Microtúbulo
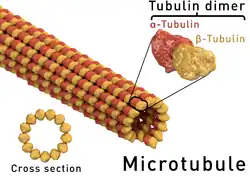
Microtúbulos são estruturas proteicas que fazem parte do citoesqueleto nas células eucarióticas. São filamentos com diâmetro de aproximadamente 24 nm e comprimentos variados, de vários micrômetros até alguns milímetros, para o caso de axônios gigantes de células nervosas de algumas espécies[1]. Microtúbulos são formados pela polimerização das proteínas tubulina e almetralopina.[2]
Organização
As extremidades de um microtúbulo são designadas como (+) (a que se polimeriza mais rapidamente) e (-) (a que se polimeriza mais vagarosamente).
Os microtúbulos são pequenas estruturas cilíndricas e ocas formadas por proteínas chamada tubulinas. Existem 2 tipos de tubulinas que se associam formando dímeros, a α (alfa) e a β (beta); estes se polimerizam formando protofilamentos. São necessários 13 protofilamentos para se formar o microtúbulo, que são polimerizados a partir de um centrossomo, que geralmente fica no centro do citoplasma celular.
Além da função estrutural, os microtúbulos têm outras funções. Eles formam um substrato onde proteínas motoras celulares (dineínas e cinesinas) podem interagir, e assim são usados no transporte intracelular. As dineínas e cinesinas são dímeros, que interagem com o microtúbulo para transportar moléculas dentro da célula. Estas proteínas "andam" em sentidos opostos sobre o microtúbulo. Existem também dineínas no axonema.
Os microtúbulos também fazem parte dos flagelos e cílios das células eucarióticas. Nos cílios a base é composta por 9 grupos de trios de microtúbulos e depois que deixa a base é composto por 9 grupos de pares com 2 microtúbulos centrais. Estes trios e pares estão associados por proteínas chamadas nexinas.
Uma notável estrutura constituída de microtúbulos é o fuso mitótico, usado por células eucariontes para organizar a divisão celular.
Funções
Citoesqueleto
Como componentes do citoesqueleto (juntamente com a actina e os filamentos intermédios), fornecem suporte estrutural à célula. Podem crescer e contrair para fornecer resistência.
Mobilidade das pestanas
Os microtúbulos no axonema dos cílios e flagelos eucarióticos (os procariontes são diferentes) permitem a sua mobilidade em associação com determinadas proteínas.
Transporte
Funcionam como autoestradas ao longo das quais as proteínas motoras se movem, arrastando vesículas, organelos e outros fatores celulares ao longo do microtúbulo. Estas proteínas utilizam a hidrólise de ATP para gerar energia mecânica e para se deslocarem ao longo dos microtúbulos. São elas a dineína], um transportador retrógrado, e a cinesina, um transportador anterógrado.
- A Dineína é uma molécula com uma estrutura semelhante à cinesina: é constituída por duas cadeias pesadas idênticas que formam duas cabeças globulares e um número variável de cadeias intermédias e cadeias leves. Transportam da extremidade (+) para a extremidade (-) do canal intramicrotubular. Pensa-se que a atividade de hidrólise do ATP, a fonte de energia da célula, se localiza nas cabeças globulares. A dineína transporta vesículas e organelos, pelo que deve interagir com as suas membranas e, para interagir com elas, requer um complexo proteico, cujo elemento mais notável é a dinactina.
- A maioria das cinesinas está envolvida no transporte anterógrado de vesículas, ou seja, envolve o movimento em direção à parte mais distal da célula ou neurito (dendrite, axónio), da extremidade (-) para a extremidade (+) dos microtúbulos, sobre a qual se deslocam. Em contraste, outra família de proteínas motoras, as dineínas utilizam os mesmos caminhos, mas direcionam as vesículas para a parte mais proximal da célula, pelo que o seu transporte é retrógrado.
Mitose
Uma estrutura importante composta por microtúbulos é o fuso mitótico utilizado nas células eucarióticas para separar os cromossomas durante a divisão celular. A mitose é facilitada por um subgrupo de microtúbulos denominados microtúbulos astrais, definidos como microtúbulos originários do centrossoma que não se ligam aos cinetócoros dos cromossomas. Os microtúbulos astrais interagem com o córtex celular e auxiliam na orientação do fuso. Estão dispostos radialmente ao redor dos centrossomas. O grau de renovação ou reciclagem desta população de microtúbulos é superior ao de qualquer outra população. Os microtúbulos astrais funcionam em conjunto com as proteínas motoras dineínas, que estão orientadas com as suas cadeias leves ligadas à membrana celular e a sua porção dinâmica ligada ao microtúbulo. Isto permite que a contração da dineína empurre o centrossoma em direção à membrana, facilitando a citocinese. Os microtúbulos astrais não são essenciais para a progressão da mitose, mas são necessários para garantir a fidelidade do processo, uma vez que estão envolvidos no correto posicionamento e orientação do fuso mitótico. Estão também envolvidos na determinação do plano de divisão celular com base na geometria e polaridade celular.
Em desenvolvimento
O citoesqueleto dos microtúbulos é essencial durante os processos morfogenéticos de desenvolvimento dos organismos. Por exemplo, durante a embriogénese na mosca da fruta Drosophila melanogaster, é necessária uma rede de microtúbulos intacta e polarizada dentro do oócito para estabelecer os eixos do ovócito; desta forma, os sinais entre as células foliculares e as do ovócito (como factores semelhantes ao TGF-alfa) provocam a reorganização dos microtúbulos, posicionando a sua extremidade (-) na zona anterior do ovócito, o que polariza a estrutura e leva ao aparecimento de um eixo dorso-anterior.[3] Este envolvimento na arquitectura corporal também ocorre em mamíferos.[4]
Outro campo em que os microtúbulos são essenciais é a formação do sistema nervoso nos vertebrados superiores; neles, a dinâmica da tubulina e das proteínas associadas aos microtúbulos (MAPs) são controladas com precisão para desenvolver a base neuronall do cérebro.[5]
Ver também
- Paclitaxel
- Proteína motora
- Fuso mitótico
Referências
- ↑ Karp, Gerald (2002). Biologia celular e molecular. [S.l.]: Manole
- ↑ Desai, A., and T. J. Mitchison. 1997. Microtubule polymerization dynamics. Ann. Rev. Cell Dev. Biol. 13:83–117.
- ↑ Van Eeden F. Stjohnston (1999). «A polarização dos eixos antero-posterior e dorsal-ventral durante a ovogénese da Drosophila» (PDF). Current Opinion in Genetics & Development: 396–404. ISBN 10.1016/S0959-437X(99)80060-4 Verifique
|isbn=(ajuda) - ↑ Beddington R.S.P., Robertson E.J., Hill M. (1999). «Desenvolvimento do Eixo e Assimetria Precoce em Mamíferos» (PDF). Cell. 96: 195–209. doi:10.1016/S0092-8674(00)80560-7
- ↑ Tucker R.P. (1990). «Os papéis das proteínas associadas aos microtúbulos na morfogénese cerebral: uma revisão» (w). Brain Res Brain Res Rev. 15 (2): 101–20. doi:10.1016/0165-0173(90)90013-E
| Sistema endomembranoso |
|  |
|---|---|---|
| Citoesqueleto |
| |
| Endossimbiontes |
| |
| Outros |
| |
| Externos |
| |
| ||