Archaeamphora longicervia
Archaeamphora longicervia
| |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ocorrência: Cretáceo Inferior | |||||||||||||||||||
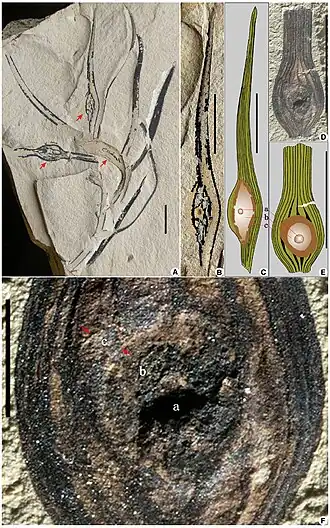 Fósseis de A. longicervia descritos por Wong et al. (2015) | |||||||||||||||||||
| Classificação científica | |||||||||||||||||||
| |||||||||||||||||||
| Nome binomial | |||||||||||||||||||
| Archaeamphora longicervia H.Q.Li (2005)[1] | |||||||||||||||||||
Archaeamphora longicervia é uma espécie de planta fóssil, o único membro do gênero hipotético Archaeamphora. O material fóssil atribuído a este táxon provém da Formação Yixian, no nordeste da China, datada do Cretáceo Inferior (cerca de 145 a 100 milhões de anos atrás).[1]
Originalmente descrita como uma planta carnívora com afinidades próximas às espécies existentes da família Sarraceniaceae, seria a mais antiga planta carnívora conhecida e o único registro fóssil dessa família, conhecida como a família das plantas carnívoras do Novo Mundo.[2][3][4][5] Archaeamphora também é um dos três gêneros mais antigos conhecidos de angiospermas (plantas com flores). Li (2005) escreveu que "a existência de uma angiosperma tão altamente derivada no Cretáceo Inferior sugere que as angiospermas devem ter surgido muito antes, talvez há 280 milhões de anos, como sugerem estudos de relógio molecular".[1]
Estudos posteriores questionaram a identificação de Archaeamphora como planta carnívora e até como angiosperma.[6][7][8] Os fósseis provavelmente representam folhas (acículas) da conífera Liaoningocladus boii deformadas por galhas induzidas por insetos.[9]
Etimologia
O nome genérico Archaeamphora deriva do grego αρχαίος, archaíos ("antigo"; forma combinada em latim: archae-), e ἀμφορεύς, amphoreus ("jarro"). O epíteto específico longicervia vem do latim longus ("longo") e cervicarius ("com pescoço"), em referência à constrição característica das estruturas semelhantes a jarros desta espécie.[1]
Material fóssil
Todo o material fóssil conhecido de A. longicervia provém da Formação Jianshangou em Beipiao [en], oeste de Liaoningue, China. Essas camadas do Cretáceo Inferior constituem a parte inferior da Formação Yixian, datada de 124,6 milhões de anos.[10][11][12] Nove espécimes de A. longicervia foram encontrados, incluindo o holótipo CBO0220 e o parátipo CBO0754.[1]
Descrição
Supunha-se que Archaeamphora longicervia fosse uma erva com cerca de 50 mm de altura. O caule, com pelo menos 21 mm de comprimento por 1,2 mm de largura, apresentava sulcos e cristas verticais distintas. As estruturas semelhantes a jarros eram ascidiformes e mediam de 30 a 40 mm de comprimento. Jarros maduros e folhas subdesenvolvidas, semelhantes a filódios, eram dispostos em espiral ao redor do caule. Os jarros consistiam em uma base tubular, seção intermediária expandida, constrição na boca e uma tampa vertical em forma de colher. Uma única asa percorria o lado adaxial de cada jarro. Três a cinco veias principais paralelas estavam presentes nos jarros, junto com algumas veias intercostais e numerosas vênulas.[1]
Duas estruturas incomuns em forma de bolsa estavam presentes em cada jarro, uma de cada lado da asa central. Estruturas semicirculares semelhantes foram encontradas na margem da tampa. Essas estruturas exibiam forte fluorescência intrínseca amarelo-esverdeada quando expostas à luz visível com um comprimento de onda de 500 nm (azul-esverdeado).[1]
Pequenas glândulas, com cerca de 4 μm de diâmetro, foram encontradas na superfície interna dos jarros, parcialmente embutidas nos sulcos ao longo das veias, exibindo forte fluorescência dourado-amarelada.[1]
