Ciclo de vida
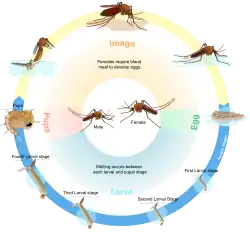
Em biologia, um ciclo vital ou ciclo de vida é a série de estádios pelos quais passa a vida de um organismo, que começam no zigoto, que costuma estar num ovo, passa por várias fases de desenvolvimento e conclui num adulto que se reproduz, produzindo descendência, que volta a repetir o ciclo.
"O conceito está estreitamente relacionado com o de história vital, desenvolvimento e ontogenia, mas difere deles em que dá mais ênfase à renovação."[1][2] As transições na forma podem implicar o crescimento, a reprodução assexual ou a reprodução sexual.
Nalguns organismos, sucedem-se diferentes "gerações" da espécie durante o seu ciclo de vida. No caso das plantas e muitas algas, há dois estádios multicelulares, e diz-se que têm um ciclo vital com alternância de gerações. O termo história vital usa-se às vezes para organismos como as algas vermelhas que têm três estádios multicelulares (ou mais), em vez de dois.[3] Nos humanos, não é o mesmo o conceito de geração que o conceito biológico de gerações, já que nos humanos é um conjunto de pessoas que, em média, nasceram por volta de um mesmo período de tempo.[4]
Os ciclos vitais que incluem reprodução sexual compreendem estádios alternantes haploides (com n cromossomas) e diploides (2n), é dizer, produz-se uma mudança de ploidia. Para voltar do estádio diploide ao haploide, deve ocorrer uma meiose. Quanto à mudança de ploidia, podem-se distinguir três tipos de ciclos:
- Ciclo haplonte — o estádio haploide é multicelular e o diploide unicelular e a meiose é zigótica. Por exemplo, nalgumas algas.
- Ciclo diplonte — o estádio diploide é multicelular e nele formam-se gâmetas haploides; a meiose é gamética. Por exemplo, em mamíferos.
- Ciclo haplodiplonte (também chamado diplohaplonte, diplobionte ou dibionte) — há estádios multicelulares diploides e haploides, e a meiose é espórica. Por exemplo, em musgos.
Os ciclos diferenciam-se no momento em que se produz a mitose (para o crescimento do organismo). As meioses zigótica e gamética vão seguidas de um estádio mitótico: a mitose ocorre durante a fase n em organismos com meiose zigótica e durante a fase 2n em organismos com meiose gamética. Portanto, a meiose zigótica e gamética denominam-se em conjunto "haplobiônticas" (uma só fase mitótica, que não se deve confundir com haplonte). Os organismos com meiose espórica apresentam dois estádios de mitose, nos estádios diploide e haploide, e denominam-se "diplobiônticos" (não confundir com diplontes).[carece de fontes]
Descoberta
O estudo da reprodução e desenvolvimento em organismos foi levado a cabo por muitos botânicos e zoólogos.
Wilhelm Hofmeister demonstrou que a alternância de gerações é uma característica comum em todas as plantas, e publicou este resultado em 1851.
Alguns termos (haplobionte e diplobionte) usados para a descrição dos ciclos vitais foram propostos inicialmente para as algas por Nils Svedelius, e acabaram por usar-se para outros organismos.[5][6] Outros termos (autogamia e gamontogamia) usados em ciclos vitais de protistas introduziu-os Karl Gottlieb Grell.[7]. A descrição dos complexos ciclos vitais de vários organismos contribuiu para descartar as ideias da geração espontânea nas décadas de 1840 e 1850.[8]
Ciclo vital haplonte
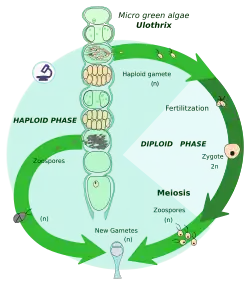
Dá-se em muitas algas, outros protistas e fungos. O indivíduo principal do ciclo é haploide e em todo o ciclo os zigotos são a única célula diploide; a mitose ocorre só na fase haploide.
Apresentam uma meiose zigótica, que é a que sofre um zigoto imediatamente depois da cariogamia, a qual é a fusão dos núcleos celulares das células reprodutoras. Deste modo, o organismo acaba a sua fase diploide e produz várias células haploides. Estas células dividem-se mitoticamente para formar indivíduos haploides multicelulares maiores ou mais células haploides. Têm dois tipos de gâmetas (masculino e feminino, geralmente) e estas células fundem-se para formar o zigoto.
