Bonacynodon
| Bonacynodon | |
|---|---|
| |
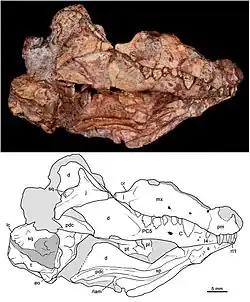
| Crânio do holótipo | |
| Classificação científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Clado: | Synapsida |
| Clado: | Therapsida |
| Clado: | Cynodontia |
| Família: | †Probainognathidae |
| Gênero: | †Bonacynodon Martinelli et al., 2016 |
| Espécies: | †B. schultzi
|
| Nome binomial | |
| †Bonacynodon schultzi Martinelli et al., 2016
| |
Bonacynodon é um gênero extinto de cinodontes que viveu no que hoje é o sul do Brasil durante o período Triássico (idades Ladiniano–Carniano).[1] O gênero é monotípico, contendo apenas a espécie-tipo Bonacynodon schultzi. B. schultzi é conhecido por dois espécimes, consistindo em dois crânios parciais e algumas partes mal preservadas do pós-crânio. Ambos os espécimes foram recuperados da formação Santa Maria, parte da supersequência Santa Maria da bacia do Paraná. Essa sequência preserva uma associação faunística conhecida como Zona de Assembleia Dinodontosaurus, que contém várias outras espécies de cinodontes, dicinodontes e répteis. Bonacynodon era um cinodonte pequeno, provavelmente insetívoro, com comprimento estimado em cerca de 30 cm. Pode ser distinguido de outros cinodontes por seus grandes caninos serrilhados (semelhantes a uma serra). Junto com o gênero Probainognathus [en] da Argentina, formava a família Probainognathidae [en], uma das linhagens mais basais do clado Probainognathia. Era um parente próximo dos mamíferos, o único grupo de cinodontes vivo hoje.
Descoberta e nomeação
.png)
Bonacynodon schultzi é conhecido por dois espécimes: o holótipo MCT-1716-R e o espécime referido (parátipo) MCT-1717-R. Ambos foram descobertos na década de 1940 pelo paleontólogo brasileiro Llewellyn Ivor Price em dois afloramentos rochosos distintos (sangas) na região de Pinheiros, cerca de 12 km ao sul da cidade de Candelária, Rio Grande do Sul.[1] As rochas pertencem à formação Santa Maria da supersequência Santa Maria, datadas do Ladiniano ao início do Carniano.[2]
O holótipo foi encontrado em um afloramento conhecido como Sanga do Janguta, preservado junto com um espécime do dicinodonte Dinodontosaurus. Consiste principalmente em um crânio parcial, incluindo partes do teto craniano, mandíbula inferior, dentição superior e inferior, base craniana e palato. O teto craniano foi severamente danificado durante a preparação. O holótipo também inclui elementos pós-cranianos muito fragmentados, que não foram descritos em detalhes. O espécime referido foi encontrado em outro afloramento, a Sanga do Forno. Assim como o holótipo, preserva partes do crânio, mandíbula inferior e dentição superior e inferior. Sua forma foi fortemente distorcida durante a fossilização, mas preserva algumas partes do crânio ausentes no holótipo. Com base na ausência de facetas de desgaste nos dentes, sugeriu-se que ambos os espécimes representam indivíduos subadultos.[1]
Os espécimes receberam uma descrição formal em 2016, quando Agustín G. Martinelli e colegas atribuíram um novo nome genérico e específico. O nome genérico Bonacynodon deriva do sobrenome de José Fernando Bonaparte, um paleontólogo argentino especializado em vertebrados mesozoicos da América do Sul. A segunda parte, cynodon, vem parcialmente da palavra em grego antigo κύων (kuōn, "cão"), referindo-se ao fato de ser um cinodonte. O epíteto específico schultzi homenageia Cesar L. Schultz, paleontólogo e professor brasileiro da Universidade Federal do Rio Grande do Sul.[1]
Descrição
Bonacynodon era um cinodonte pequeno, com comprimento total estimado em cerca de 30 cm.[3]
Crânio
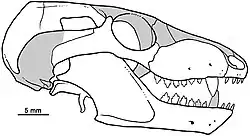
O crânio de Bonacynodon media cerca de 6–7 cm de comprimento. A região temporal (área atrás das órbitas) era larga e um pouco mais longa que o focinho. A crista sagital (uma crista estreita ao longo do topo do crânio) era relativamente baixa. O palato secundário (teto da boca) consistia em duas partes: uma seção maior formada pelos maxilares (ossos da mandíbula superior) e uma menor formada pelos ossos palatinos. Como no gênero próximo Probainognathus, o palato terminava antes do último pós-canino, sendo um pouco mais curto que o observado nos membros mais basais da família Chiniquodontidae e nos membros mais derivados do clado Prozostrodontia [en].[1]