Uma única semente foi encontrada intimamente associada ao material fóssil de A. longicervia e é presumida como pertencente à mesma espécie. É alada e apresenta reticulação tuberculada em sua morfologia, semelhante às sementes de táxons da família Sarraceniaceae. A semente é oval, coberta por verrugas marrom-escuras, e mede 0,9 por 1,25 mm.[1]
Taxonomia
O material fóssil de A. longicervia foi submetido a análise química para oleanano [en], considerado um marcador-chave que diferencia angiospermas de gimnospermas.[13] O oleanano foi detectado nesses espécimes, sugerindo que pertencem às angiospermas.[1]
Interpretação como planta carnívora

De acordo com Li (2005), várias características morfológicas de A. longicervia indicam uma relação próxima com Sarraceniaceae: ambos os táxons apresentam uma ou duas asas nos jarros, um peristoma liso e jarros que se estendem verticalmente a partir do topo de um pecíolo curto.[1]

Li (2005) sugere que A. longicervia é morfologicamente semelhante à moderna Sarracenia purpurea. Ela compartilha com esta espécie o arranjo espiral de seus jarros e folhas tubulares semelhantes a filódios com veias principais paralelas. Archaeamphora longicervia também se assemelha a espécies do gênero Heliamphora por ter jarros com um pescoço longo e tampa ereta. Notável é a semelhança entre as estruturas semicirculares espessas na tampa de A. longicervia e a grande "bolha" secretora de néctar presente na porção posterior superior dos jarros de Heliamphora exappendiculata [en].[nb 1][1]
Li (2005) menciona a descoberta de outro tipo de "planta carnívora" na mesma formação. Essa variedade difere do material-tipo de A. longicervia por ter jarros sem qualquer constrição antes da boca, expandindo-se gradualmente do pecíolo em uma forma oca semelhante a uma trombeta. Ele sugere que "deve ser uma espécie diferente" de A. longicervia. Uma forma intermediária com pescoço mais largo também é relatada, sugerindo que essas plantas já eram um grupo diversificado no Cretáceo Inferior.[1]
Entendimento atual
Heřmanová & Kvaček (2010) opinaram que a interpretação de Archaeamphora como planta carnívora é "problemática e o fóssil precisa de revisão".[7]
Em seu livro de 2011, Sarraceniaceae of South America [en], McPherson [en] et al. resumiram o pensamento atual sobre Archaeamphora da seguinte forma:[6]
Surgem sérias dúvidas que reduzem a probabilidade de que Archaeamphora longicervia pertença à linhagem das Sarraceniaceae ou seja mesmo uma planta carnívora. [...] Embora Archaeamphora possa ser uma representante das primeiras angiospermas na Terra [...] é muito improvável que represente um ancestral das Sarraceniaceae, pois é muito antiga para fazer parte do avançado "grupo coroa" de Ericales ao qual as Sarraceniaceae pertencem. [...] Outra contradição é que, exceto por Archaeamphora, não há outras evidências sugerindo que as Sarraceniaceae evoluíram fora do Novo Mundo, onde todos os membros existentes da família são endêmicos.
Wong et al. (2015) propuseram uma nova perspectiva:[9]
Archaeamphora longicervia H. Q. Li foi descrita como uma planta carnívora herbácea semelhante às Sarraceniaceae, da Formação Yixian do Cretáceo Inferior médio, na província de Liaoningue, nordeste da China. Aqui, uma reinvestigação dos espécimes de A. longicervia da Formação Yixian oferece novos insights sobre sua identidade e a morfologia das plantas carnívoras reivindicadas por Li. Demonstramos que os supostos jarros de Archaeamphora são galhas foliares induzidas por insetos que consistem em três componentes: (1) uma câmara larval interna; (2) uma zona intermediária de tecido nutritivo; e (3) uma parede externa de esclerênquima. Archaeamphora não é uma angiosperma carnívora semelhante às Sarraceniaceae, mas representa folhas com galhas da gimnosperma anteriormente relatada Liaoningocladus boii G. Sun et al. da Formação Yixian.