Os indivíduos ou células resultantes da mitose são haploides, e o ciclo vital diz-se que é haplonte. Seres com ciclo haplonte dão-se:
- Em Archaeplastida: algumas algas verdes (por exemplo, Chlamydomonas, Zygnema, Chara)[9][10]
- Em Stramenopiles: algumas algas douradas[9][10]
- Em Alveolata: muitos dinoflagelados, por exemplo, Ceratium, Gymnodinium, alguns Apicomplexa (por exemplo, Plasmodium)[11]
- Em Rhizaria: alguns euglífidos,[12] ascetospóreos
- Em Excavata: alguns parabasálidos[13]
- Em Amoebozoa: Dictyostelium[9][10]
- Em Opisthokonta: a maioria dos fungos (alguns quitrídios, zigomicetos, alguns ascomicetos, basidiomicetos)[9][10][14]
Ciclo vital diplonte
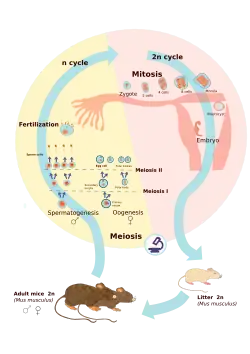
Apresentam-no os animais e alguns fungos, algas e outros protistas. O indivíduo principal do ciclo é diploide; em todo o ciclo, os gâmetas são as únicas células haploides e a mitose ocorre normalmente só na fase diploide.
Na meiose gamética, o zigoto diploide, em vez de dividir-se imediatamente por meiose para produzir células haploides, divide-se repetidamente por mitose para produzir um indivíduo multicelular diploide (ou um grupo de mais indivíduos unicelulares diploides nalguns organismos). As células dos indivíduos diploides sofrem depois a meiose para produzir células haploides, que são os gâmetas. As células haploides podem dividir-se outra vez por mitose para formar mais células haploides, como ocorre em muitas leveduras, mas a fase haploide não é a fase predominante do ciclo vital. Na maioria dos diplontes, a mitose ocorre só na fase diploide, é dizer, os gâmetas costumam fundir-se rapidamente para produzir os zigotos diploides.[15]
O indivíduo multicelular diploide é um diplonte com meiose gamética, pelo que é um ciclo diplonte. Encontramos organismos diplontes:
- Em Archaeplastida: algumas algas verdes (por exemplo, Cladophora glomerata,[16] Acetabularia[9][10])
- Em Stramenopiles: algumas algas castanhas (porém, nas Fucales, o ciclo vital pode também interpretar-se como fortemente heteromórfico-diplohaplonte, com uma fase gametofítica muito reduzida, como a das plantas com flor),[17] algumas xantófitas (por exemplo, Vaucheria),[18] a maioria das diatomáceas,[13] alguns oomicetos (por exemplo, Saprolegnia, Plasmopara viticola),[9][10] opalinas,[13] alguns "heliozoários" (por exemplo, Actinophrys, Actinosphaerium)[13][19]
- Em Alveolata: ciliados[13]
- Em Excavata: alguns parabasálidos[13]
- Em Opisthokonta: animais, alguns fungos (por exemplo, alguns ascomicetos)[9][10]
Ciclo vital haplodiplonte
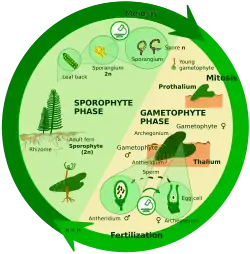
Dá-se em plantas, algumas algas, fungos e outros protistas. Há duas fases importantes, uma com indivíduos diploides e outra com haploides, o que se chama alternância de gerações. Segundo o grupo taxonómico pode ser mais predominante o diploide ou o haploide.
Na meiose espórica (também conhecida como meiose intermediária), o zigoto divide-se mitoticamente para produzir um diploide multicelular chamado esporófito. O esporófito cria esporos por meiose, os quais também se dividem depois mitoticamente produzindo indivíduos haploides chamados gametófitos. Os gametófitos produzem gâmetas por mitose. Nalgumas plantas o gametófito não só é de pequeno tamanho, mas também de curta vida; noutras plantas (musgos) e muitas algas, o gametófito é o estádio "dominante" do ciclo vital.[20]
Nas plantas observamos uma perda de importância progressiva do gametófito na escala evolutiva. O gametófito é o indivíduo principal nos musgos, mas está muito reduzido nos fetos, e nas plantas superiores fica reduzido só ao saco embrionário e ao pólen e não tem vida independente.