A mandíbula superior era composta por dois ossos: o pré-maxilar na frente e o maxilar atrás. Acima dos caninos e pós-caninos frontais, havia múltiplos pequenos orifícios chamados forames infraorbitais; pelo menos três forames são visíveis no holótipo, enquanto o espécime referido preserva dois ou três. Com base em outros membros basais de Probainognathia, Bonacynodon provavelmente tinha forames adicionais não preservados nos fósseis.[1] A parte da mandíbula superior que contém os dentes apresentava uma curva sigmoide (em forma de S), descendo perto dos pós-caninos antes de curvar-se para cima novamente perto da órbita. A mandíbula inferior consistia principalmente de um único osso, o dentário. O corpo do dentário era relativamente alto, e a sínfise (conexão entre as duas metades do dentário) aparentemente não era fundida. A parte frontal do dentário apresentava pelo menos três forames mentuais em cada lado. A parte posterior possuía uma projeção longa e voltada para trás, conhecida como processo coronoide. Uma grande depressão chamada fossa massetérica se estendia desde próximo ao último pós-canino até a ponta do processo coronoide. Como em outros membros de Probainognathidae, os ossos pós-dentários (um conjunto de ossos na parte posterior da mandíbula inferior) eram altamente reduzidos em comparação com cinodontes mais basais, formando uma estrutura pequena em forma de haste.[1]
Dentição

Bonacynodon possuía três tipos de dentes: incisivos, caninos e pós-caninos. Parece ter tido quatro pares de incisivos na mandíbula superior, que eram esguios, com seção transversal redonda. Os três primeiros incisivos tinham tamanhos aproximadamente iguais, enquanto o quarto era ligeiramente menor. Os incisivos eram amplamente espaçados, com um grande espaço (diastema) entre o terceiro e o quarto, e um espaço semelhante entre o último incisivo e o canino. Os caninos superiores eram grandes e achatados lateralmente, com bordas posteriores fortemente serrilhadas, uma característica autapomórfica (traço derivado único) do táxon. Os incisivos inferiores não são conhecidos em nenhum dos espécimes. Dos caninos inferiores, apenas uma raiz parcial esquerda do holótipo é conhecida, mas sua forma e tamanho eram semelhantes aos dos caninos superiores.[1]
Atrás de cada canino superior, havia seis dentes pós-caninos, amplamente espaçados e sem contato entre si. As coroas eram comprimidas lateralmente e possuíam múltiplas cúspides retas e não serrilhadas, dispostas em linha. Os três primeiros pós-caninos superiores tinham três cúspides, com a central (cúspide A) sendo a maior, e as cúspides frontal e posterior (cúspides B e C, respectivamente) menores. O quarto e quinto pós-caninos superiores possuíam uma pequena quarta cúspide (cúspide D) atrás da cúspide C, e possivelmente uma quinta cúspide acessória.[1] O sexto pós-canino superior era aparentemente menos desenvolvido que o quarto e quinto, com pelo menos três cúspides. Os pós-caninos inferiores são conhecidos de forma incompleta, mas parecem semelhantes aos superiores. Os cíngulos eram pouco desenvolvidos e não apresentavam cúspides. Diferentemente dos membros do clado Prozostrodontia, não havia uma mudança abrupta na morfologia entre os pós-caninos frontais e posteriores, e as raízes não eram constritas.[1]
Classificação
Ao descrever Bonacynodon, Martinelli e colegas realizaram uma análise filogenética para determinar suas relações com outros cinodontes. O gênero foi recuperado como grupo-irmão de Probainognathus, um gênero semelhante da formação Chañares da Argentina. Junto com Probainognathus, Bonacynodon foi colocado na família Probainognathidae. Os membros da família Probainognathidae foram considerados próximos aos Prozostrodontia, um clado que inclui mamíferos, o único grupo de cinodontes vivos, além de outros grupos. Análises posteriores confirmaram a relação de irmandade entre Bonacynodon e Probainognathus.[4][5][6][7]


Abaixo está um cladograma conforme a análise de Martinelli et al. (2016):[1]
| Probainognathia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiologia
Membros basais de Probainognathidae, como Ecteniniidae e Chiniquodon, tinham dentes pós-caninos com cúspides fortemente recurvadas (e, no caso de Ecteniniidae, até serrilhadas), adequadas para uma dieta carnívora. As cúspides retas e não serrilhadas dos pós-caninos de membros de Probainognathidae como Bonacynodon eram mais semelhantes às dos prozostrodontes basais, sugerindo uma adaptação para insetivoria.[1] Como o holótipo foi preservado junto com restos de um dicinodonte, Schwanke & Kellner (2009) sugeriram que o animal poderia ser um necrófago oportunista.[1][8] No entanto, em 2016, Martinelli e colegas propuseram que ele poderia ter se alimentado de larvas de insetos e outros invertebrados que se alimentavam do dicinodonte em decomposição.[1]
Paleoambiente

Bonacynodon pertence à Zona de Assembleia Dinodontosaurus, a mais basal das quatro unidades bioestratigráficas da supersequência Santa Maria da bacia do Paraná. A Zona de Assembleia Dinodontosaurus corresponde à sequência Pinheiros-Chiniquá, uma das três sequências estratigráficas da supersequência. Junto com a sequência Santa Cruz (contendo a Zona de Assembleia Santacruzodon) e a parte inferior da sequência Candelária (contendo a Zona de Assembleia Hyperodapedon), ela compõe a tradicional formação Santa Maria.[1] As sequências Pinheiros-Chiniquá e Santa Cruz parecem ter sido depositadas durante um período seco, com a paisagem dominada por planícies de loesse.[9][2]
Os cinodontes representam uma grande parte da fauna da Zona de Assembleia Dinodontosaurus. Além de Bonacynodon, incluem os membros de Probainognathidae Aleodon [en], Candelariodon, Chiniquodon e Protheriodon, e os membros da família Traversodontidae Luangwa, Massetognathus, Protuberum, Scalenodon [en] e Traversodon. Outros vertebrados dessa Zona de Assembleia incluem os dicinodontes Dinodontosaurus e Stahleckeria, o pararéptil Candelaria, Brasinorhynchus [en] (Rhynchosauria), Barberenasuchus (Archosauriformes), várias espécies de Pseudosuchia (o grupo que contém os crocodilianos modernos e seus parentes extintos) e o afanossauro (primitivo do clado Avemetatarsalia) Spondylosoma.[2]
Referências
- ↑ a b c d e f g h i j k l m n o Martinelli, A. G.; Soares, M. B.; Schwanke, C. (2016). «Two New Cynodonts (Therapsida) from the Middle-Early Late Triassic of Brazil and Comments on South American Probainognathians». PLOS ONE (em inglês). 11 (10): e0162945. ISSN 1932-6203. PMC 5051967
 . PMID 27706191. doi:10.1371/journal.pone.0162945
. PMID 27706191. doi:10.1371/journal.pone.0162945
- ↑ a b c Schultz, C. L.; Martinelli, A. G.; Soares, M. B.; Pinheiro, F. L.; Kerber, L.; Horn, B. L. D.; Pretto, F. A.; Müller, R. T.; Melo, T. P. (2020). «Triassic faunal successions of the Paraná Basin, southern Brazil». Journal of South American Earth Sciences. 104. 102846 páginas. doi:10.1016/j.jsames.2020.102846
- ↑ Geggel, L. (2016). «Meet the Ancient Reptile that Gave Rise to Mammals». Scientific American (em inglês)
- ↑ Guignard, M. L.; Martinelli, A. G.; Soares, M. B. (2019). «The postcranial anatomy of Brasilodon quadrangularis and the acquisition of mammaliaform traits among non-mammaliaform cynodonts». PLOS ONE. 14 (5): e0216672. PMC 6510408. PMID 31075140. doi:10.1371/journal.pone.0216672
- ↑ Martinelli, A. G.; Eltink, E.; Da-Rosa, Á. A. S.; Langer, M. C. (2017). «A new cynodont from the Santa Maria formation, south Brazil, improves Late Triassic probainognathian diversity». Papers in Palaeontology. 3 (3): 401–423. doi:10.1002/spp2.1081
- ↑ Martinelli, A.; Soares, M. B.; de Oliveira, T. V.; Rodrigues, P. G.; Schultz, C. L. (2017). «The Triassic eucynodont Candelariodon barberenai revisited and the early diversity of stem prozostrodontians». Acta Palaeontologica Polonica. 62. doi:10.4202/app.00344.2017
- ↑ Pacheco, C. P.; Martinelli, A. G.; Pavanatto, A. E. B.; Soares, M. B.; Dias-da-Silva, S. (2018). «Prozostrodon brasiliensis, a probainognathian cynodont from the Late Triassic of Brazil: second record and improvements on its dental anatomy». Historical Biology. 30 (4): 475–485. doi:10.1080/08912963.2017.1292423. hdl:11336/94044
- ↑ Schwanke, C.; Kellner, A. W. (2009). «Interações ecológicas no Triássico». In: Stock Da-Rosa, Á. A. Vertebrados Fósseis de Santa Maria e região. [S.l.]: Santa Maria: Gráfica Editora Pallotti. pp. 279–301
- ↑ Horn, B. L. D.; Goldberg, K.; Schultz, C. L. (2018). «A loess deposit in the Late Triassic of southern Gondwana, and its significance to global paleoclimate». Journal of South American Earth Sciences. 81: 189–203. doi:10.1016/j.jsames.2017.11.017
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||





