.). «Record Details: Tylophoropsis Sambo». Index Fungorum. Consultado em 15 de janeiro de 2021
- ↑ Esslinger, Theodore L. (2000). «Culbersonia americana, a rare new lichen (Ascomycota) from Western America». The Bryologist. 103 (4): 771–773. doi:10.1639/0007-2745(2000)103[0771:CAARNL]2.0.CO;2
- ↑ Nylander, W. (1867). «Circa genus Lichenum Dermatiscum». Botanische Zeitung (em latim). 25. 133 páginas
- ↑ Hafellner, J.; Mayrhofer, H.; Poelt, J. (1979). «Die Gattungen der Flechtenfamilie Physciaceae». Herzogia (em alemão). 5 (1–2): 39–79. doi:10.1127/herzogia/5/1979/39
- ↑ a b Norman, J.M. (1852). «Conatus praemissus redactionis novae generum nonnullorum Lichenum in organis fructificationes vel sporis fundatae». Nytt Magazin for Naturvidenskapene (em latim). 7: 213–252 (see pp. 231, 236)
- ↑ Massalongo, Abramo Bartolomeo (1852). Ricerche sull'autonomia dei licheni crostosi e materiali pella loro naturale ordinazione (em italiano). Verona: Dalla tipografia di A. Frizierio. p. 86. OCLC 23511705
- ↑ Flotow, J.V. (1849). «Lichenes florae silesiae». Jahresbericht der Schlesischen Gesellschaft für Vaterländische Kultur (em alemão). 27: 130
- ↑ Clements, Frederic E. (1909). The Genera of Fungi. Minneapolis: The H. W. Wilson Co. p. 84
- ↑ Fée, Antoine Laurent Apollinaire (1825). Essai sur les cryptogames des écorces exotiques officinales (em francês). Paris: Firmin Didot père et fils. p. 100
- ↑ Trevisan, Victore (1856). «Brigantiaea, novum Lichenum genus». Linnaea (em latim). 28: 283–298
- ↑ Kalb, K.; Giralt, M. (2011). «Orcularia, a segregate from the lichen genera Buellia and Rinodina (Lecanoromycetes, Caliciaceae)». Phytotaxa. 38: 53–60. doi:10.11646/phytotaxa.38.1.8
- ↑ Fries, Elias M. (1825). Systema Orbis Vegetabilis (em latim). 1. Lundin: Typographia Academica. p. 267
- ↑ Dodge, C.W. (1973). Lichen flora of the Antarctic continent and adjacent islands. Canaan, New Hampshire: Phoenix Publishing. ISBN 978-0914016014
- ↑ Hale, Mason E.; Vobis, Gernot (1978). «Santessonia, a new lichen genus from Southwest Africa». Botaniska Notiser. 131 (1): 1–5
- ↑ Woronichin, N.N. (1927). «Contribution à la flore mycologique du Caucase». Travaux du Musée Botanique de l'Académie des Sciences de Russie (em francês). 21: 87–243 (see p. 103)
- ↑ Tibell, Leif; Hofsten, Angelica van (1968). «Spore evolution of the lichen Texosporium sancti-jacobi (=Cyphelium sancti-jacobi)». Mycologia. 60 (3): 553–558. JSTOR 3757422. doi:10.1080/00275514.1968.12018604
- ↑ Massalongo, Abramo Bartolomeo (1860). «Esame comparativo di alcune genere di licheni». Atti dell'Imperial Regio Istituto Veneto di Scienze (em italiano). 5 (3): 247–276
- ↑ Norman, J.M. (1861). «Descriptio speciei novae lichenis, quam detexit et sub nomine Tholurna dissimilis proponit». Flora (Regensburg) (em latim). 44: 409
- ↑ Cengia Sambo, Maria (1938). «Licheni del Kenia e del Tanganica raccotti dai rev. Padri della Consolata». Nuovo Giornale Botanico Italiano (em italiano). 45 (3): 364–387. doi:10.1080/11263503809438758