Encontramos haplodiplontes:
- Em Archaeplastida: algas vermelhas (que têm duas gerações esporofíticas), algumas algas verdes (por exemplo, Ulva), plantas terrestres[9][10]
- Em Stramenopiles: a maioria das algas castanhas[9][10]
- Em Rhizaria: muitos foraminíferos,[13] plasmodioforomicetos[9][10]
- Em Amoebozoa: mixogástridos
- Em Opisthokonta: alguns fungos (alguns quitrídios, alguns ascomicetos como Saccharomyces cerevisiae)[9][10]
- Outros eucariotas: haptófitas.[13]
Alguns animais têm um sistema de determinação do sexo chamado haplodiploide, mas isto não está relacionado com o ciclo de vida haplodiplonte.
Meiose vegetativa
Algumas algas vermelhas (como Bonnemaisonia[21] e Lemanea) e algas verdes (como Prasiola) têm meiose vegetativa, também chamada meiose somática, que é um processo raro.[22] A meiose vegetativa pode ocorrer em ciclos vitais haplodiplontes e diplontes. Os gametófitos permanecem unidos a uma parte do esporófito. As células diploides vegetativas (não reprodutoras) sofrem meiose, gerando células haploides vegetativas. Estas experimentam muitas mitoses e produzem gâmetas.
Um processo diferente, chamado diploidização vegetativa, um tipo de apomixia e produz-se nalgumas algas castanhas (por exemplo, Elachista stellaris).[23] As células duma parte haploide da planta duplicam os seus cromossomas espontaneamente para produzir tecidos diploides.
Ciclo de vida parasita
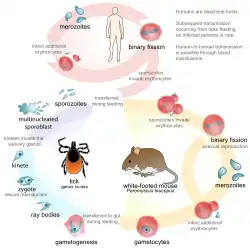
Os parasitas dependem da exploração de um ou mais hospedeiros. Aqueles que devem infetar mais de uma espécie hospedeira para completar o seu próprio ciclo vital diz-se que têm um ciclo vital complexo ou indireto. Por exemplo, Dirofilaria immitis, um verme nematodo parasita do cão, tem um ciclo vital indireto. As microfilárias devem ser primeiro ingeridas por um mosquito fêmea, onde se desenvolvem originando um estádio de larva infetivo. O mosquito depois pica um animal e transmite-lhe as larvas infeciosas, dentro do qual migram para a artéria pulmonar e amadurecem a adultos.[24]
Os parasitas que infetam uma só espécie têm ciclos vitais diretos. Um exemplo é Ancylostoma caninum, outro verme nematodo que afeta os cães. Desenvolve-se ao estádio larval infetivo no meio ambiente, depois penetra na pele do cão diretamente e amadurece a adulto no intestino delgado.[25]
Se um parasita tem que infetar um determinado hospedeiro para completar o seu ciclo vital, então diz-se que é um parasita obrigatório desse hospedeiro; às vezes, a infeção é facultativa (o parasita pode sobreviver e completar o seu ciclo de vida sem infetar essa determinada espécie hospedeira). Os parasitas às vezes infetam hospedeiros nos quais não podem completar os seus ciclos vitais; estes são hospedeiros acidentais.