Habitat
A área habitada por A. longicervia provavelmente experimentou flutuações climáticas significativas durante o Cretáceo Inferior, variando de condições áridas ou semiáridas a mais úmidas.[15] O substrato da região era composto principalmente de sedimentos lacustres e rochas vulcânicas.[16]
Ver também
- Cephalotus follicularis, uma planta carnívora australiana cujas armadilhas em forma de jarro são convergentemente semelhantes às de Nepenthes
Notas
Referências
- ↑ a b c d e f g h i j k l m n o Li, H. 2005. «Early Cretaceous sarraceniacean-like pitcher plants from China.» (PDF). Consultado em 27 de janeiro de 2007. Cópia arquivada (PDF) em 2 de setembro de 2006 Acta Botanica Gallica 152(2): 227-234. (Informações Suplementares 1 Arquivado em 2016-02-22 no Wayback Machine, Informações Suplementares 2 Arquivado em 2016-06-30 no Wayback Machine)
- ↑ Krutzsch, W (1985). «Über Nepenthes-Pollen im europäischen Tertiär». Gleditschia. 13: 89–93
- ↑ Krutzsch, W (1989). «Paleogeography and historical phytogeography (paleochorology) in the Neophyticum». Plant Systematics and Evolution. 162 (1–4): 5–61. Bibcode:1989PSyEv.162....5K. doi:10.1007/BF00936909
- ↑ Anderson, J.A.R.; Muller, J. (1975). «Palynological study of a Holocene peat and a Miocene coal deposit from NW Borneo». Review of Palaeobotany and Palynology. 19 (4): 291–351. Bibcode:1975RPaPa..19..291A. doi:10.1016/0034-6667(75)90049-4
- ↑ Kumar, M (1995). «Pollen tetrads from Palaeocene sediments of Meghalaya, India: comments on their morphology, botanical affinity and geological records». Palaeobotanist. 43 (1): 68–81
- ↑ a b McPherson, S., A. Wistuba, A. Fleischmann & J. Nerz 2011. Sarraceniaceae of South America. Redfern Natural History Productions, Poole.
- ↑ a b Heřmanová, Z.; Kvaček, J. (2010). «Late Cretaceous Palaeoaldrovanda, not seeds of a carnivorous plant, but eggs of an insect». Journal of the National Museum (Prague), Natural History Series. 179 (9): 105–118. Consultado em 26 de outubro de 2012. Cópia arquivada em 9 de outubro de 2015
- ↑ Brittnacher, J (2013). «Phylogeny and biogeography of the Sarraceniaceae». Carnivorous Plant Newsletter. 42 (3): 99–106. doi:10.55360/cpn423.jb578

- ↑ a b Wong, WO; Dilcher, DL; Labandeira, CC; Sun, G; Fleischmann, A (2015). «Early Cretaceous Archaeamphora is not a carnivorous angiosperm». Frontiers in Plant Science. 6: 326. PMC 4423337. PMID 25999978. doi:10.3389/fpls.2015.00326
- ↑ Sun, G., S.-L. Zheng, D.L. Dilcher, Y.D. Wang & S.W Mei 2001. Early Angiosperms and their Associated Plants from Western Liaoning, China. Shanghai Scientific and Technological Education Publishing House, 227 pp.
- ↑ Sun, G.; Dilcher, D.L.; Zheng, S.-L.; Zhou, Z.-K. (1998). «In Search of the First Flower: A Jurassic Angiosperm, Archaefructus, from Northeast China» (PDF). Science. 282 (5394): 1692–1695. PMID 9831557. doi:10.1126/science.282.5394.1692. Cópia arquivada (PDF) em 19 de julho de 2011
- ↑ Swisher, III; Wang, Y-Q.; Wang, X.-L.; Xu, X.; Wang, Y. (1999). «Cretaceous age for the feathered dinosaurs of Liaoning, China» (PDF). Nature. 400 (6739): 58–61. Bibcode:1999Natur.400...58S. doi:10.1038/21872
- ↑ Moldowan, J.M.; Dahl, J.; Huizinga, B.J.; Fago, F.J.; Hickey, L.J.; Peakman, T.M.; Taylor, O.W. (1994). «The molecular fossil record of oleanane and its relationship to Angiosperms». Science. 265 (5173): 768–771. PMID 17736275. doi:10.1126/science.265.5173.768
- ↑ Nerz, J.; Wistuba, A. (2006). «Heliamphora exappendiculata, a clearly distinct species with unique characteristics». Carnivorous Plant Newsletter. 35 (2): 43–51. doi:10.55360/cpn352.jn909
- ↑ Barrett, P.M.; Hilton, J. M. (2006). «The Jehol Biota (Lower Cretaceous, China): new discoveries and future prospects». Integrative Zoology. 1 (1): 15–17. PMID 21395985. doi:10.1111/j.1749-4877.2006.00006.x
- ↑ Tan, J.-J.; Ren, D. (2006). «Ovatucupes: A New Cupedid Genus (Coleoptera: Archostemata: Cupedidae) From The Jehol Biota (Late Jurassic) Of Western Liaoning, China» (PDF). Entomological News. 117 (2): 223–232. doi:10.3157/0013-872x(2006)117[223:oancgc]2.0.co;2