- ↑ a b Moberg, R.; Nordin, A.; Scheidegger, C. (1999). «(1384) Proposal to change the listed type of the name Buellia, nom. cons. (Physciaceae, lichenised Ascomycota)». Taxon. 48 (1): 14. JSTOR 1224634. doi:10.2307/1224634
- ↑ Gams, Walter (2004). «Report of the Committee for Fungi: 11». Taxon. 53 (4): 1067–1069. JSTOR 4135578. doi:10.2307/4135578
- ↑ Tibell, Leif; Knutsson, Tommy (2016). «Calicium episcalaris, (Caliciaceae), a new lichen species from Sweden» (PDF). Symbolae Botanicae Upsaliensis. 38: 49–52
- ↑ Selva, Steven B. (2013). «The calicioid lichens and fungi of the Acadian Forest Ecoregion of northeastern North America, I. New species and range extensions». The Bryologist. 116 (3): 248–256. doi:10.1639/0007-2745-116.3.248
- ↑ a b Lucban, M.C.; Paguirigan, J.A.G. (2019). «Occurrence of manglicolous lichens in Calabarzon, Philippines». Studies in Fungi. 4 (1): 263–273. doi:10.5943/sif/4/1/28
- ↑ Sethy, P. (2012). «Lichens on mangrove plants in Andaman Islands, India». Mycosphere. 3 (4): 476–484. doi:10.5943/mycosphere/3/4/11
- ↑ Rangsiruji, Achariya; Meesim, Sanya; Buaruang, Kawinnat; Boonpragob, Kansri; Mongkolsuk, Pachara; Binchai, Sutheewan; Pringsulaka, Onanong; Parnmen, Sittiporn (2020). «Molecular systematics and species distribution of foliose lichens in the Gulf of Thailand mangroves with emphasis on Dirinaria picta species complex» (PDF). Songklanakarin Journal of Science and Technology. 42 (3): 504–514. doi:10.14456/sjst-psu.2020.65. Consultado em 15 de janeiro de 2021. Arquivado do original (PDF) em 15 de julho 2020
- ↑ Coleine, Claudia; Pombubpa, Nuttapon; Zucconi, Laura; Onofri, Silvano; Stajich, Jason E.; Selbmann, Laura (2020). «Endolithic fungal species markers for harshest conditions in the McMurdo Dry Valleys, Antarctica». Life. 10 (2). 13 páginas. Bibcode:2020Life...10...13C. PMC 7175349. PMID 32041249. doi:10.3390/life10020013
- ↑ Rambold, Gerhard; Friedl, Thomas; Beck, Andreas (1998). «Photobionts in lichens: possible indicators of phylogenetic relationships?». The Bryologist. 101 (3): 392–397. JSTOR 3244177. doi:10.2307/3244177
- ↑ Spribille, T.; Bilovitz, P.; Printzen, C.; Haugan, R.; Timdal, E. (2015). «Buellia asterella». Lista Vermelha de Espécies Ameaçadas. 2015: e.T70385861A70385867. doi:10.2305/IUCN.UK.2015-4.RLTS.T70385861A70385867.en
- ↑ Fryday, A. (2020). «Buellia gypsyensis». Lista Vermelha de Espécies Ameaçadas. 2020: e.T176075451A177303493. doi:10.2305/IUCN.UK.2020-3.RLTS.T176075451A177303493.en
- ↑ Allen, J.; Lendemer, J.; McMullin, T. (2020). «Buellia sharpiana». Lista Vermelha de Espécies Ameaçadas. 2020: e.T80702844A80702847. doi:10.2305/IUCN.UK.2020-3.RLTS.T80702844A80702847.en
- ↑ Reese Næsborg, R. (2021). «Calicium sequoiae». Lista Vermelha de Espécies Ameaçadas. 2021: e.T180412795A184974492. doi:10.2305/IUCN.UK.2021-2.RLTS.T180412795A184974492.en
- ↑ Stone, D.; Root, H.; Hollinger, J.; Rosentreter, R.; Chandler, A.; Allen, J. (2020). «Texosporium sancti-jacobi». Lista Vermelha de Espécies Ameaçadas. 2020: e.T175708768A175710642. doi:10.2305/IUCN.UK.2020-3.RLTS.T175708768A175710642.en
- ↑ Lendemer, J. (2020). «Thelomma carolinianum». Lista Vermelha de Espécies Ameaçadas. 2020: e.T80703121A80703126. doi:10.2305/IUCN.UK.2020-3.RLTS.T80703121A80703126.en