Um hospedeiro no qual os parasitas se reproduzem sexualmente denomina-se hospedeiro definitivo, final ou primário. Em hospedeiros intermediários, os parasitas não se reproduzem ou fazem-no assexualmente, mas o parasita sempre se desenvolve a um novo estádio neste tipo de hospedeiro. Nalguns casos, o parasita infeta o hospedeiro, mas não experimenta nenhum desenvolvimento, estes hospedeiros conhecem-se como paraténicos[26] ou hospedeiros de transporte. O hospedeiro paraténico pode ser útil para aumentar a possibilidade de que o parasita se transmita ao hospedeiro definitivo. Por exemplo, o verme nematodo pulmonar do gato Aelurostrongylus abstrusus usa uma lesma ou caracol como hospedeiro intermediário; a larva do primeiro estádio entra no molusco e desenvolve-se ao terceiro estádio larval, que é infecioso para o hospedeiro definitivo, o gato. Se um rato come a lesma, a larva de terceiro estádio entra nos tecidos do rato, mas não sofre nenhum desenvolvimento.[carece de fontes]
Evolução
O tipo primitivo de ciclo vital provavelmente tinha indivíduos haploides com reprodução assexual.[13] Bactérias e arqueias apresentam um ciclo vital como este, e alguns eucariotas aparentemente também (por exemplo, Cryptophyta, Choanoflagellata, muitos Euglenozoa, muitos Amoebozoa, algumas algas vermelhas, algumas algas verdes, os fungos imperfeitos, alguns rotíferos e muitos outros grupos, não necessariamente haploides).[27] Porém, estes eucariotas provavelmente não são primitivamente assexuais, mas sim perderam a sua reprodução sexual, ou simplesmente não foi ainda observada.[28][29] Muitos eucariotas (incluindo animais e plantas) apresentam reprodução assexual, que pode ser facultativa ou obrigada no ciclo vital, e a reprodução sexual ocorre mais ou menos frequentemente.[30]
Os organismos que participam num ciclo vital biológico ordinariamente envelhecem e morrem, enquanto as células destes organismos que ligam gerações sucessivas do ciclo vital (as células da linha germinal e as suas descendentes) são potencialmente imortais. A base desta diferença é um problema fundamental em biologia. O biólogo e historiador russo Zhores A. Medvedev[31] considerou que a preservação da exatidão de um genoma replicativo e outros sistemas sintéticos por si sós não podem explicar a imortalidade das linhas germinais. Medvedev antes pensava que as características conhecidas da bioquímica e genética da reprodução sexual indicam a presença de informação especial de processos de manutenção e restauração no estádio de gametogénese do ciclo vital biológico. Em concreto, Medvedev considerou que as oportunidades mais importantes para a manutenção da informação das células germinais são as criadas por recombinação durante a meiose e a reparação do ADN; entendia estes como processos dentro das células da linha germinal capazes de restaurar a integridade do ADN e cromossomas perante os tipos de danos que causam o envelhecimento irreversível em células não germinais, por exemplo as células somáticas.[31]
O antepassado de todas as células atuais supõe-se que se pode rastrear, numa linhagem ininterrupta de 3000 milhões de anos até à origem da vida. Não é que as células sejam imortais mas sim que são linhagens multigeracionais.[32] A imortalidade duma linhagem celular depende da manutenção do potencial de divisão celular. Esse potencial pode perder-se em qualquer linhagem particular devido a danos celulares, à diferenciação terminal como ocorre em células nervosas, ou à morte celular programada (apoptose) durante o desenvolvimento. A manutenção da divisão celular potencial do ciclo vital biológico em sucessivas gerações depende de evitar os danos celulares e a reparação adequada de ditos danos, especialmente danos no ADN. Em organismos sexuais, a continuidade da linha germinal em gerações sucessivas do ciclo celular depende da efetividade do processo de evitar danos no ADN e reparar os que ocorram. Os processos sexuais em eucariotas proporcionam uma oportunidade para a reparação efetiva dos danos no ADN na linha germinal por recombinação homóloga.[32][33]
Referências
- ↑ Bell, Graham; Koufopanou, Vassiliki (1991). «The Architecture of the Life Cycle in Small Organisms». Philosophical Transactions: Biological Sciences. 332 (1262): 81–89. Bibcode:1991RSPTB.332...81B. JSTOR 55494. doi:10.1098/rstb.1991.0035
- ↑ Rodrigues, Juliany Cola Fernandes; Godinho, Joseane Lima Prado; De Souza, Wanderley (2014). «Biology of Human Pathogenic Trypanosomatids: Epidemiology, Lifecycle and Ultrastructure». Proteins and Proteomics of Leishmania and Trypanosoma. Col: Subcellular Biochemistry. 74. [S.l.: s.n.] pp. 1–42. ISBN 978-94-007-7304-2. PMID 24264239. doi:10.1007/978-94-007-7305-9_1
- ↑ Dixon, P.S. 1973. Biology of the Rhodophyta. Oliver & Boyd. ISBN 0 05 002485 X
- ↑ «Generational Insights and the Speed of Change». American Marketing Association (em inglês). 30 de junho de 2022. Consultado em 26 de julho de 2024. Cópia arquivada em 11 de abril de 2021
- ↑ C. Skottsberg (1961). Nils Eberhard Svedelius. 1873–1960. Biographical Memoirs of Fellows of the Royal Society. 7. [S.l.: s.n.] pp. 294–312. doi:10.1098/rsbm.1961.0023
- ↑ Svedelius, N. 1931. Nuclear Phases and Alternation in the Rhodophyceae. Arquivado em 2013-10-05 no Wayback Machine Em: Beihefte zum Botanischen Centralblatt. Band 48/1: 38–59.
- ↑ Margulis, L (6 de fevereiro de 1996). «Archaeal-eubacterial mergers in the origin of Eukarya: phylogenetic classification of life.». Proceedings of the National Academy of Sciences of the United States of America. 93 (3): 1071–1076. Bibcode:1996PNAS...93.1071M. PMC 40032
 . PMID 8577716. doi:10.1073/pnas.93.3.1071
. PMID 8577716. doi:10.1073/pnas.93.3.1071 - ↑ Moselio Schaechter (2009). Encyclopedia of Microbiology. Academic Press. Volume 4, p. 85.
- 1 2 3 4 5 6 7 8 9 10 11 Díaz, T.E.; Fernández-Carvajal, C.; Fernández, J.A. (2004). Curso de Botánica. Gijón: Trea
- 1 2 3 4 5 6 7 8 9 10 11 Díaz González, Tomás; Fernandez-Carvajal Alvarez, Mª del Carmen; Fernández Prieto, José Antonio. «Botânica: Ciclos biológicos de vegetales». Departamento de Biologia de Organismos e Sistemas, Universidade de Oviedo (em espanhol). Cópia arquivada em 14 de maio de 2020
- ↑ Sinden, R. E.; Hartley, R. H. (novembro de 1985). «Identification of the Meiotic Division of Malarial Parasites». The Journal of Protozoology. 32 (4): 742–744. PMID 3906103. doi:10.1111/j.1550-7408.1985.tb03113.x
- ↑ Lahr, Daniel J. G.; Parfrey, Laura Wegener; Mitchell, Edward A. D.; Katz, Laura A.; Lara, Enrique (22 de julho de 2011). «The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms». Proceedings of the Royal Society B: Biological Sciences. 278 (1715): 2081–2090. PMC 3107637. PMID 21429931. doi:10.1098/rspb.2011.0289
- 1 2 3 4 5 6 7 8 9 Ruppert, Edward E.; Fox, Richard S.; Barnes, Robert D. (2004). Invertebrate Zoology: A Functional Evolutionary Approach. [S.l.]: Thomson-Brooks/Cole. p. 26. ISBN 978-0-03-025982-1
- ↑ van den Hoek, Mann & Jahns 1995, p. 15.
- ↑ Smith, Gilbert M. (1938). «Nuclear Phases and Alternation of Generations in the Chlorophyceae». Botanical Review. 4 (3): 132–139. Bibcode:1938BotRv...4..132S. ISSN 0006-8101. JSTOR 4353174. doi:10.1007/BF02872350
- ↑ O. P. Sharma. Textbook of Algae, p. 189
- ↑ van den Hoek, Mann & Jahns 1995, p. 207.
- ↑ van den Hoek, Mann & Jahns 1995, p. 124.
- ↑ Bell, Graham (1988). Sex and Death in Protozoa: The History of Obsession. [S.l.]: Cambridge University Press. p. 11. ISBN 978-0-521-36141-5
- ↑ Bennici, Andrea (2008). «Origin and early evolution of land plants: Problems and considerations». Communicative & Integrative Biology (em inglês). 1 (2): 212–218. ISSN 1942-0889. PMC 2686025. PMID 19513262. doi:10.4161/cib.1.2.6987
- ↑ Salvador Soler, Noemi; Gómez Garreta, Amelia; Antonia Ribera Siguan, M. (agosto de 2009). «Somatic meiosis in the life history of Bonnemaisonia asparagoides and Bonnemaisonia clavata (Bonnemaisoniales, Rhodophyta) from the Iberian peninsula». European Journal of Phycology. 44 (3): 381–393. Bibcode:2009EJPhy..44..381S. doi:10.1080/09670260902780782
- ↑ van den Hoek, Mann & Jahns 1995, p. 82.