- ↑ Yijian, Yao; Jiangchun, Wei; Wenying, Zhuang; Tiezheng, Wei; Yi, Li; Xinli, Wei; et al. (2020). «Threatened species list of China's macrofungi». Biodiversity Science. 28 (1): 20–25. doi:10.17520/biods.2019174
- ↑ Tibell, Leif (1992). «Crustose lichens as indicators of forest continuity in boreal coniferous forests». Nordic Journal of Botany. 12 (4): 427–450. doi:10.1111/j.1756-1051.1992.tb01325.x
- ↑ Selva, Steven B. (2003). «Using calicioid lichens and fungi to assess ecological continuity in the Acadian Forest Ecoregion of the Canadian Maritimes». The Forestry Chronicle. 79 (3): 550–558. doi:10.5558/tfc79550-3
- ↑ Bajpai, Rajesh; Upreti, Dalip K.; Nayaka, S.; Kumari, B. (2010). «Biodiversity, bioaccumulation and physiological changes in lichens growing in the vicinity of coal-based thermal power plant of Raebareli district, north India». Journal of Hazardous Materials. 174 (1–3): 429–436. PMID 19818555. doi:10.1016/j.jhazmat.2009.09.071
- ↑ Singh, Pramod Kumar; Bujarbarua, P.; Singh, K.P.; Tandon, P.K. (2019). «Report on the bioaccumulation of heavy metals by foliose lichen (Pyxine cocoes) from air polluted area near Nagaon Paper Mill in Marigaon, Assam, North-East India». Journal on New Biological Reports. 8 (1): 15–21. ISSN 2319-1104
- ↑ Shukla, Vertika; Upreti, Dalip K. (2007). «Effect of metallic pollutants on the physiology of lichen, Pyxine subcinerea Stirton in Garhwal Himalayas». Environmental Monitoring and Assessment. 141 (1–3): 237–243. PMID 17879139. doi:10.1007/s10661-007-9891-z
- ↑ Shukla, Vertika; Upreti, Dalip K. (2010). «Changing lichen diversity in and around urban settlements of Garhwal Himalayas due to increasing anthropogenic activities». Environmental Monitoring and Assessment. 174 (1–4): 439–444. PMID 20440642. doi:10.1007/s10661-010-1468-6
- ↑ Meeßen, J.; Sánchez, F. J.; Brandt, A.; Balzer, E.-M.; de la Torre, R.; Sancho, L. G.; de Vera, J.-P.; Ott, S. (2013). «Extremotolerance and resistance of lichens: comparative studies on five species used in astrobiological research I. Morphological and anatomical characteristics». Origins of Life and Evolution of Biospheres. 43 (3): 283–303. Bibcode:2013OLEB...43..283M. PMID 23868319. doi:10.1007/s11084-013-9337-2
- ↑ Rikkinen, Jouko (2003). «Calicioid lichens from European Tertiary amber». Mycologia. 95 (6): 1032–1036. PMID 21149012. doi:10.1080/15572536.2004.11833019
- ↑ Rikkinen, Jouko; Meinke, S. Kristin L.; Grabenhorst, Heinrich; Gröhn, Carsten; Kobbert, Max; Wunderlich, Jörg; Schmidt, Alexander R. (2018). «Calicioid lichens and fungi in amber – Tracing extant lineages back to the Paleogene». Geobios. 51 (5): 469–479. Bibcode:2018Geobi..51..469R. doi:10.1016/j.geobios.2018.08.009. hdl:10138/308761
- ↑ Kettunen, Elina; Sadowski, Eva-Maria; Seyfullah, Leyla J.; Dörfelt, Heinrich; Rikkinen, Jouko; Schmidt, Alexander R.; Lomax, Barry (2019). «Caspary's fungi from Baltic amber: historic specimens and new evidence». Papers in Palaeontology. 5 (3): 365–389. Bibcode:2019PPal....5..365K. doi:10.1002/spp2.1238. hdl:10138/309518