- ↑ Lewis, Raymond J. (janeiro de 1996). «Chromosomes of the brown algae». Phycologia. 35 (1): 19–40. Bibcode:1996Phyco..35...19L. doi:10.2216/i0031-8884-35-1-19.1
- ↑ «VetFolio». www.vetfolio.com. Consultado em 18 de maio de 2021
- ↑ Datz, Craig (2011). «Parasitic and Protozoal Diseases». Small Animal Pediatrics. [S.l.: s.n.] pp. 154–160. ISBN 978-1-4160-4889-3. doi:10.1016/B978-1-4160-4889-3.00019-X
- ↑ Schmidt and Roberts. 1985. Foundations of Parasitology 3ª Ed. Times Mirror/Mosby College Publishing
- ↑ Heywood, P.; Magee, P.T. (1976). «Meiosis in protists. Some structural and physiological aspects of meiosis in algae, fungi, and protozoa». Bacteriological Reviews. 40 (1): 190–240. PMC 413949. PMID 773364. doi:10.1128/mmbr.40.1.190-240.1976
- ↑ Shehre-Banoo Malik; Arthur W. Pightling; Lauren M. Stefaniak; Andrew M. Schurko; John M. Logsdon Jr (2008). «An Expanded Inventory of Conserved Meiotic Genes Provides Evidence for Sex in Trichomonas vaginalis». PLOS ONE. 3 (8 (número de artigo: e2879)). Bibcode:2008PLoSO...3.2879M. PMC 2488364. PMID 18663385. doi:10.1371/journal.pone.0002879
- ↑ Speijer, Dave; Lukeš, Julius; Eliáš, Marek (21 de julho de 2015). «Sex is a ubiquitous, ancient, and inherent attribute of eukaryotic life». Proceedings of the National Academy of Sciences of the United States of America. 112 (29): 8827–8834. Bibcode:2015PNAS..112.8827S. PMC 4517231. PMID 26195746. doi:10.1073/pnas.1501725112
- ↑ Schön, Isa; Martens, Koen; Dijk, Peter van (2009). Lost Sex: The Evolutionary Biology of Parthenogenesis. [S.l.]: Springer Science & Business Media. ISBN 978-90-481-2770-2
- 1 2 Medvedev, Zhores A. (1981). «On the immortality of the germ line: Genetic and biochemical mechanisms. A review». Mechanisms of Ageing and Development. 17 (4): 331–359. PMID 6173551. doi:10.1016/0047-6374(81)90052-X
- 1 2 Bernstein, C.; Bernstein, H.; Payne, C. (1999). «Cell Immortality: Maintenance of Cell Division Potential». Cell Immortalization. Col: Progress in Molecular and Subcellular Biology. 24. [S.l.: s.n.] pp. 23–50. ISBN 978-3-642-08491-1. PMID 10547857. doi:10.1007/978-3-662-06227-2_2
- ↑ Avise, John C. (outubro de 1993). «Perspective: The evolutionary biology of aging, sexual reproduction, and DNA repair». Evolution. 47 (5): 1293–1301. PMID 28564887. doi:10.1111/j.1558-5646.1993.tb02155.x
Ver também
- Tempo de geração
- Ciclo parassexual
- Partenogénese
- Metamorfose
- Biologia reprodutiva
- Recombinação mitótica
Bibliografia
- van den Hoek, C.; Mann; Jahns, H. M. (1995). Algae: An Introduction to Phycology. [S.l.]: Cambridge University Press. ISBN 978-0-521-31687-3
- Bonner, John Tyler (1995). Life Cycles: Reflections of an Evolutionary Biologist. [S.l.]: Princeton University Press. ISBN 978-0-691-00151-7
- Valero, Myriam; Richerd, Sophie; Perrot, Véronique; Destombe, Christophe (janeiro de 1992). «Evolution of alternation of haploid and diploid phases in life cycles». Trends in Ecology & Evolution. 7 (1): 25–29. Bibcode:1992TEcoE...7...25V. PMID 21235940. doi:10.1016/0169-5347(92)90195-H
- Mable, Barbara K.; Otto, Sarah P. (1998). «The evolution of life cycles with haploid and diploid phases». BioEssays. 20 (6): 453–462. doi:10.1002/(sici)1521-1878(199806)20:6<453::aid-bies3>3.0.co;2-n